
Abstract
DDIT4 is a mitochondrial and tumor-related protein involved in anti-tumor therapy resistance, proliferation, and invasion, etc. Its expression level increases under the stress such as chemotherapy, hypoxia, and DNA damage. A number of clinical studies have confirmed that DDIT4 can change the behavior of tumor cells and the prognosis of patients with cancer. However, the role of DDIT4 in promoting or suppressing cancer is still inconclusive. This article summarized the four characteristics of DDIT4 including a mitochondria-related protein, interactions with various protein molecules, immune and metabolic cell related proteins and participator in the oxygen sensing pathway, which may be related to the progress of cancer.
Introduction
DNA damage-inducible transcript 4 (DDIT4) is a tumor-associated protein as well as a metabolic and immune related protein, which is highly expressed under the stress conditions such as chemotherapy, hypoxia, and DNA damage. It showed significant differences in expression levels in malignant cancer such as lung cancer, 1 bladder urothelial carcinoma, 2 and ovarian cancer, 3 and DDIT4 affected the prognosis of patients with triple-negative breast tumours 4 and lung adenocarcinoma. 5 DDIT4 acts as a “protective umbrella” in the anti-tumor therapy of many cancers while it was involved in DNA damage repairing in some specific environment. The possible reasons for the interesting phenomenon included that. First, DDIT4 was a mitochondria-related protein 6 that participates in the function and survival of tumor cells by affecting the autophagy and apoptosis. Second, it directly participated in the activation of several important pathways through interactions with various protein molecules including mTORC1, P53, HIF, ATF4, and miR-221. Thirdly, the function changed in many tumor related cells including adipocytes, immune cells, and muscle cells via the up-regulation of DDIT4 expression (Figure 1). Lastly, DDIT4 may affect the occurrence and development of cancer by affecting the oxygen sensing regulation of tumor cell and tumor-related cells.

The signaling pathways of DDIT4 and cancer.
Structure, location, and expression regulation of DDIT4
DDIT4, is also known as regulated in development and DNA damage responses (REDD1) or RTP801, which locates on chromosome 10 (10q22.1) and has a length of 2.1 kb. 7 As there are multiple lysine residues at the carboxyl terminus, the protein band is located at 35 KD in Western blot while Its molecular weight is 25 kDa. 8 the previous literature showed that the multiple lysine residues at the carboxyl terminus regulated protein oligomerization and its nucleus-cytoplasm trafficking depending on ubiquitination or acetylation, 9 which may affected the expression and location of DDIT4. The DDIT4 is characterized by a sandwich structure with anti-parallel alpha-helices and a beta-sheet encompassing an uncommon psi-loop motif while the loop between helix α2 and strand β1 as a functional hot spot that is essential for DDIT4 signaling. 10
DDIT4 is mainly expressed in the cytoplasm, 8 and it was confirmed by immunofluorescence staining and confocal microscopy that DDIT4 and mitochondria had obvious co-localization, and it reduced ROS production and regulated metabolism. 11 In DDIT4(-/-) animal model, deregulated DDIT4/TXNIP pro-oxidant complex and ATG4B activity caused accumulation of defective mitochondria, leading to impaired oxidative phosphorylation, and poor energy storage. 12 In the process of inflammation-mediated injury, DDIT4 will affect the autophagy and mitochondrial biogenesis in articular cartilage, and then increased the severity of osteoarthritis. 13
