
Abstract
Introduction:
Grasshoppers and locusts are widely distributed worldwide, causing significant losses in agriculture. The origin and functions of their haemocytes are not entirely understood.
Objectives:
Insect haemocytes arbitrate cellular defence and participate in humoral defences. Due to their importance, the haemocytes of 35 species of grasshoppers and locusts from China were morphologically examined in this study. We aim to highlight a simple method for the morphological examination of insect haemocytes.
Methods:
The haemocytes were observed, counted and compared under a light microscope after Wright-Giemsa staining.
Results:
High complexity in form and shape were observed in the haemocytes. These include prohaemocytes, plasmatocytes, granulocytes, vermicytes, podocytes and megakaryocytes. No clear relationship was seen between the haemocyte type and their phylogenetic relationship among the three families examined. The high abundance of plasmatocytes and granulocytes suggests their importance in the immunity of grasshoppers and locusts. The minor haemocyte populations including prohaemocytes, vermicytes and podocytes may not be always present in individuals.
Conclusion:
All examined species shared similarities in their haemocyte types. Wright-Giemsa staining is a simple and efficient method for evaluating haemocytes.
Introduction
Insects thrive on Earth in various habitats and exhibit species richness, partially because of their efficient and versatile immune systems.1–3 Unlike mammalian species, insects lack adaptive immunity and rely only on innate immunity to protect themselves from pathogens and invaders.4,5 The innate immunity of insects consists of two tightly interconnected components: humoral and cellular defences. 6 Insect haemocytes arbitrate cellular defence and participate in humoral defences.2,7–9 Thus, the capacity to isolate and distinguish haemocyte types is fundamental to the investigation of insect immunity. Insect haemocytes, which are motile cells with a lifespan of days to weeks, are responsible for regulating the extracellular environment and immune responses such as the synthesis, uptake and transport of specific materials, phagocytosis, nodulation, encapsulation, wound healing and melanisation.1,4,8,10
The insect immune response shares strong structural and functional similarities with the mammalian innate immune system. 5 Although insects are excellent models for evaluating mammalian innate immunity, only a few studies have examined the cellular defence of insects including the fruit fly (Drosophila melanogaster), greater wax (Galleria mellonella) and tobacco hornworm (Manduca sexta).2,4 Orthopterans are distributed worldwide and many have a destructive impact on cereals and pastures.11,12 They can proficiently eradiate substantial amounts of aboveground vegetation during outbreaks. 13 Because of this feeding behaviour, several locust and grasshopper species such as the migratory locust (Locusta migratoria), desert locust (Schistocerca gregaria), Chinese rice grasshopper (Oxya chinensis), and Italian locust (Calliptamus italicus) are considered as the most notorious crop pests.4,14–17 Thus, studies of Orthopteran have focused on developing effective control strategies. 12
Despite advances in the understanding of Orthopteran physiology, behaviour and control, haemocyte morphology has not been widely examined. The criteria and methods used to classify haemocytes are not unique, and many haemocyte types remain controversial.7,18,19 The origin and functions of haemocytes are not entirely understood. 1 In this study, we morphologically evaluated haemocytes from 35 species of grasshoppers and locusts in China to improve the classification of grasshopper haemocytes and determine the validity of reported haemocyte types.
Materials and Methods
Insects
Thirty-five species in three families and 27 genera of grasshopper were collected from different locations in China (Table 1). Samples were collected alive and kept in a well-ventilated room in the laboratory. The sampled species were fed with rice for at least 1 week before blood smear preparation.
Differential haemocyte counts in the sampled grasshopper and locust species from China.

PR: prohaemocyte; PL: plasmatocyte; GR: granulocyte; VE: vermicyte; PO: podocyte; ME: megakaryocyte, absent ‘–’.
By comparing all examined species, Kruskal–Wallis test showed statistical difference for PR: H (30, N = 152) = 51.86, p < 0.01; PL: H (33, N = 336) = 76.26, p < 0.01; GR: H (33, N = 336) = 84.97, p < 0.01.
No statistical difference was found for VE: H (8, N = 57) = 5.03, p = 0.75; PO: H (7, N = 21) = 1.71, p = 0.97; ME: H (20, N = 58) = 17.37, p = 0.63.
