
Abstract
Nest building behavior has been intensely applied as a parameter for severity assessment in mice. In contrast, only a limited number of studies have reported nest building data from rats. Here, we assessed nest building in rats in two different facilities addressing the hypotheses that the vendor, previous experience with the nesting material as well as sex of the rats has an impact on the performance. Data from two study sites and three raters were compared to obtain information about the robustness of nest complexity scoring. The findings demonstrate a generally poor nest building performance in rats with a pronounced day-to-day fluctuation, and site-specific differences. Application of a newly developed scoring system resulted in an intermediate inter-rater reliability. Previous experience with the nesting material did not exert a consistent impact on nest complexity scores. Sex differences proved to depend on vendor and animal facility without consistent findings supporting a superior performance in female or male rats. In conclusion, our findings argue against a robust and consistent influence of sex and familiarity with the nesting material. The comparison between facilities suggests that local conditions need to be considered as influencing factors, which should be explored in more detail by future multicenter approaches. Considering the day-to-day fluctuation and the intermediate inter-rater reliability, we highly recommend to base nest complexity evaluation on means from several subsequent days analyzed by a group of experienced raters.
Nest building behavior has been intensely validated as a well-being parameter in laboratory mice.1–4 This species shows a strong natural motivation to engage in nest building activity, probably to create a cage subarea as a shelter with an optimized microclimate concerning temperature and light exposure. Assessment of nest complexity and level of soiling revealed that these parameters can be significantly affected by distress and pain in mice, and can therefore be useful for severity assessment.1,2,5–13
In rats it has been reported that although rats preferred cages with nesting material,14–16 they did not construct complex nests when coarse paper strips (Enviro-dri®) were offered as enrichment. 16 This led van Loo and Baumans to hypothesize that in rats nest building behavior is not genetically determined, but needs to be learned. 17 To confirm this hypothesis they compared nest building in female and male Wistar rats (U:WU) with a different history of exposure to two different types of nest material (Kleenex® tissues or Enviro-dri®). Data from this study revealed that the older the rats at first exposure to Enviro-dri®, the poorer the nest building performance, although the nest-building performance improves over time. 17
Despite this report in 2004, which suggested an importance of learning young in rats and recommended nesting material for enrichment in rats, and despite the fact that nest building has been applied as a well-being parameter in mice, only few studies have assessed nest building behavior in rats except from studies focusing on maternal behavior.18,19 This might be related to problems with nest building performance in rats. In line with this assumption, we also faced respective problems, when we initiated studies with nest building assessment in a research consortium focused on evidence-based severity assessment in rats. To explore possible reasons for these difficulties we assessed nest building in rats in two different facilities in a systematic manner. We addressed the hypotheses that the vendor, previous experience with the nesting material, as well as sex of the rats have an impact on the nest building performance. In addition, we compared data between study sites and between raters to obtain information about the robustness of nest complexity scoring. For these analyses, Sprague Dawley rats have been selected as the most frequently used rat breed worldwide.
Materials and methods
All investigations were conducted in line with the German Animal Welfare act and the EU directive 2010/63/EU. The health status of the animal facilities of the two institutes met the FELASA guidelines. 20
Animals and experimental design
Researchers at Ludwig-Maximilians-University, Munich (LMU) and Hanover Medical School (MHH) performed experiments to investigate the influence of (i) vendor, (ii) familiarity with the nesting material Enviro-dri® (Claus GmbH, Limburgerhof, Germany), and (iii) sex on nest building performance. The choice of the nesting material has been based on pilot studies and previous experience of the research consortium.21–23 Also based on pilot data, a power analysis was carried out and revealed a necessary group size of n = 5 pairs. This minimum number was considered in all subgroups of animals.
For the “naïve” group (i.e. unfamiliar to Enviro-dri®), rats were obtained from Charles River Laboratories (Sulzfeld, Germany; CR) where the nesting material is conventional pulp (Tork Standard Papierwischtücher two-layer, Mannheim, Germany). These rats were tested either at MHH or LMU, in the following termed MHH/CR/pulp and LMU/CR/pulp. For the “familiar” group, rats that were raised with the nesting material Enviro-dri® from birth onward were either obtained from Envigo (ENV) at LMU or bred in-house as F1 from SD rats purchased from CR at MHH. These groups were termed LMU/ENV/Enviro or MHH/CR-F1/Enviro.