Under normal conditions, DDIT4 is ubiquitously expressed at low levels in most adult tissues, with the lowest expression in brain, skeletal muscle, and intestine. 7 The expression level of DDIT4 is regulated by a variety of factors, such as hypoxia, 7 ionizing radiation, 8 exercise, 14 hexaldehyde, 15 methyl methane sulfonate (MMS), 16 dexamethasone, 17 energy depletion, 18 arsenite, 19 and endoplasmic reticulum stress. 20 In contrast, testosterone may reduce the increase of DDIT4 protein in skeletal muscle and L6 myoblast cells by affecting FOXO1 expression. 21 The study about regulation of DDIT4 showed that DDIT4 may be a cancer related protein, at the same time, the bridge between DDIT4 and cancer may be associated with oxidative stress, metabolism and autophagy. The half-life of DDIT4 protein is very short (about 5 min), and its degradation is affected by intracellular ubiquitin. After releasing from the stress environment, glycogen synthase kinase 3 (GSK3) is exposed to binding sites by inducing DDIT4 (Thr25, Thr2, and Ser19) phosphorylation, while the CUL4A-DDB1-ROC1-beta-TRCP E3 ligase complex mediates DDIT4 ubiquitination and protein degradation. 22
Correlation between DDIT4 and tumor and exploration of its underlying mechanism
In Gene Expression Profiling Interactive Analysis (GEPIA), DDIT4 mRNA expression profile is significantly altered in tumor tissues, with the significant difference in ovarian serous cystadenocarcinoma, kidney renal papillary cell carcinoma, and esophageal carcinoma. 23 The differences of expression level of DDIT4 is characterized by complexity and tissue heterogeneity. A silico analysis in online platforms, in pooled datasets from KM Plotter and meta-analysis of individual datasets from SurvExpress indicated that in acute myeloid leukemia, breast cancer, multiple glioblastoma, colon cancer, skin cancer, and lung cancer, high levels of DDIT4 were significantly associated with poor prognosis. In contrast, high DDIT4 expression was linked to improved prognosis of gastric cancer. Meanwhile, it was not associated with the prognosis of ovarian cancer. 24 Survival analysis about DDIT4 in GEPIA shows that the patients with various tumor including adrenal cortical carcinoma, cervical squamous carcinoma, head and neck squamous cell carcinoma, renal color cell cancer, acute myeloid leukemia, lung adenocarcinoma, mesothelioma, and high expression in patients with pancreatic adenocarcinoma tended to poor prognosis related to high DDIT4, and lower expression of DDIT4 in brain lower grade glioma indicates poor prognosis. 23 (Table 1)
The relationship between of DDIT4 and OS/PFS in patients with cancer.

Cut-off high: 50%; date from GEPIA. 23 Logrank p < 0.05 means that different DDIT4 expression level in tumor patients have different survival rates.
ACC: adrenocortical carcinoma; BLCAB: ladder urothelial carcinoma; BRCAB: reast invasive carcinoma; CESC: cervical squamous cell carcinoma and endocervical adenocarcinoma; CHOL: cholangio carcinoma; COAD: colon adenocarcinoma; DLBC: lymphoid neoplasm diffuse large B-cell lymphoma; ESCA: esophageal carcinoma; GBM: glioblastoma multiforme; HNSC: head and neck squamous cell carcinoma; KICH: kidney chromophobe; KIRC: kidney renal clear cell carcinoma; KIRP: kidney renal papillary cell carcinoma; LAML: acute myeloid leukemia; LGG: brain lower grade glioma; LIHC: liver hepatocellular carcinoma; LUAD: lung adenocarcinoma; LUSC: lung squamous cell carcinoma; MESO: mesothelioma; OV: ovarian serous cystadenocarcinoma; PAAD: pancreatic adenocarcinoma; PCPG: pheochromocytoma and paraganglioma; PRAD: prostate adenocarcinoma; READ: rectum adenocarcinoma; SARC: sarcoma; SKCM: skin cutaneous melanoma; STAD: stomach adenocarcinoma; TGCT: testicular germ cell tumors; THCA: thyroid carcinoma; THYM: thymoma; UCEC: uterine corpus endometrial carcinoma; UCS: uterine carcinosarcoma; UVM: uveal melanoma.