Blood smear preparation
Ten adults (five male and five females) were selected randomly from each species, and a total of 50 slides (i.e. five slides for each grasshopper) were prepared from the haemolymph. The prothoracic leg bases of each selected grasshopper were disinfected with 75% ethanol. The skin of each base was pierced to extract the haemolymphs with a pipette (MicroPette Plus, Scilogex, Rocky Hill, CT, USA). The blood smears were stained with Wright-Giemsa stain and observed under a light microscope (Olympus BX-51, Tokyo, Japan). The Wright-Giemsa staining procedure was amended from Ye et al. 20 and Dunning and Safo. 21
Wright-Giemsa staining reagents:
Solution A: 1 g Wright powdered dye, 1 g Giemsa powdered dye, and 10 mL glycerol were mixed well and dissolved in 500 mL methanol; Buffer (pH 7.2): 1 g potassium phosphate monobasic and 1 g dibasic sodium phosphate powder was dissolved in 100 mL distilled water; Solution B: Wright-Giemsa Staining (Baso Diagnostics, Inc., Guangdong, China) solution. 4–5 drops of solution A were added to the smears for 1 min; 2 drops of solution B were added to the smears for 2 min; 10–20 drops of buffer were added to the smears evenly; Smears were stained for 10–20 min, washed with distilled water, and air-dried.
Combine as follows:
Haemocyte identification
The study by Gupta 18 was used as a reference for haemocyte identification in this study. Descriptions from later studies were considered.19,22–24 Previously described haemocytes in different grasshoppers and locusts included: prohaemocytes, small spherical cells with minimum cytoplasm containing no inclusions; plasmatocytes, spherical, oval, or spindle-shaped cells with abundant cytoplasm and likely to contain pinocytotic inclusions; granulocytes (or granular cells), polymorphic shaped cells filled with small granules; vermicytes, agranular or granular worm-shaped cells with extremely elongated cytoplasm and nucleus, and megakaryocytes, large cells measured more than 30 µm in diameter with nearly no cytoplasm and no inclusions.
Haemocytes counting and statistical methods
For morphological comparison, haemocytes were systematically counted, and the first 100 cells of each grasshopper or locust observed were identified and recorded. All additional cells were examined for photographic records and to assess any missed types during counting. Data obtained during the study were analysed using Statistica 12 software (StatSoft, Tulsa, OK, USA). 25 Statistical differences were examined by Kruskal–Wallis test since samples were not normally distributed. The means and standard errors were used to summarise the data sets. When the p-value was less than 0.01, the difference was considered significant.
Results
The haemocytes observed in this study varied distinctively in their size, shape and cellular contents. Six morphological categories of haemocytes were found, which included prohaemocytes, plasmatocytes, granulocytes, vermicytes, podocytes and megakaryocytes. The number of different types of haemocyte varied among species (Table 1). Four species, Calliptamus barbarous, Diabolocatantops pinguis, Phlaeoba infumata and Pseudotmethis rubimarginis contained all six types. Only plasmatocytes and granulocytes were found in all examined species, accounting for more than 90% of the total haemocytes observed (Figure 1). Prohaemocytes, plasmatocytes, granulocytes and megakaryocytes were observed in all three examined families. Vermicytes and podocytes were not found in Pyrgomorphidae. Since only two species were studied in this family, Atractomorpha sinensis and Aularches miliaris, it was unreasonable to conclude Pyrgomorphidae was free of vermicytes and podocytes. More examples include that vermicyte and podocytes were found in Bryodema nigroptera but not B. gebleri.
