
Abstract
Mice are widely used in studies investigating the effect of diet on metabolic risk factors, such as lipid profiles and plasma glucose levels. An important factor that is usually not taken into account is the biological age of the experimental models. The up-to-date identified experimental confounders do not cover all the parameters that may affect the results of animal studies. The aim of this study was to investigate the effects of a high-fat diet on the metabolic profile, hepatic and renal function in mice of differing ages. For this purpose two groups of male C57BL/6J mice were used, consisting of 10-week-old mice and 54-week-old mice in each group. Both groups followed identical high-fat diets for 12 weeks. The younger mice showed smaller increases in body weight, serum total cholesterol, glucose and urea levels while they had higher increases in high-density lipoprotein cholesterol levels than the older mice. Our results indicate the necessity to consider an experimental animal’s age as a confounding factor when researching or interpreting metabolic studies. Age adjustment is warranted in all animal research while a uniform approach regarding the age of the animal models should be applied in experimental studies.
The aging process is associated with progressive insulin resistance, 1 inhibition of lipolysis 2 and less hepatic control of glucose production and triglyceride secretion.3,4 In addition metabolic disorders, such as dyslipidaemia and hyperinsulinaemia, increase with age. 5 Thus, when investigating the metabolic impact of dietary or other interventions using experimental animal models, it is essential to take into consideration the age of the animals in order to interpret the results. 6
The aim of this study was to test the impact of age on the changes in lipid and glucose homeostasis in young and middle-aged mice receiving a high-fat diet (HFD).
Materials and methods
Dietary analysis of the diet with 45 kcal% fat from lard.
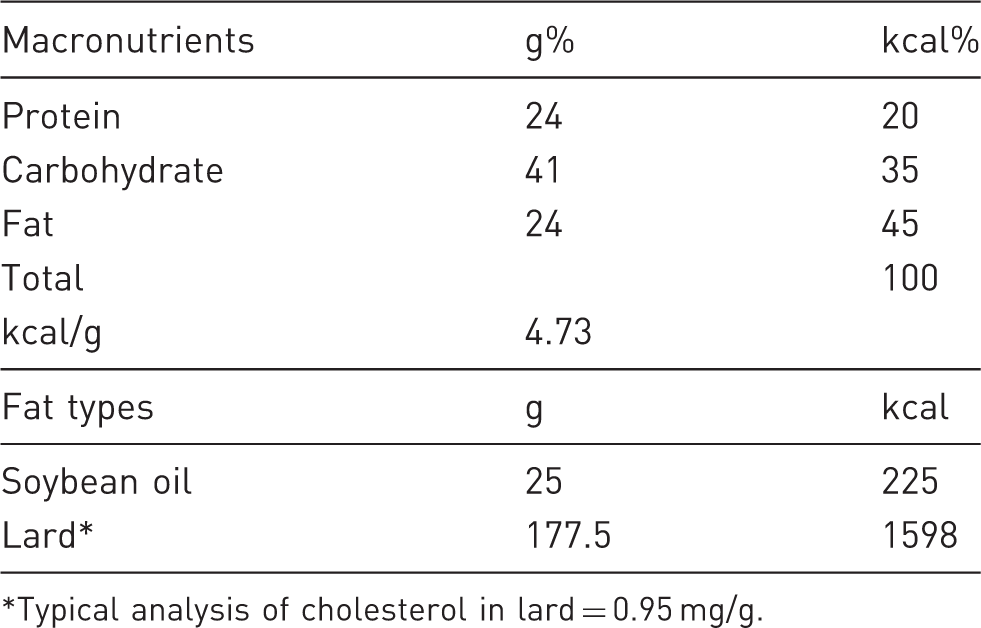
Typical analysis of cholesterol in lard = 0.95 mg/g.
Blood samples were collected at the beginning and at the end of the experimental period (at 09:00 h, after a 12 h fast), from the medial retro-orbital venous plexus under light ether anaesthesia. Serum concentrations of total cholesterol (TCHOL) and triglycerides were determined using an enzymatic PAP commercial kit (‘biosis’, Biotechnological Applications, Athens, Greece), while high-density lipoprotein (HDL) cholesterol and glucose levels were determined by enzymatic photometric methods. Serum samples for determination of serum glutamic pyruvic transaminase (SGPT), serum glutamic oxaloacetic transaminase (SGOT), uric acid, urea, total serum proteins and creatinine levels were measured in the same batch using an automated analyzer (Technicon RA-XT, Technicon, Dublin, Ireland) and commercially available kits. Body weights were recorded during the whole study.
The baseline levels and the percentage changes of the above parameters from the baseline were compared between groups, using the Student’s t-test for the normal distributions or the non-parametric Mann–Whitney’s U-test for the non-normal distributions. Statistical significance was set at P < 0.05 (two-tailed).