DDIT4 is involved in the survival and drug resistance of many types of tumor cells through a variety of pathways mediated autophagy pathways. 25 DDIT4 promoted the survival of glioblastoma cell and anti-tumor therapy resistance by inhibiting mTORC1, while DDIT4 interfered with tumor-cell death induced by temozolomide, radiotherapy, or hypoxia. 25 The sensitivity of bladder urothelial carcinoma tumor cells to paclitaxel markedly enhanced after silencing DDIT4 expression, and the accurate mechanism may be associated with the DDIT4-eukaryotic elongation factor-2 kinase (eEF2k)-autophagy axis. 2 Overexpressed DDIT4 cell lines led to high expression a levels of HSP27, HSP70, and AKT proteins, which affected lung cancer cell survival and resistance to ionizing radiation. 1 DDIT4 was a critical effector of autophagy via regulation of autolysosome formation and promoted the resistance to proteasome inhibitors (bortezomib) in prostate cancer 26 and myeloma. 27 Ras-mediated transformation of human ovarian epithelial cells relied on DDIT4, which facilitated expression of autophagy-related proteins and inhibited tumor cell apoptosis. 3
DDIT4 is not only involved in tumor survival, anti-tumor therapy resistance and anti-apoptosis, but also participated in important tumor behaviors such as proliferation and invasion. The expression levels of DDIT4 in non-small cell lung cancer (NSCLC) tissue and serum was significantly higher than control group, and knockdown of DDIT4 significantly inhibited the proliferation of A549 and SW900 cells. 28 The over-expression of DDIT4 in ovarian cancer was significantly related to serous cancer, advanced stage, ascites, partial or no response to chemotherapy, overall survival, and tumor-free survival. Meanwhile, the results of transwell migration and invasion assays of various human ovarian epithelial cancer cell lines suggested that DDIT4 was associated with cell invasion and migration. 29 DDIT4 was involved in invasive growth of prostate cancer cells in vitro and in vivo through a loss of function studies. DDIT4 was a common downstream target protein of PI3K-kinase and HIF-1, and played an important role in invasive tumor growth. 30