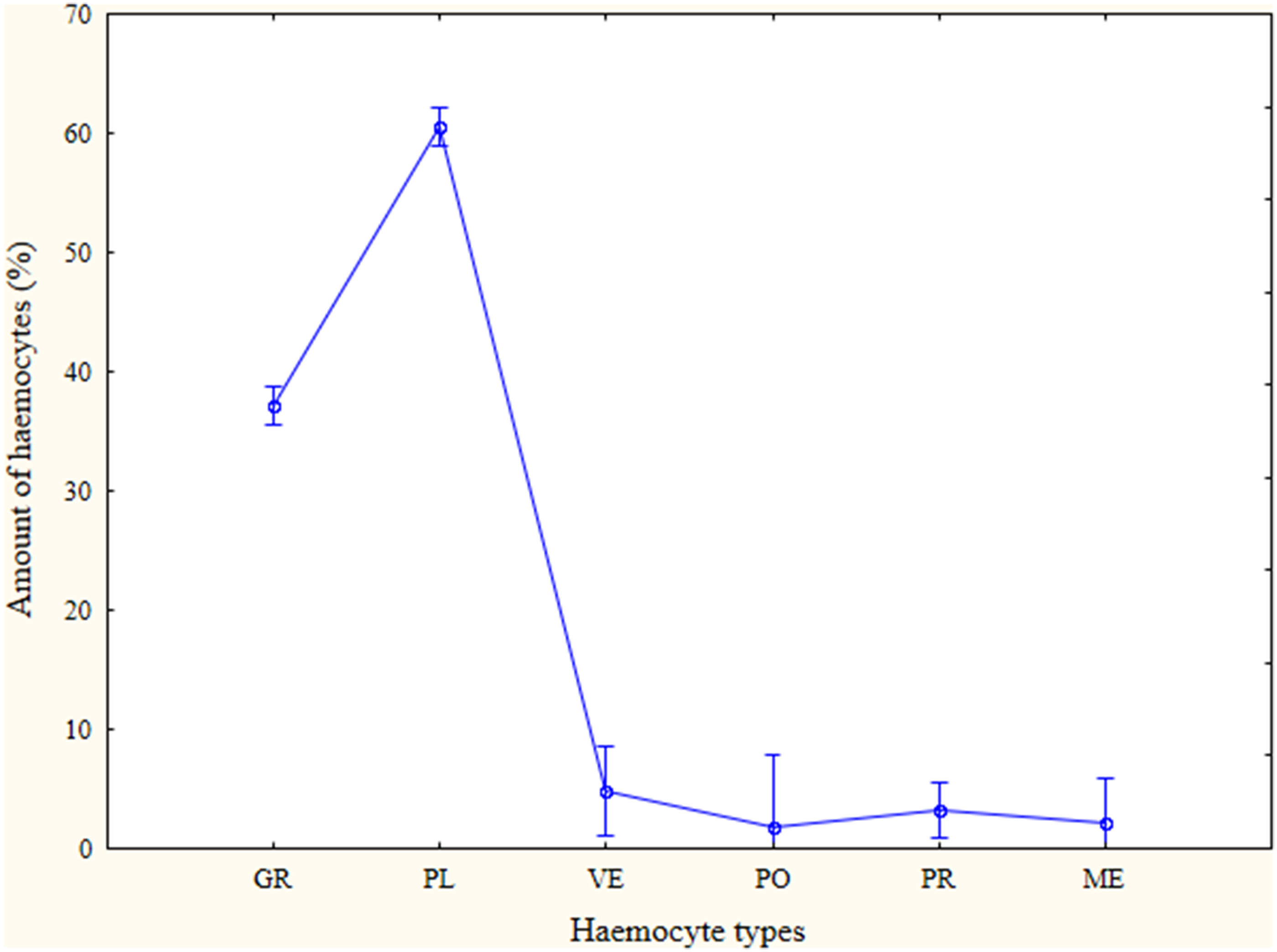
The prevalence of different haemocytes summarised from all examined species.
Plasmatocytes were the most abundant cell type found in this study while granulocytes being the second most abundant type (Table 1 and Figure 1). The other types were less encountered. The prevalence of prohaemocytes, plasmatocytes and granulocytes differed between the examined species (Table 1). Vermicytes, podocytes and megakaryocytes showed no difference between species.
Prohaemocytes were found in all studied species except Gastrimargus marmoratus, Trilophidia annulata and Phlaeoba antennata. They were the smallest haemocytes found, which displayed a round or oval shape of approximately 6–12 µm in diameter (Figure 2). The Wright-Giemsa-stained purple or violet nucleus occupied the whole cell and contained nearly no visible cytoplasm.