Results
No differences were recorded in baseline glucose, TCHOL, HDL cholesterol, SGPT, uric acid, urea and creatinine levels; while baseline triglyceride (P = 0.002), SGOT (P = 0.005), total proteins (P = 0.002) and body weight levels (P = 0.017) differed between the two groups. As compared with the older mice (initial body weight, mean ± SD: 23.25 ± 1.83 (g); final body weight, mean ± SD: 39 ± 5.45 (g)), the 10-week-old mice (initial body weight, mean ± SD: 21.25 ± 1.48 (g); final body weight, mean ± SD: 30.87 ± 2.47 (g)), had a lower percentage increase in body weight levels from the baseline (mean ± SD: 0.680 ± 0.232 (g) versus 0.456 ± 0.12 (g); 54-week-old mice versus 10-week-old mice, respectively, (P = 0.032)). The percentage increase in serum glucose levels (P = 0.001) and in total cholesterol levels (P = 0.001), following high-fat feeding, were also lower in the 10-week-old mice as compared with the older animals. There was a significant increase from the baseline in the percentage change of HDL cholesterol levels in the young mice compared with the 54-week-old mice (P = 0.001) (Figure 1). No difference was recorded in the percentage change in triglyceride levels (P = 0.462). The young mice presented a lower percentage increase from the baseline in serum urea levels (0.022 (0.061); median (interquartile range) (mg/dL)) compared with the older animals (0.367 (0.152); median (interquartile range) (mg/dL)) (P < 0.001), while the two groups did not differ concerning the percentage changes in serum hepatic enzymes and in the other parameters related to the renal function (SGOT: P = 0.401; SGPT: P = 0.315; uric acid: P = 0.745; creatinine: P = 0.551) (data not shown).
Percentage changes from baseline in serum glucose levels (mg/dL), total cholesterol levels (mg/dL) and high-density lipoprotein (HDL) cholesterol levels (mg/dL) in the two studied groups (10 and 54-week-old mice).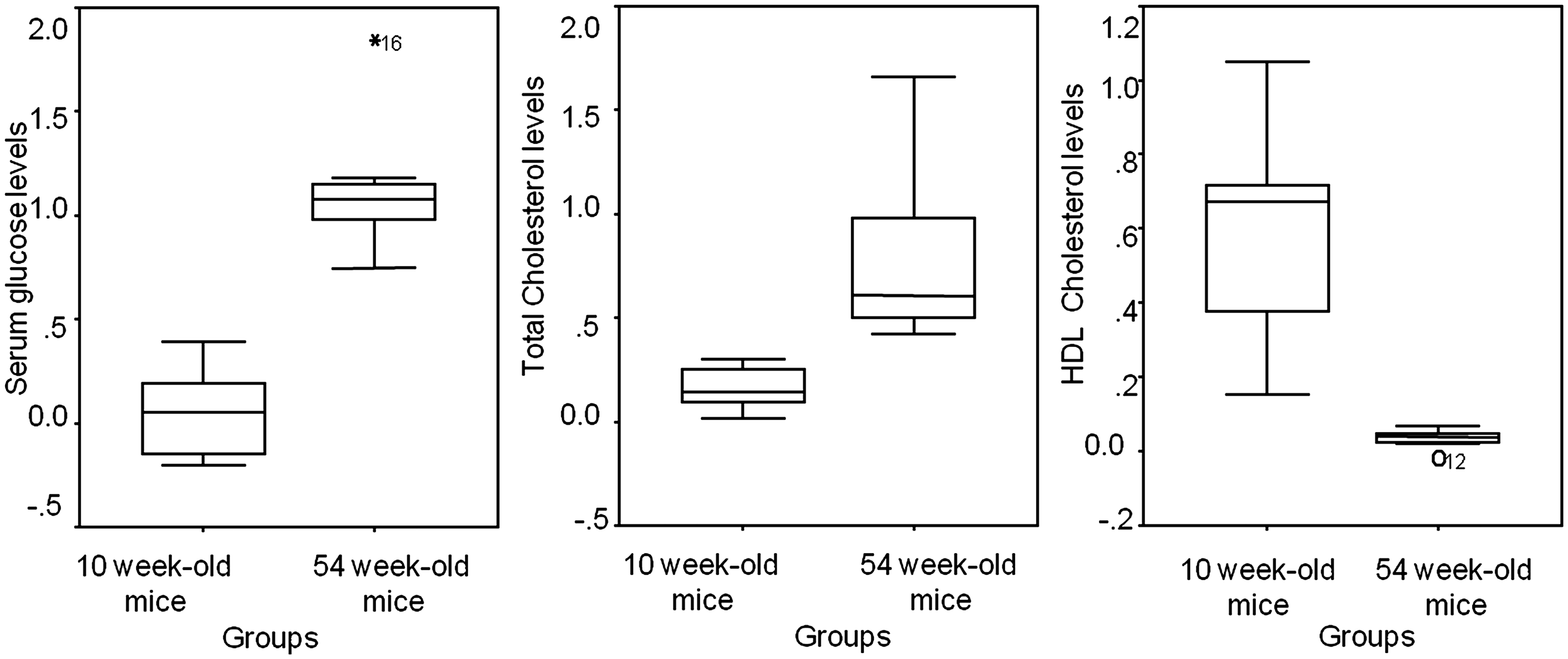
Discussion
HFD feeding may lead to different metabolic responses in C57BL/6J male mice, 7 not only due to changes in the quantity and quality of dietary fat, 8 or genetic variability,9 but also due to the age of the experimental animals.10–12 Middle-aged mice appear to be more susceptible to developing insulin resistance while gaining more weight under HFD feeding compared with young animals. 13 Tschen et al. concluded that young mice resist the development of HFD-induced hyperglycaemia better. 14 Among primates and rodents, older animals are reported to develop more extensive atherosclerotic damage than younger animals when both groups are fed an atherogenic diet. 15
In this study we investigated in detail the impact of age on different blood parameters related mainly to metabolic pathways in a common murine model of diet-induced hypercholesterolaemia and metabolic syndrome. Apart from lipid and carbohydrate markers, we decided to examine the age-related modifications caused by HFD feeding in some important parameters of renal and hepatic function related to organismal aging. The purpose of our work was to indicate that experimental animal studies, performed under ‘controlled’ conditions and thus having minimum confounders, in comparison to clinical studies, must at least be uniform and comparable. Besides the commonly identified confounders, the animal’s age is of the outmost importance especially given all the recent research activity on aging and its relationship with chronic diseases.