DDIT4 have a context dependent contrasting role in cancer as oncogene or tumor suppressor. The expression levels of DDIT4 protein in highly invasive H1299 cells was higher than that in less invasive H460 cells. The intervention of DDIT4 protein expression could affect the invasive ability of NSCLC cell lines. 31 Melatonin enhances arsenic trioxide (ATO)-induced apoptosis in human breast cancer cells by continuously up-regulating of DDIT4, and its potential molecular mechanism may be related to mTORC1 and p38/JNK. 32 ZY0511, a Lysine-specific demethylase1 (LSD1) inhibitor, could significantly up-regulate the expression of mTORC1 inhibitor DDIT4 in HeLa and HCT116 cells. 33 DDIT4 is involved in many aspects of tumor behavior such as anti-tumor therapy resistance, proliferation, invasive activity and apoptosis, and its heterogeneity may be related to tumor microenvironment and tumor gene mutation levels (Figure 2).
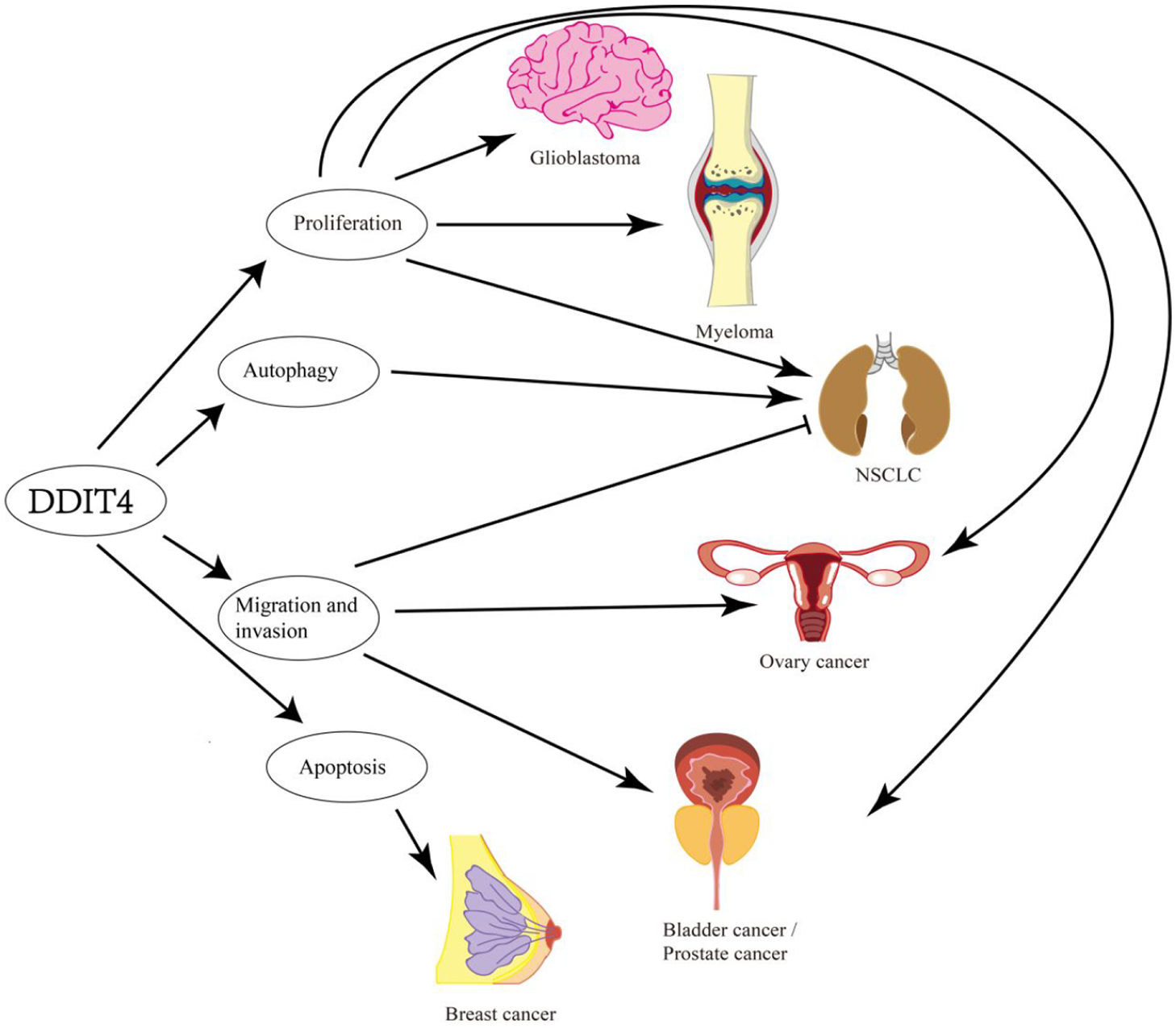
The relationship between DDIT4 and cancer.
DDIT4 is a tumor-related protein and affects many cancers including glioblastoma, bladder urothelial carcinoma tumor cells, prostate cancer, myeloma, ovarian epithelial cells, non-small cell lung cancer, breast cancer. The main influence of tumor behavior is autophagy, proliferation, apoptosis, migration, and invasion.
DDIT4 directly acts on tumor cells through a variety of tumor-related proteins
The phosphorylation level of mammalian target of rapamycin complex 1 (mTORC1) is regulated by DDIT4, 34 located at the core of multiple metabolic pathways, and it is a target for tumor treatment, 35 especially in the cancer exhibiting activated AKT/mTORC cascade. 36 Functional studies proved that DDIT4 inhibited mTORC1 by promoting the formation of TSC1/2 complex by binding to 14-3-3. 37 At the same time, through the structural analysis of DDIT4, the literature denied the direct binding of DDIT4 and 14-3-3, and believed that DDIT4 may affect the phosphorylation of TSC2 through protein phosphatase 2A (PP2A), and then inhibited the phosphorylation of mTORC1. 38 The DDIT4/mTORC1 signaling pathway appeared to be different in a variety of tumor cells. DDIT4 expression which mediated mTOR inhibition and growth inhibition in breast cancer cells in the process of baicalein treatment. 39 A short hairpin RNA-mediated gene suppression test showed that DDIT4/mTORC1 mediated therapy resistance in glioblastoma.