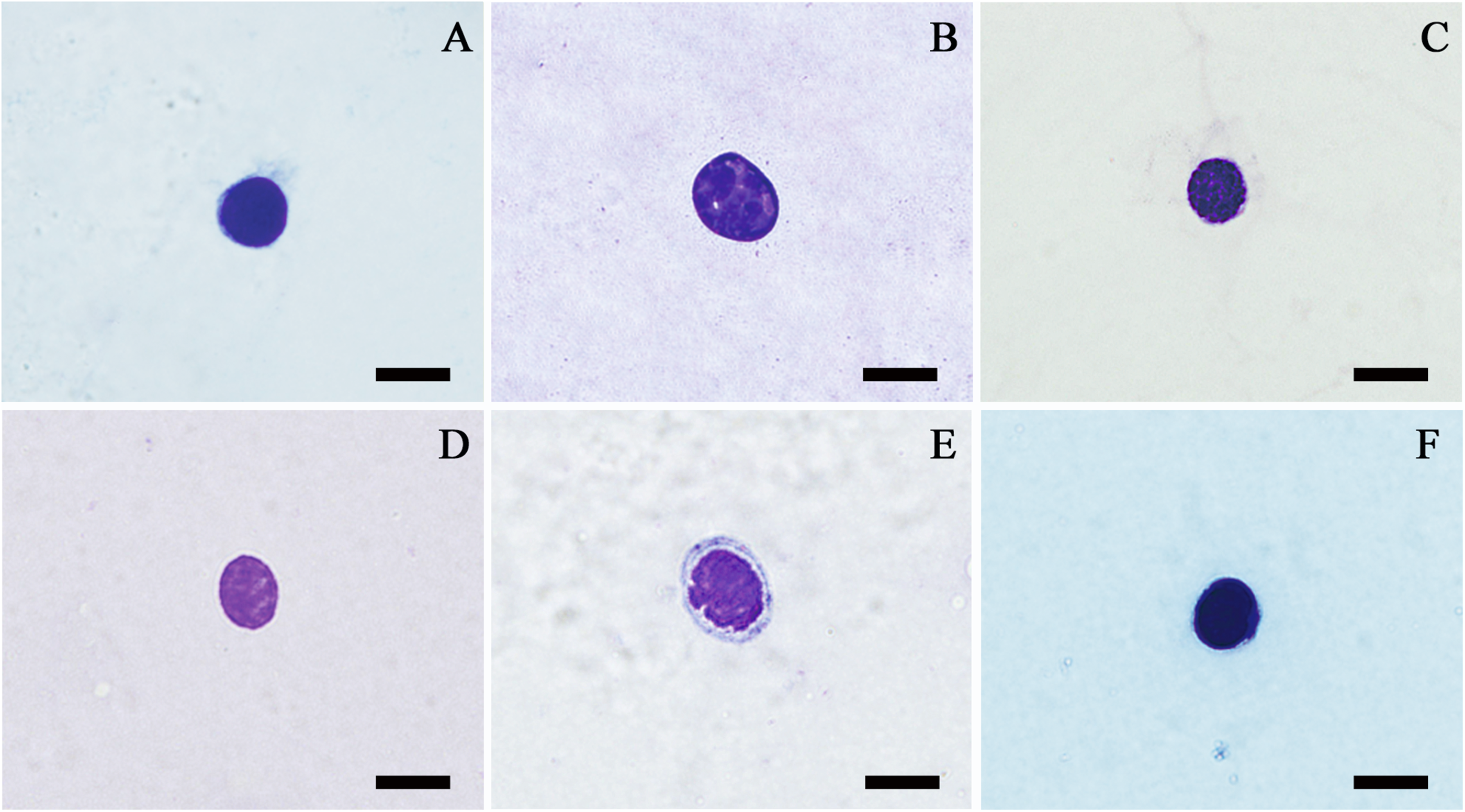
Prohaemocytes of Haplotropis brunncriana (A), Locusta migratoria (B), Dasyhippus peipingensis (C), Aiolopus tumulus (D and E) and Pseudotmethis rubimarginis (F) evaluated by light microscopy; bars denote 5 µm.
Plasmatocytes were found in all species examined. They varied enormously in size and shape, mostly because of the divergence of their pseudopodia and other cytoplasmic projections. Round, oval, spindle and irregularly shaped plasmatocytes had a size of 5–34 µm in diameter (Figure 3). The size and shape of plasmatocytes can differ markedly even within the same individual. The cytoplasm of plasmatocytes was clear, pinkish red or light blue. The nucleus of plasmatocytes was stained pink, purple or dark blue with a size of 6–21 µm in diameter. Occasionally, the nucleus of plasmatocytes was eccentrically located.

Plasmatocytes of Calliptamus barbarus (A), Choroedocus violaceipes (B), Catantops pinguis (C), Calliptamus abbreviatus (D), Ceracris fasciata fasciata (E), Bryodemella tuberculatum dilutum (F), Locusta migratoria (G), Acrida cinerea (H) and Phlaeoba infumata (I) evaluated by light microscopy; bars denote 5 µm.
Granulocytes were found in all species observed in this study. The round, oval, spindle and irregularly shaped granulocytes were 6–44 µm in diameter under the light microscope (Figure 4). The centrally located nucleus was dyed with Wright-Giemsa stain as dark red or bluish purple with a diameter of 5–24 µm. Most of the granulocyte cytoplasm was dyed as light blue, while other parts were transparent or pink. Granular inclusions or granules were used to separate granulocytes from plasmatocytes. Most granules found in the cytoplasm were stained as pink, purple or dark blue. The granules in the cytoplasm had either a uniform or patchy distribution, and the density of the granules ranged from few to packed.