At LMU, in total 20 virgin female and 20 virgin male Crl:CD (SD) rats were investigated. Male (n = 10) and female (n = 10) rats were obtained from Envigo (the Netherlands) and CR at approximately 12 weeks old. Rats were pair-housed by sex and vendor, i.e. familiarity with nesting material.
At MHH, in total 28 virgin female and 26 virgin male Crl:CD (SD) rats of approximately 12 weeks were used with 14 female and 12 male rats from CR, and 14 female and 14 male rats from the F1 generation (parental generation from CR) bred in the Central Animal Facility of MHH. Rats were pair-housed by sex and familiarity with nesting material.
For additional information regarding the animal husbandry, see supplemental material.
All animals were housed for other studies or training purposes, as approved by the government of Upper Bavaria (license number ROB-55.2-2532.Vet_03-15-11 and ROB-55.2-2532.Vet_02-14-120) and the Lower Saxony State Office for Consumer Protection and Food Safety, LAVES (AZ 16/2315, 15/1933 and 17/2477).
Rats were housed in pairs and received 28 g of Enviro-dri® once weekly upon cage cleaning. Nesting material was placed in the back left corner of the cage. The amount of 28 g was chosen based on the outcome of pilot studies testing 14, 21, and 28 g for pair-housed rats. The placement of the material was standardized for all facilities with the only criterion for our decision that the corner should be located opposite to the feeding rack.
Nest complexity scoring
Every morning between 08:00 and 09:00 photos of the nests were taken for image-based scoring, including at least one side view and one top down view. These photos were then arranged as one photo assembly for each pair of rats and day.
For scoring of the nest building behavior we first followed a score modified according to van Loo and Baumans (see Table 1).
17
After analysis of the scores by two raters, which showed tremendous disparities, a discussion among the collaborating groups revealed that the nest complexity and shapes were not described in unambiguous detail by the score levels. Review of nest complexity scores in mice indicated the use of more detailed grading systems with 4–6 levels.2,24,25 Following this concept, a more detailed scoring system was developed (Figure 1, Table 1). With this score, the complexity of 120 nests (based on two to three photos per nest) from the different animal facilities was described in detail by two experienced raters, which subsequently agreed on what score should be given and how this score should be described best.
New nest score. A: score 0 = the nest material is almost untouched. B: score 1 = nest material is touched and distributed over more than half of the cage's floor area without a visible nesting area. C: score 2 = a nesting area is clearly visible, which is smaller than half of the cage base area (i.e. less than 1/5 cage Type IV Makrolon cage), the nest is flat with no visible indentation, the edges of the nest may have a frayed shape. D: score 3 = the nest area is clearly defined and delineated and the nest has either an appreciable height (at least 1/5 of the cage height) with a minimal indentation or is flat but with a prominent indentation. E: score 4 = the nest has both an appreciable height (at least 1/4 of the cage height) and a prominent indentation. Nest scoring scheme.

The robustness of this scoring system was tested by three raters (referred to as raters 1–3) from both institutes not aware of the group allocation (including history of nesting material, vendor, and sex) and origin of the animals. Raters were first provided with the training set of 120 nests described above. The training set had to be passed with an accuracy level of at least 80%. Thereafter, all raters analyzed a comprehensive set of 861 nests from LMU and MHH.
The elaborate score has been applied for all data sets provided and discussed in this manuscript. Please note that the data sets from individual raters are illustrated in Supplementary figures 2–4.
The evaluation was focused on weeks two and three following arrival in the animal facility or postnatal weeks 10–12 in the F1 generation reared at MHH (referred to as weeks 1 and 2 of evaluation). Animals at MHH that were purchased from CR (i.e. not reared at MHH with Enviro-dri®) were not exposed to nest material during the first week after arrival. Animals at LMU received nesting material from the day of their arrival. Therefore, it was possible to perform an additional analysis of nest building performance at the first days (days 1–4) following arrival at LMU (referred to as week 0).