p53 is one of the most important tumor suppressor proteins. The high-frequency mutation of p53 in tumor cells and its extensive pathway signaling have caused great concern. DDIT4 and p53-mediated DNA damage response and p63-mediated epithelial cell development were first proposed in 2002. RNA in situ hybridization showed that p63-deficient embryos showed virtually no expression of DDIT4, and after p63 expression, DDIT4 restored expression in mouse embryonic fibroblasts. 8 Activation of p53/DDIT4 could affect some signaling pathways such as cell proliferation, metabolism, and cell differentiation. 40 DNA damage-induced DDIT4/TSC2/mTORC1 was a key signaling pathway downstream of p53.40,41 Both knockdowns of DDIT4 and p53 had promoted the development of pro-oncogene Myc-driven lymphoma to a similar extent. 42 DDIT4 may mediate the tumor suppressive effect of p53 by affecting DNA damage repair and cell cycle control. 42 As an effector protein of p53, DDIT4 could also inhibits p53 expression in a feedback way. In DDIT4-/- cells, the level of p53 expression and apoptosis induced by DNA damage were significantly higher than that of wild cell lines, and p53 was deregulated, because of the increased protein synthesis of p53. 43
Autophagy induced by DDIT4 is an important cause of anti-tumor therapy resistance in cancer. The DDIT4-mediated proliferation, apoptosis, and autophagy were found to be negatively regulated by miR-22, which enhanced sensitivity to paclitaxel of bladder urothelial carcinoma by inhibiting DDIT4-eEF2k autophagy axis. 2 Inhibition of SIRT1/2 inhibited mTORC1 signaling pathway in NSCLC cells through acetylation of HSPA5 and subsequent activation of ATF4 (activating transcription factor 4) and DDIT4, thereby inducing autophagy and tumor resistance. 44 DDIT4 promoted autophagosome formation and increased autophagy by regulating key effects of autophagy, which had mediated advanced prostate cancer cells resistance to bortezomib. 26 mTORC1 phosphorylated the formation of Unc-51-like kinases 1 (ULK1), Autophagy-related protein 13 (ATG13) and Focal adhesion kinase family-interacting protein of 200 kDa (FIP200) complexes that inhibited autophagy. DDIT4 promoted the formation of ULK1-ATG13-FIP200 complex to induce autophagy through mTORC1. 45
DDIT4 had a wide range of function in tumor cells, and its interacting process also included expression levels of miRNA, 46 methylation of histone H3K4 promoter, 34 and nuclear localization of enhancer of rudimentary homolog (ERH). 47 DDIT4 was the bona fide target of miR-221, while the miR-221/DDIT4/mTORC1 signaling pathway was involved in the growth of tumorigenic murine hepatic progenitor cells. 46 Lysine-specific demethylase1 Inhibitor (ZY0511) up-regulated DDIT4 expression by altering the methylation level of histone H3K4 promoter. 33 Drosophila enhancer of the original homolog was nuclear localization and exerts its biological function through RPS3, RPL19 and DDIT4, among which DDIT4 may promote tumor cell metastasis through ERH. 47
DDIT4 indirectly affects tumor cells by regulating the function of immune cells around the tumor
DDIT4 was highly expressed in the lungs of smoking mice and activated NF-κB to cause alveolar inflammation, oxidative stress, and apoptosis of alveolar septal cells. 48 DDIT4 in neutrophils from patients with active systemic lupus erythematosus (SLE) mediated the release of neutrophil extracellular traps (NET) by promoting cell autophagy. 49 DDIT4 in patients with Familial Mediterranean fever (FMF) was prone to co-centralization with pyrin and nucleotide-binding domains, leucine-rich repeats/pyrin-containing domains 3, leading to autophagy-mediated NET Release and high IL-1β expression. 50 The overexpression of lncDDIT4 in naive CD4+ T cells inhibited IL-17 transcription by increasing activation of DDIT4/mTORC pathway. 51
The accumulation of dysfunctional mitochondria in macrophages may be involved in colitis and inflammatory bowel disease. DDIT4 was a key mediator in that eliminates dysfunctional mitochondria characterized by low membrane potential and a high level of reactive oxygen species. The low level of reactive oxygen species in mitochondria related to DDIT4 in macrophages induced by IL-10 and hypoxia emphasize the metabolic reprogramming of immune cells can affect its cell function. 52 DDIT4 mediated its related immune response through mTORC1 and directly effects with IκBα. 53 Hypoxic TAMs had the characteristics of angiogenesis and immunosuppression and it upregulated the expression of DDIT4 under hypoxic environment. DDIT4-mediated mTORC1 inhibition reduces the level of glycolysis in hypoxic TAMs, and inhibited vascular hyperactivation and the formation of quiescent vascular junctions. 54 DDIT4 knockout TAMs promoted abnormal angiogenesis and metastases. 54 DDIT4 indirectly affects the angiogenesis and metastases of tumor cells through the metabolic reprogramming of immune cells, and it desires for further study.