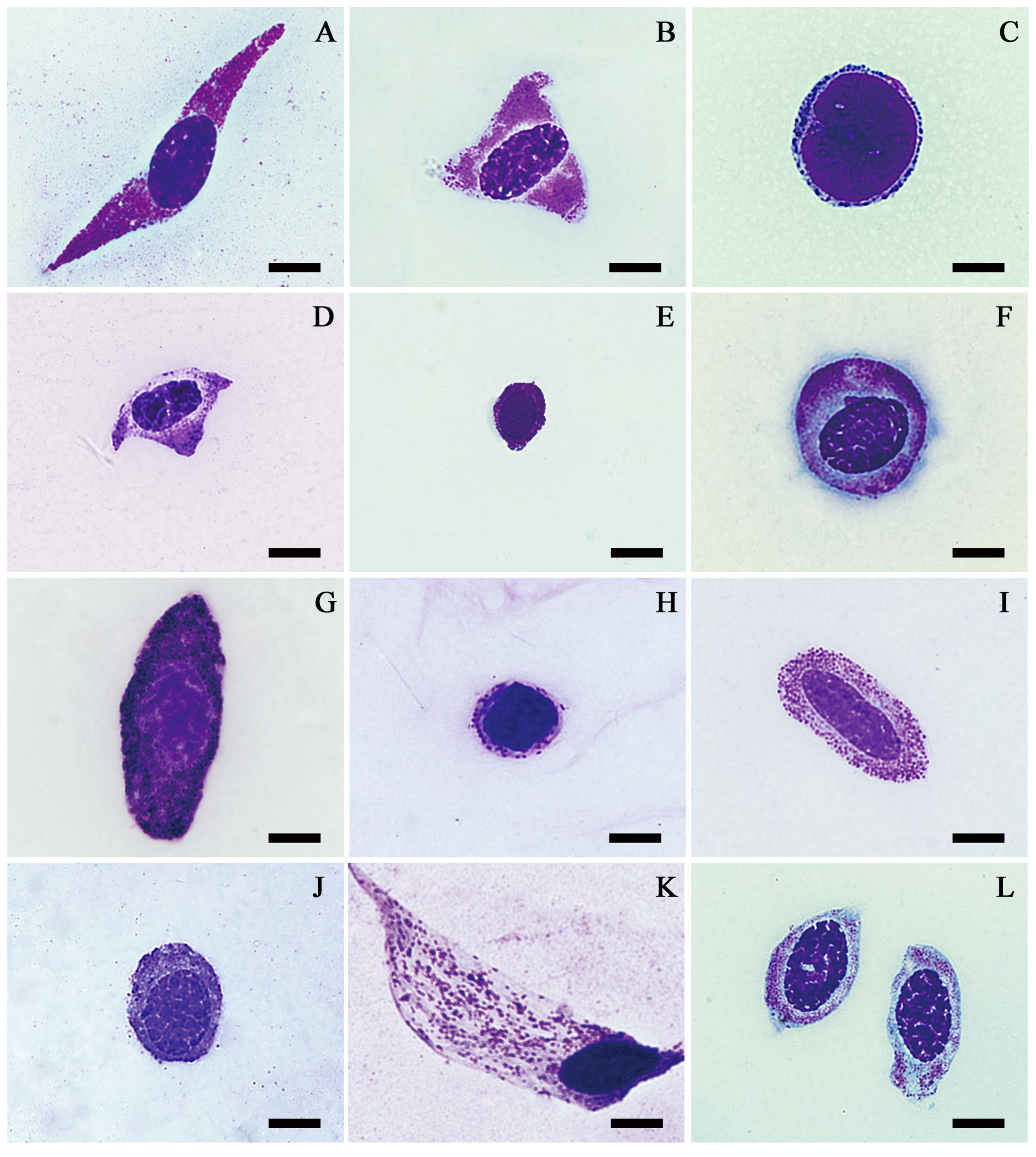
Granulocytes of Phlaeoba infumata (A), Catantops pinguis (B), Mecostethus grossus (C), Locusta migratoria (D), Gesonula punctifrons (E), Pseudotmethis rubimarginis (F), Bryodemella tuberculatum dilutum (G), Bryodema gebleri (H), Choroedocus violaceipes (I), Calliptamus italicus (J), Bryodema nigroptera (K) and Catantops pinguis (L) evaluated by light microscopy; bars denote 5 µm.