Statistical analysis
GraphPad Prism (Version 5.04; GraphPad, La Jolla, CA, USA) and R version 3.3.2 were used for statistical analysis. 26 A two-way analysis of variance (ANOVA) with a Bonferroni post-hoc test was used for the comparison of different familiarity with nesting material and sex, and for the comparison of vendor and sex for days 1–4 after arrival at LMU. A repeated Three-way-ANOVA followed by a post-hoc test adjusted for multiple testing using a Benjamini and Hochberg correction was used for comparing the effect of site, sex and the different time windows within animals from the same vendor (CR). Data represent mean with standard deviation (SD) of the median of days 4–6 of all raters (main figures) or for individual raters (see supplementary figures). Mean nest scores were calculated based on average nest scores of days 4–6. R version 3.3.2 was used to create the timelines in Supplementary figure 1. 26 The R package “irr” was used to calculate the intraclass correlation coefficient (ICC) and Kendall's coefficient of concordance W. 27 Significant differences between groups are shown as asterisks (* p < 0.05; trend is shown as asterisk in brackets (*) < 0.01).
Results
Course of nest complexity scores
Following exposure to new nest material, the daily scores increased at least to some extent during testing, but showed a relatively high level of fluctuation (Supplementary figure 1). Mean nest scores of days 4–6 of each week were used to compare overall performance; the selection of these days was based on previous experiments in which it was observed that nest quality plateaued following day 4 and often decreased again at day 7. 22
Inter-rater reliability scores
Using a scoring scheme modified according to van Loo and Baumans, 17 the different raters reached an inter-rater reliability score, as measured by the ICC of 0.54 (p < 0.001) and measured by the Kendall's coefficient of concordance W of 0.60 (p = 0.0475). Using the more detailed training and scoring scheme the inter-rater reliability score reached an ICC of 0.79 (p < 0.001) and Kendall's W of 0.63 (p < 0.001). Results from individual raters can be found in Supplementary figures 2–4.
Comparison between sites, origin, and sex
A full comparison of nest building scores was possible using the CR groups, where data is available for both sites (Figure 2), both sexes, and the two subsequent weeks following introduction of the nesting material. The three-way ANOVA indicated an overall significant difference between the two sites (F(1,37) = 5.884, p = 0.0203) and an overall difference between weeks 1 and 2 (F(1,37) = 5.328, p = 0.0267). Individual comparisons revealed that a difference between sites was only evident for female rats reaching higher nest complexity scores at LMU compared with MHH in week 2 (p = 0.0141). Further comparisons confirmed a significant difference between weeks 1 and 2 (p = 0.0449) in females at LMU. Lastly the only sex difference found was between the MHH groups in the second week, with males building more complex nests as compared with females (p = 0.0380).