DDIT4 participates in the tumorigenesis and development of cancer by affecting the function of adipocytes and muscle cells
Functional changes of other normal cells besides the tumor cell is one of the most important characteristics of the cancers, including the metabolic reprogramming of adipocytes and skeletal muscle cells. Fatty-acid synthase (FAS) was up-regulated in many cancers including breast cancer, prostate cancer and ovarian cancer, and the inhibition of FAS activated caspase-8 or to trigger apoptosis in tumor cells via DDIT4. 55 FAS inhibition promoted ATF4-dependent DDIT4 transcription induction and caspase-2 activation, leading to the death of ovarian cancer cells. 56 In DDIT4-/- mouse, the release of ATG4B activity and the blocking of autophagy flux resulted in the accumulation of defective mitochondria, which led to impaired oxidative phosphorylation, depletion of muscle ATP, and poor exercise capacity. 12 Skeletal muscle depletion was a strong prognostic factor in epithelial ovarian cancer, but the exact signaling pathway was blocked. 57 DDIT4 may be involved in the proliferation and apoptosis of tumor cells by affecting muscle cells and adipocytes.
DDIT4 could affect the number and function of adipocytes and skeletal muscle cells. It decreased the number of skeletal muscle cells in the absence of energy via a DDIT4-dependent mechanism coupling mitochondrial respiration and anabolic processes 6 and DDIT4 was involved in limiting muscle loss in glucocorticoid-mediated atrophy. 58 In 3T3-L1 adipocytes, silencing DDIT4 by siRNA led to increased mTORC1 activity and inhibited insulin signaling pathways and adipogenesis. 59 DDIT4 was a target gene regulated by p53, and its expression could increase the degradation of adipocytes. 41 The decreased number of skeletal muscle cells and accumulation of adipose tissue affected the network of inflammatory pathways, insulin resistance and tumor growth. 60 adipocytes may be a barrier to treatment because they are involved in the resistance mechanisms of various breast cancer treatments and mediate tumorigenesis and development through cytokines. 61 At the same time, adipose tissue could mediate breast cancer cell proliferation and participate in resistance hormone therapy through leptin. 62
DDIT4 is an important hypoxia-inducing protein and is involved in oxygen sensing signaling pathways
The expression level of DDIT4 obviously increased under hypoxic conditions. Hypoxia led to increased expression of ataxia telangiectasia mutated (ATM)-dependent phosphorylation of hypoxia-inducible factor 1α(HIF-1α) and its downstream DDIT4/mTORC1 signal excessive activated. 63 Under hypoxic stress, overactivity of reactive oxygen species (ROS)-induced activator protein-1 (AP-1)/c-Jun signal increased DDIT4 protein level and DDIT4 promoter-driven luciferase activity, and deletion of AP-1 bound proximal region of DDIT4 promoter site significantly inhibits the activation of DDIT4 by c-Jun. 64 DDIT4 induction by hypoxia and the related signals were relayed to mTORC1 through different pathways in a tissue-specific manner. In the central region of the liver lobule, DDIT4 could change the expression levels of hypoxia/mTORC1 pathway, while the primary hepatocytes are not related to DDIT4. 65
DDIT4-/- mouse embryonic fibroblasts cells maintained high expression of HIF-1α through ROS. At the same time, HIF-1α induced glycolytic metabolism gave tumor cells a growth advantage under hypoxic conditions. 11 Akt/GSK3 expression increased mitochondrial membrane potential and promoted ROS expression when DDIT4-/- cells exposed to hyperglycemia. 11 However, DDIT4/TXNIP complex expression was sufficient to induce ROS, suppressed ATG4B activity and activated autophagy. 12 At present, the relationship between DDIT4 and ROS is unclear, and further research is needed to explore its accurate signaling pathway.