Vermicytes were recognised in several examined species by their distinct worm-shaped structure which cannot be regarded as plasmatocytes (Figure 5). The cell body and nucleus of vermicytes were stretched. Vermicytes were 30–115 µm long with clear to light blue or pink cytoplasm. The purple or dark blue nucleus was mostly in the centre of the cell with a diameter of 11–24 µm. Vermicytes were either absent or sparse and filled with purple or dark blue granules.

Vermicytes of Pseudotmethis rubimarginis (A and C), Calliptamus barbarus (B, D and F) and Catantops pinguis (E) evaluated by light microscopy; bars denote 5 µm.
A group of haemocytes with multiple elongated cytoplasmic extensions and containing granules did not match the previously described haemocyte characteristics of grasshoppers and locusts. These haemocytes were morphologically consistent to rarely found podocytes observed in other insects. Podocytes, which were not previously found in grasshoppers or locusts, were found in seven examined species including B. tuberculatum dilutum, C. barbarous, C. violaceipes, D. pinguis, P. infumata, P. rubimarginis and S. splendens. They had multiple cytoplasmic extensions (3–4) of 12–80 µm long (Figure 6). All species containing podocytes had vermicytes, but not vice versa. Podocytes can be agranular or granular. The cytoplasm was dyed transparent to blue. The round or oval nucleus was centrally located with a diameter of 9–18 µm. The nucleus and granules were stained as purple or dark blue. The distinctive feature of podocytes was their multiple cytoplasm extensions radiating from the centrally located nucleus. Three to four extensions with a length of more than 40 µm were observed.
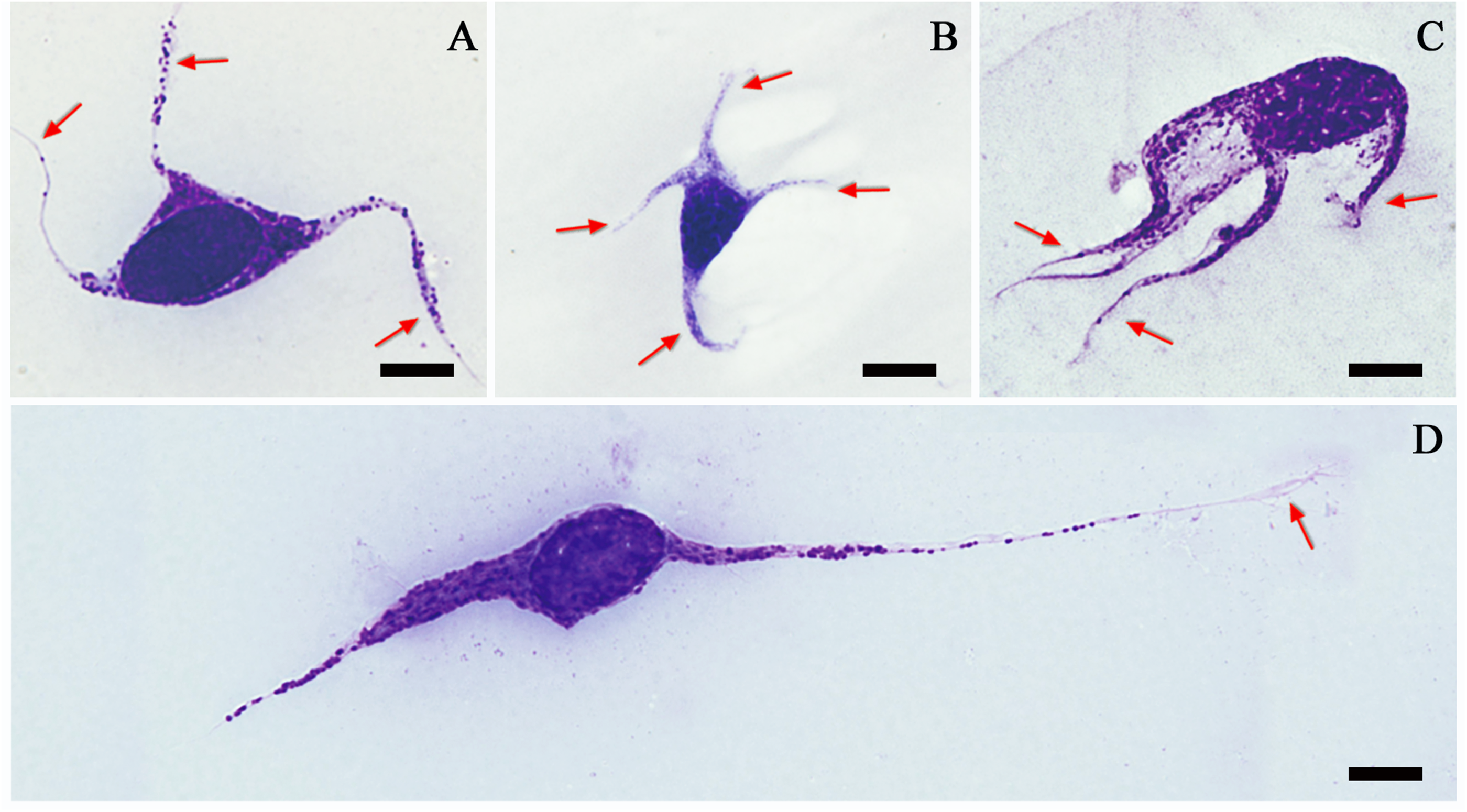
Podocytes of Pseudotmethis rubimarginis (A and D) and Calliptamus barbarous (B and C) evaluated by light microscopy showing multiple cytoplasmic extensions (arrows); (D) this haemocyte was classified as a podocyte because of its round nucleus and extremely elongated cytoplasmic extension (arrow); bars denote 5 µm.