Comparison of nest scores of Charles River rats at two different sites (LMU and MHH), two sexes and two time points. Shown are the mean nest scores for days 4–6 (+/- SD) of each week after receiving new nest material. Significant differences between groups are indicated by asterisks (three-way ANOVA with post-hoc test adjusted for multiple testing using Benjamini and Hochberg; * p < 0.05).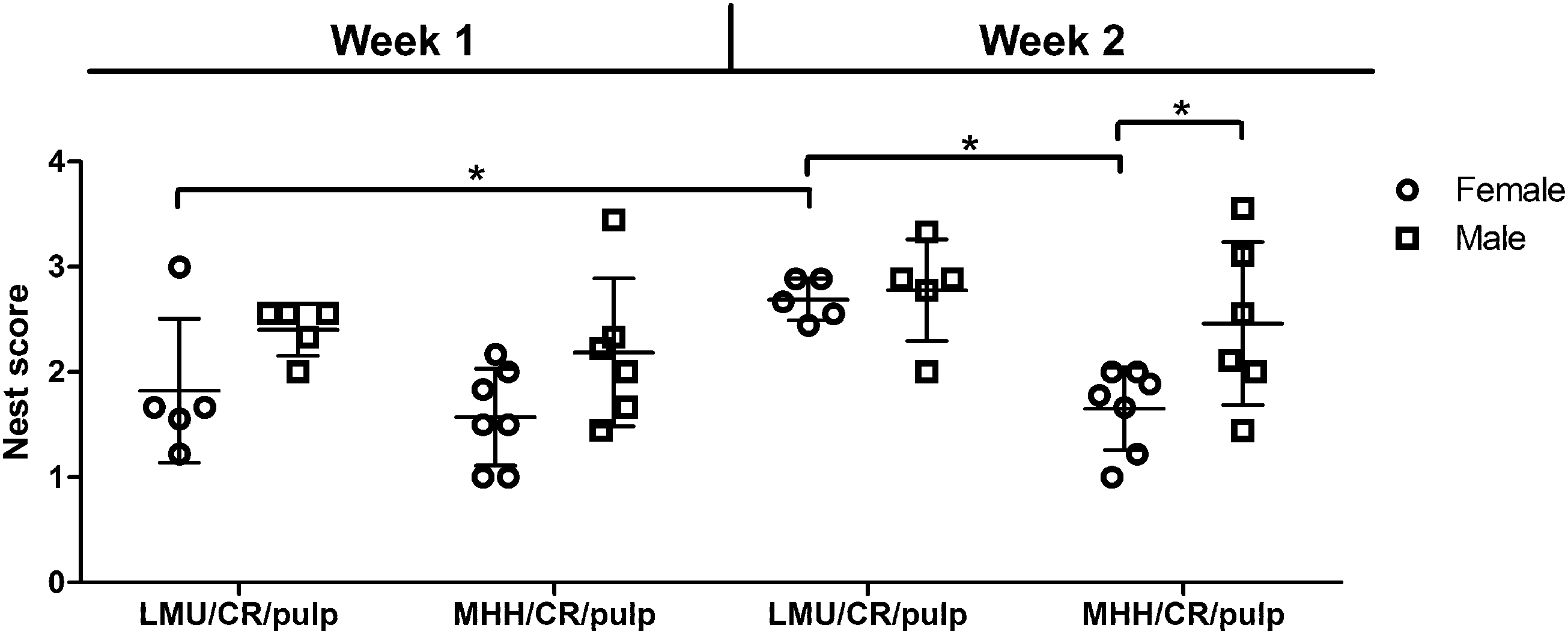
Scores of individual raters were characterized by a relatively high level of variation, as already indicated by ICC analyses. Thereby, results of raters 2 and 3 proved to be more consistent than results of rater 1 (Supplementary figure 2(a) to (c)).
Comparison of rats at LMU did not reveal significant differences between animals supplied by Envigo and CR (Figure 3(a)). In order to test whether early exposure to Enviro-dri® has an impact on nest building performance in animals of the same breed, nest scores were compared between animals from CR (MHH/CR/pulp) and animals reared with Enviro-dri® (MHH/CR-F1/Enviro) at MHH. The respective comparison did not indicate a significant difference (Figure 3(b), Supplementary figure 3(d) to (f)). Direct comparison between females and males only revealed a tendency for better nest building in females as compared with males for two of the three raters (Supplementary figure 3(a) and (b), LMU/ENV/Enviro female against LMU/ENV/Enviro male: p < 0.1). This was confirmed by mean scores of all raters (Figure 3(a), LMU/ENV/Enviro female against LMU/ENV/Enviro male: p < 0.05). Surprisingly, statistical analysis revealed a significant higher complexity of nests in males as compared with females in the MHH/CR/pulp group (Figure 3(b) and Supplementary figure 3(d) to (f), sex (F(1,23) = 5.004, p = 0.035), MHH/CR/pulp female against MHH/CR/pulp male: p < 0.05).
Mean nest score comparison of vendor and sex for groups tested either at LMU (a) or at MHH (b). Shown are the mean nest scores for days 4–6 (+/- SD) after receiving new nest material in second week of nest scoring. At LMU rats from Envigo (LMU/ENV/Enviro) were compared with rats from Charles River (LMU/CR/pulp) (a), at MHH rats from Charles River reared at MHH (MHH/CR-F1/Enviro) were compared with rats from Charles River (MHH/CR/pulp) (b). Significant differences between groups are indicated by asterisks (two-way ANOVA with a Bonferroni post-hoc test; * p < 0.05).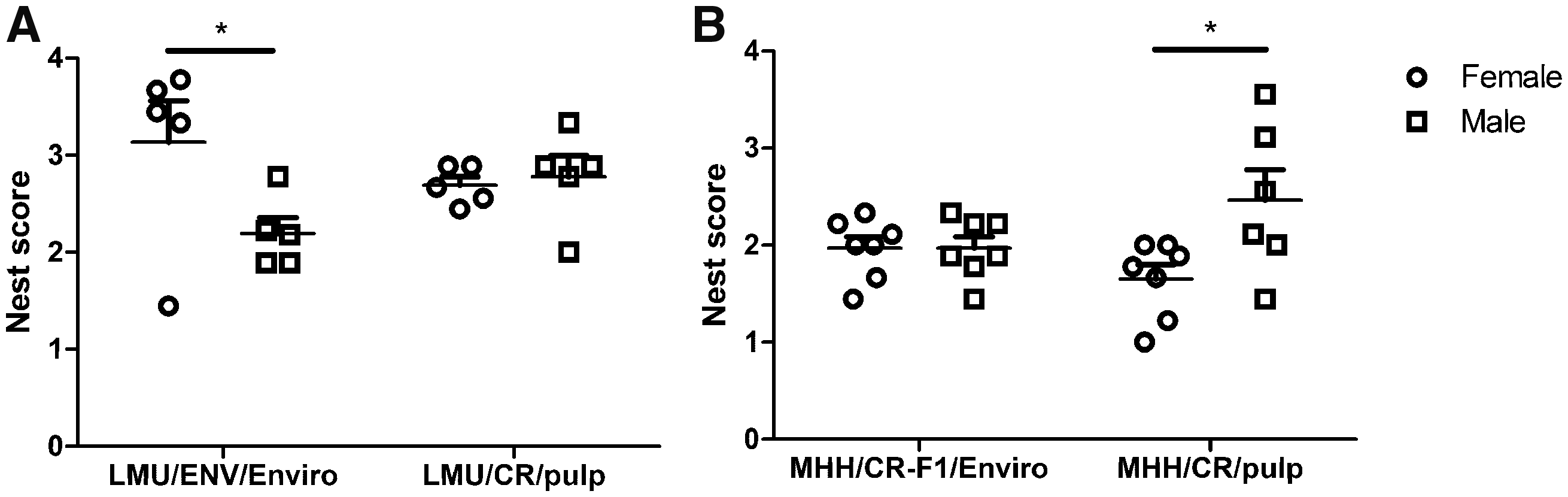
Course of nest complexity following arrival or first exposure to nest material
In the animal facility at LMU, animals were offered nest material immediately following arrival in accordance with the local regulations. During the first days of exposure (mean scores of days 1–4 – week 0) a pronounced group difference proved to be evident with nest complexity scores of female SD rats from Envigo, i.e. the LMU/ENV/Enviro group, exceeding scores in all other groups by rater 1 (Figure 4 and Supplementary figure 4 for raters 1–3, interaction (F(1,16) = 25.39, p < 0.001); sex (F(1,16) = 7.087, p = 0.017); vendor (F(1,16) = 17.5, p < 0.001), LMU/ENV/Enviro female against all others p < 0.001). Shown are the mean nest scores for days 4–6 (+/- SD).