Current research supported oxygen sensing is a complex signal network, and the study of hypoxia-related protein DDIT4 was a potential visitor in the oxygen sensing pathway. HIF-1 was widely expressed in mammalian cells in a hypoxic culture environment. Erythropoietin (EPO) gene enhancer mediated transcriptional activation in hypoxic cells. 66 The RNA and protein levels of HIF-1α and HIF-1β induced by EPO in hypoxic mammalian cells. 67 Von Hippel-Lindau (VHL) tumor suppressor gene product pVHL played a key role in the degradation of HIF-1. In VHL-defective cells, the expression of HIF-1α subunit was obviously increased, and HIF-1 was continuously activated. 68 The interaction between HIF-1 and pVHL was iron-dependent, and HIF-1 separated from pVHL in cells exposed to iron chelation or cobalt ion environments. 69 In the case of sufficient oxygen, HIF was targeted by E3 ubiquitin ligase containing pVHL, and the hydroxylation of proline requires ferrous ions and oxygen molecules. 70 The interaction between pVHL and specific regions of the HIF-1α subunit is regulated through hydroxylation of a proline residue (HIF-1α P564) by an enzyme we have termed HIF-α prolyl-hydroxylase (HIF-PH). 71 Silencing of prolyl-hydroxylases 2 (PHD2) was sufficient to stabilize and activated HIF-1α in normoxia, providing a HIF-1 dependent auto-regulation mechanism driven by oxygen tension. 72 High expression of DDIT4 was found in the iron-deficient environment, and the increased DDIT4 expression level was quickly reversed after the iron increased. 73 DDIT4 may sense the local oxygen tension of the tissue through HIF, iron deficiency and ROS, and participates in the adjustment of the oxygen perception pathway by changing the expression level of ROS.
Limitations and future perspectives
Our review provides a valuable summary of the function of DDIT4 in cancer, but there are still some limitations in this study. First, our explanation of the contradictory function of DDIT4 in cancer need a better exploration. The current literature involved the multiple roles of DDIT4, including a mitochondria related protein, 6 interactions with various protein molecules, immune and metabolic cell related proteins and participator in the oxygen sensing pathway, while the definition of DDIT4 should be more appropriate. DDIT4 was an important regulatory protein which related to mitochondrial function, 54 and it was involved in the regulation of mitochondrial function in varied fields. However, the current study lacked a certain amount of high-quality literature directly describing the regulation of mitochondrial in cancer.
In the future, DDIT4 may find a balance in the mTORC1, p53, HIF, autophagy, and oxygen sensing signal pathway while DDIT4 will get a unique label in special pathological state at the process of cancer more than as a mitochondrial related protein. As we can see that DDIT4 affected tumor cell metastases through macrophages, 54 and more work is needed to be done because the role of adipocytes, stellate cells and fibroblasts in the malignant behavior of the tumor may be a novel strategy in cancer treatment. The type of DDIT4 gene mutation in different cancer and whether they affected the survival of cancer in the harsh microenviroment.
Conclusions
DDIT4 as a mitochondrial related protein was involved in the mTORC1, p53, HIF, autophagy, and an oxygen sensing signal pathway. And it is a tumor associated protein as well as a metabolic and immune related protein, which is highly expressed under the stress conditions such as chemotherapy, hypoxia, and DNA damage and it shows obvious potential value in cancer treatment.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (no. 81572442 and no. 81800070), the Natural Science Foundation of Fujian Province of China (no. 2020J01961) and Medical innovation in Fujian Province (2019-CX-27). These funding sources played key supportive role for bioinformatics analysis.
Author biographies
He has participated in the National Natural Science Foundation of China and the Natural Science Foundation of Fujian Province. Three papers were published as the co-author or correspondent author index by SCI.