Many enormous haemocytes with a remarkably large nucleus, which we regarded as megakaryocytes, were found in 21 species (Figure 7). They displayed a round or oval shape with a diameter of 22–55 µm. The nucleus of megakaryocytes occupied the entire cell with a diameter of 17–55 µm. The cytoplasm of megakaryocytes was often invisible. The nucleus was purplish red or bluish purple after Wright-Giemsa staining. Small blocks of chromatins were uniformly distributed in the nucleus.

Megakaryocytes of Bryodema gebleri (A), Phlaeoba antennata (B and D), Bryodema mongolicum (C), Ceracris fasciata fasciata (E) and Oxya chinensis (F) evaluated by light microscopy; bars denote 5 µm.
Phagosomes or pinocytic vesicles of up to 2 µm in diameter and attachments of foreign objects of more than 3 µm in diameter were found in plasmatocytes of some species. Phagosomes or pinocytic vesicles of up to 5 µm in diameter were observed in the granulocytes (Figure 8).

Phagocytes of Phlaeoba infumata (A), Haplotropis brunncriana (B), Trilophidia annulata (C), Stethophyma grossum (D–F), Diabolocatantops pinguis (G), Chondracris rosea (H) and Pseudotmethis rubimarginis (I) attaching or engulfing phagosome-like vacuoles and large particles; arrows point to some of the engulfed particles.
Discussion
Prohaemocytes, plasmatocytes, granulocytes, vermicytes, podocytes and megakaryocytes were found in the examined species. The high abundance of plasmatocytes and granulocytes found was similar to in previously reported grasshoppers and locusts.12,19
The classification of insect haemocytes remains challenging, partially because of the absence of a standardised identification protocol. 19 Marked differences exist in the classification schemes between Drosophila and other insects. 26 The classification criteria were found to be ambiguous for specific haemocyte types. Some studies consider adipohaemocytes as a distinguishable haemocyte, while others report them as granulocytes, spherulocytes, cystocytes or even plasmatocytes.10,18,27–29 The adipohaemocytes reported in A. cinerea were morphologically similar to spherulocytes with a high number of large granules (their lipid droplets) which push the nucleus aside. 30 However, whether these refractive granules are lipids and if adipohaemocytes have specific roles in insect circulation require further analysis.