Comparison of mean nest scores for days 1-4 after arrival at LMU Munich for male and females of Envigo (LMU/ENV/Enviro) and Charles River (LMU/CR/pulp) pairs. Increased nest scores were found in female rats from Envigo as compared with all other groups. Significant differences between groups are indicated by asterisks (two-way ANOVA with a Bonferroni post-hoc test; * p < 0.05).
Discussion
To explore the suitability of nest complexity scoring as a basis for severity assessment in rats we compared nest building performance of virgin male and female pair-housed SD rats from different vendors and with different familiarity with the nesting material Enviro-dri® in two animal facilities. When considering that only scores ≥ 3 indicate a more complex nest structure, it is notable that respective median score levels were only rarely reached in the facility of MHH, and only on single days at LMU. Only for female SD rats purchased from Envigo and kept at LMU the median nest score determined at the majority of testing days reached levels above 3. These data indicate a generally poor nest building performance of rats in line with previous descriptions by Manser and colleagues. 15
In mice different experimental paradigms that cause distress and pain can result in a reduction of nest complexity scores.1,2,5–13 Therefore, Jirkof suggested that alterations in nest building behavior and changes in burrowing behavior along with specific disease markers might serve as a valuable indicator of well-being in mice. 2 The poor nest building performance in rats, however, raises doubt on a comparable suitability of nest building behavior as a generalizable severity assessment parameter. Nevertheless, nest building in female SD rats purchased from Envigo, i.e. rats with the highest nest complexity scores in the present study, has been previously reported as suitable for severity assessment in a rat model for epilepsy. 22
Another drawback of nest scoring seems to be the high inter-rater variability, which constitutes a general problem of “qualitative scoring systems”. With the application of a simple scoring system following the description by van Loo and Baumans, 17 we observed tremendous discrepancies in the scores given. Thus, a more detailed scoring system was developed, which, together with a training set, improved the inter-rater reliability at least to some extent. Nevertheless, even this more detailed grading system resulted in rather intermediate levels of inter-rater reliability, which poses problems with regard to the reproducibility of data sets. Therefore, in line with a statement by Jirkof , 2 we recommend to carefully consider inter-rater variability and to use the mean values of scores provided by a group of raters in order to guarantee a higher robustness of findings. Moreover, considering the progressive increase during the first days and day-to-day fluctuations, we strongly recommend to use the mean of days with rather stable nest complexity.