We included in our study two different age groups of mice, young adult (10 weeks old) and middle-aged (54 weeks old) mice, based on previous studies.13,15,16
HFD induced more severe aberrations in body weight gain in older animals. Nishikawa et al., 16 reported that HFD feeding increases body weight and fat–body weight ratio while decreasing glucose tolerance, as compared with a control diet, to a greater extent in younger mice than in middle-aged mice. The authors based their results on respective control groups, and considered that these differences were due to the older normal fat-fed animals being already basically obese to some extent.
TCHOL, glucose and urea serum levels were more affected in older mice as compared with the younger animals in our study. Percentage change in HDL cholesterol levels was increased in younger animals although their TCHOL percentage increase was lower. This can be explained as being due to more efficient cardiovascular protection and reduced severity of metabolic disturbance in younger animals. 15
Collins et al. reported that middle-aged LDLR−/− mice developed worse metabolic syndrome, diabetes, and atherosclerosis than 3-month-old LDLR−/− mice when both were fed an HFD. Although serum glucose, triglyceride and HDL cholesterol levels were more disturbed in the older animals, in contrast to our results, serum TCHOL levels and body weight gains did not differ between the two studied groups of mice. 15
Regarding serum urea levels, aging aggravated renal function parameters in apolipoprotein E-deficient hypercholesterolaemic mice, 17 which is similar to our findings.
Although the liver has a remarkable ability to regenerate and respond to stress, these capabilities are restricted with aging. 18 Thus, we expected that young mice would adapt better to diet-induced metabolic alterations regarding serum hepatic function biomarkers. Surprisingly, serum hepatic enzyme modifications caused by HFD feeding in our study did not appear to be affected by age.
Impaired adaptive and homeostatic mechanisms leading to susceptibility to environmental or internal stresses are associated with old age in most species. 19 However, how and when these changes are phenotypically expressed is not yet clear. Overall, the liver in older mice exhibits decreases in growth/proliferation pathways and metabolism that appear through increased oxidative stress markers, as well as increases in inflammatory and stress response genes. 20 Such changes are associated with significant impairment of many hepatic and renal metabolic and detoxification activities, 20 which may however not be present in serum liver and renal markers of metabolism. 21 The same changes were also observed in this study regarding serum creatinine and uric acid levels and hepatic enzymes serum concentrations. Conformation of the aging processes in hepatic and renal tissues would thus require further investigation, e.g. oxidative stress and inflammatory hepatic status assessments.
In total, the above findings provide proof of the importance of the age factor in biomedical studies investigating metabolism and HFD feeding. Also, age should be considered in experimental animal studies in order to establish reproducible and comparable research data.
One of the limitations of this study is the lack of untreated control groups, which are necessary for eliminating alternate explanations of experimental results. Thus, further experiments are necessary in order to quantify accurately the effects of age in both treated and untreated control mice. Previous studies have shown that under normal fat feeding, younger mice gain proportionally more weight than older mice for a nine-week experimental period, 16 due probably to the normal growth processes. It is interesting that in our study with high-fat feeding, this relation was exactly the opposite. A possible explanation may be that the metabolic changes caused by the HFD overrode the normal changes in metabolic parameters caused by the physiological growth processes, resulting in further disturbances in the older animals.
Even though there is no accurate metabolic prediction for an aging murine model the quantification of the observed physiological organismal decline may contribute to more efficient evaluation of scientific outcomes. Recording the changes that occur in naturally aging mice and possibly producing an algorithm predicting all the age-related metabolic modifications may lead to a new era of tightly controlled, accurate and efficient animal experiments.
Footnotes
Declaration of interests
No conflicts of interest are declared.
Funding
We received no financial support for the research and/or authorship of this article.
Acknowledgements
We wish to thank Panagiotis Tsakiropoulos and Nikos Tsakiropoulos for their kind assistance in laboratory techniques. We would also like to thank Kosmas I Paraskevas for his assistance in the preparation of this manuscript.