Prohaemocytes have been found in all reported haemocytes of Orthoptera.4,10,19,31 They are thought to be the stem cells of insect haemocytes that differentiate into other cells types. 26 Several studies have supported this hypothesis.7,27,32 Apart from the simple morphology, prohaemocytes of mosquitos (Anopheles gambiae and Aedes aegypti) were not labelled by any functional markers used. 7 Thus, prohaemocytes are likely to be the progenitor cell. The abundance of prohaemocytes was highest during early instars of L. migratoria and was extremely low in later stages. 27 Prohaemocytes were found to increase in number during recovery following blood loss. 32 The function of the mosquito prohaemocyte differs from that in other insect species as being phagocytic. 33 Because of the low abundance of prohaemocytes in many species, their importance in insect cellular defence has been questioned. Further studies beyond the evaluation of cytological features are needed to confirm that prohaemocytes are progenitors of insect haemocytes. 34
The small stained granules observed in the cytoplasm were used to distinguish granulocytes from plasmatocytes.2,19,34,35 Plasmatocytes and granulocytes are frequently observed in major arthropod groups and recognised as the primary defensive cells.2,18,36 The role of granulocytes, phagocytosis, is supported by their strong acid phosphatase activity.9,10,33,37 Additionally, plasmatocytes and granulocytes were found to participate in other processes such as encapsulation. 2 In Lepidoptera, for example, Gallaria mellonella and Clitumnus extradentalus, capsule formation is biphasic. Granulocytes contact foreign objects that cannot be phagocytosed and disintegrate or degranulate, liberating material that promotes the attachment of plasmatocytes to form multiple layers around the target into a capsule.2,9
The presence of vermicytes and podocytes has rarely been reported in grasshoppers and locusts. Vermicytes and podocytes are occasionally observed in specific stages of insect life, and many studies have classified vermicytes and podocytes as a subclass or variant form of plasmatocytes.32,34,35,38 Distinguishable features exist between vermicytes and podocytes, including the nucleus shape and number of cytoplasmic extensions. The function of vermicytes and podocytes has not been confirmed. The distinct morphology and extended pseudopodia enable vermicytes and podocytes to attach to foreign objects. However, vermicytes and podocytes have not been shown to emerge in encapsulation; thus, their role as plasmatocytes remains to be confirmed. 34 In this study, species that processed podocytes were found to contain vermicytes. Thus, the development of both vermicytes and podocytes likely occurs under similar conditions and stimuli.
Enormously large cells, megakaryocytes, which contain nearly no cytoplasm, were found in serval species and differ from insect haemocytes described in the literature. They are morphologically like an enlarged version of prohaemocytes. These cells have not been found in insects except in our previous studies as contaminated samples in L. migratoria and conceivably haemocytes in B. nigroptera.24,39 In this study, many species contained megakaryocytes with relatively low abundance. Because of the limited cytoplasm and absence of specialised inclusions, the capacity of megakaryocytes to synthesise specific enzymes or peptides is low. Whether megakaryocytes fall off from the surrounding tissues or circulatory haemocytes specific to grasshoppers and locusts remains questionable.
We did not detect adipohaemocytes in Acrida cinerea as previously reported by Yu et al. 30 Additionally, we observed no spherulocytes in L. migratoria as reported by Duressa and Huybrechts. 12 Specific extrinsic factors may cause variations in haemocyte types. Haemocytes are more susceptible than any other cell type to different environmental factors such as temperature, insecticides or plant-derived compounds.8,40
The classification of cystocytes or coagulocytes is also controversial. Some authors prefer using the name ‘cystocyte’ to illustrate its morphology,18,35 while others use ‘coagulocyte’ to indicate its function.12,27 An early study by Costin 27 claimed that cystocytes (their coagulocytes) are histochemically distinct from granulocytes according to periodic acid-Schiff staining, while most studies classify cystocytes as having a similar morphology to granulocytes with fewer small granules and are often lysed in vitro with cellular contents or coagulum scattered around the naked nucleus.18,41,42 It is difficult to distinguish cystocytes from granulocytes based on granule abundance, and the substance around the nucleus is assumed to be clotted haemolymph without functional evidence. In a study of L. migratoria, cystocytes (their coagulocytes) were identified as a sub-type of granulocytes. 19 Studies are needed to determine whether cystocytes should be considered as granulocytes. Cystocytes were reported to contain lysozyme and form endocytosed vesicles against the injected iron saccharate,32,42 which is functionally indistinguishable from granulocytes. The most widely reported function of cystocytes is the coagulation of insect haemolymph.2,18 However, Dushay 43 found that separation of the haemocytes and plasma can block haemolymph coagulation, and the haemocytes and plasma factors cooperatively activated the coagulation process. Additionally, the contents of granulocytes were involved in haemolymph coagulation and wound healing of the greater wax moth G. mellonella,10,43,44 L. migratoria 44 and mosquitoes. 33 Thus, functional evidence is required to confirm the classification and function of cystocytes.
Conclusions
Insects contain haemocytes that show considerable variation in morphology and function. Plasmatocytes and granulocytes are the essential types found in grasshoppers and locusts and participate in phagocytosis. The light microscopic investigation combined with Wright-Giemsa staining is effective for examining the morphology of insect haemocytes. However, genetic and functional markers should be used to enable comprehensive analysis. Orthopterans are essential to the ecosystem and economy. A better understanding of their immune system would have not only economic benefits but also provide models for understanding the human immune system.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the National Natural Science Foundation of China (grant number 30770239) and the Natural Science Foundation of Shanxi Province, China (grant number 2009011048).