In mice a high level of motivation for nest building has generally been reported in both sexes.28,29 In rats, previous studies also did not find differences between males and females, although often only one sex was tested.15,17,30,31 Overall, we also found no consistent differences between males and females, although nest complexity scores at different locations may differ between sexes. Male SD rats purchased from CR constructed more complex nests than female rats at MHH, whereas nest building performance was better in female than in male rats from Envigo at LMU. Thus, relevant differences between sexes can occur which should be considered in the study design.
It has been suggested that the nest building performance of rats largely depends on their previous experience and that exposure to a specific nest material during their development results in an improved nest building performance later on. 17 This theory seems to be confirmed by highest nest complexity scores observed in female SD rats from Envigo that grew up with Enviro-dri® as nesting material. However, nest building in SD rats purchased from CR, which grew up with standard pulp as nesting material, did not differ from those of the F1 offspring raised up with Enviro-dri® at MHH, e.g. rats with the same genetic background. This argues against a relevance of previous exposure to the specific nest material. In fact, the maximum individual nest scores in male SD rats at MHH were even higher in those animals that grew up at CR without exposure to Enviro-dri®.
Taken together, these data do not support the “importance of learning young” as suggested by van Loo and Baumans. 17 On the other hand, the progressive increase of nest building performance observed in female SD rats from CR at LMU indicate that the impact of the history and experience should not be neglected, although the level of its impact seems to depend on the local husbandry conditions, sex of the animals, and other factors.
Finally, an inter-site comparison between SD rats supplied by CR showed higher nest building performance especially of female SD rats at LMU despite the fact that the room climate and light conditions have been set to comparable standards according to European guidelines in all animal facilities with continuous daily control. Although rats have a better body surface to weight relation, and therefore less pronounced needs in terms of support for thermoregulation, an impact of the surrounding air temperature on nest building activity in rats has already been described in 1927. 32 Moreover, studies in mice indicated a major influence of the room temperature. 33 Thus, we speculate that differences in the microclimate related to the local technical system might have resulted in differences in the felt temperature with an impact on the motivation for nest building. In this context, it is of interest that the air exchange rates at LMU were considerably higher than at MHH. Differences in the distance of the transport from the commercial vendor to the animal facility, as well as in the handling of animals by personnel in the facilities may also constitute important variables that can significantly influence behavioral patterns. To determine influencing factors on the robustness of nest building data it may be of future interest to assess nest building performance in rodents in a multi-center study, which has been shown to be a valuable approach to gain information about robustness, reproducibility, and confounding factors.34,35
In conclusion, our findings demonstrate a generally poor nest building performance in virgin pair-housed rats as compared with mice. The data argue against a robust and consistent influence of sex and of familiarity of the specific nesting material. The comparison between facilities suggests that local conditions need to be considered as influencing factors, which should be explored in more detail by multicenter approaches.
The day-to-day fluctuation of scores and the intermediate inter-rater reliability results in the recommendation to base nest complexity scoring on means from several subsequent days analyzed by a group of experienced raters.
Supplemental Material
Supplemental material for Nest-building performance in rats: impact of vendor, experience, and sex
Supplemental Material for Nest-building performance in rats: impact of vendor, experience, and sex by Kerstin Schwabe, Lena Boldt, André Bleich, Roelof Maarten van Dijk, Simeon Oscar Arnulfo Helgers, Christine Häger, Marta Nowakowska, Ann-Kristin Riedesel, Katharina Schönhoff, Birgitta Struve, Jürgen Wittek and Heidrun Potschka in Laboratory Animals
Footnotes
Acknowledgments
The authors thank Sieglinde Fischlein, Verena Buchecker, Sabine Vican, Katharina Gabriel, Uwe Birett, Monika von Iterson, and Daniel Ahrens for their excellent technical assistance and Ute Lindauer, Annika Bach, and Ekaterina Harder for their input and support in developing the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The study was funded in part by the German Research Foundation (Deutsche Forschungsgemeinschaft - DFG) FOR 2591 Consortium. Reference numbers are listed in the supplemental material.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.