
Abstract
The cell surface glycoprotein CD44 is involved in active cell movement, cancer metastasis, and morphogenesis. We studied its expression in fetal human skin using an antibody specific for CD44v3 and another that recognizes all CD44 forms. In embryonic and early fetal skin, only cells with dendritic morphology expressed CD44. The first keratinocyte expression of CD44 occurred in the basal cells on the eleventh week. Later, the suprabasal cells also turned positive, whereas periderm and the terminally differentiated cells remained negative at all stages. Maturation of the early mesenchyme towards dermis at the eleventh week was associated with an increase in the number of CD44-positive cells, and later the fibrous extracellular matrix also became CD44-positive. During hair induction, the epithelium showed a transient downregulation of CD44. Later, the follicular cells regained CD44. Cells in the primordial dermal papilla displayed a continuously strong signal. The sweat gland anlagen showed faint CD44 positivity. Exon 3 was present in the CD44 of keratinocytes and their derivatives but was absent in dermis. CD44 expression in human fetal skin is a relatively late event, associated with maturation and adult-type differentiation both in epidermal keratinocytes and in dermal fibroblastic cells.
Adult skin is one of the major organs that show high expression of CD44 under normal tissue homeostasis, both in the epithelial and dermal cells (Brown et al. 1991; Wang et al. 1992). Furthermore, the major CD44 isoform in adult epidermis is a specific large variant (epican) (Haggerty et al. 1992; Tuhkanen et al. 1997), a heparan sulfate proteoglycan rarely expressed in other normal tissues. The heparan sulfate chain resides in exon v3 and may serve in growth factor (especially FGF) binding and presentation (Bennett et al. 1995; Sherman et al. 1998).
Adult epidermal keratinocytes show distinct changes in CD44 expression during their differentiation from basal cells to corneocytes (Wang et al. 1992; Terpe et al. 1994; Seelentag et al. 1996; Tuhkanen et al. 1998), with closely related changes in the content of hyaluronan (Wang et al. 1992). The differentiating cells in adult hair follicles also undergo major alterations in CD44 and hyaluronan (Wang et al. 1992). When the normal differentiation is disturbed, e.g., in psoriasis (Tammi et al. 1994) and squamous cell cancer (Hirvikoski et al. 1999), both CD44 and HA are locally lost from the cell surface. In transgenic mice expressing antisense CD44 under a keratin promoter, epidermal growth and differentiation are severely disturbed, resulting in impaired wound healing (Kaya et al. 1997). One of the findings in these transgenic animals was the apparent loss of epidermal hyaluronan into the underlying dermis (Kaya et al. 1997), suggesting that CD44 is necessary for the retention of HA in epidermis. Indeed, CD44 was recently shown to bind and organize hyaluronan in the pericellular matrix of epidermal keratinocytes (Tammi et al. 1998).
During embryonic development, CD44 is often expressed in sites of active morphogenesis, such as the apical ectodermal ridge (Yu et al. 1996), developing tooth, ear, and nose (Yu and Toole 1997), and skin (Underhill 1993). The CD44v3 heparan sulfate proteoglycan in the apical ectodermal ridge correlates with FGF level (Sherman et al. 1998) rather than hyaluronan (Yu et al. 1996) and contributes to the growth of the mesenchymal cells in limb bud elongation (Sherman et al. 1998). In early murine skin, the growth phase of the hair follicle was associated with elevated levels of CD44 in the dermal papilla (Underhill 1993). CD44-positive cells were present in the epidermis and dermis after the twelfth day and eleventh week of gestation in the rat (Weber et al. 1996b) and human (Terpe et al. 1994), respectively.
The present work was undertaken to establish a more systematic description of the temporal and spatial changes of CD44 expression in human fetal skin We also wanted to reveal the time of appearance of the heparan sulfate proteoglycan CD44v3 because of its high abundance in adult epidermis (Tuhkanen et al. 1997) and its suggested growth-promoting function during development. It is shown that CD44 appears relatively late compared to hyaluronan (Ågren et al. 1997), both in epidermis and in dermis. The appearance of CD44 and its v3 variant parallels the start of stratification in epidermis. In dermis, CD44 emerges with the formation of the fibrillar extracellular matrix.
Materials and Methods
Fetal Material
Human fetal skin was obtained after legally approved abortions in Kuopio University Hospital. The use of fetal skin for these studies was approved by the Ethics Committee of the hospital, and the authors were in no way involved in decisions to terminate the pregnancies. Fifteen fetuses at estimated gestational ages (EGA) of 6–20 weeks were included in the study. Immediately after abortion, samples of skin were removed from the back of the fetus at the level of the iliac crest and from the head at the site of the anterior fontanelle. The skin was immersed overnight in a fixative containing 2% paraformaldehyde, 2.5% glutaraldehyde, and 0.5% cetylpyridinium chloride in 0.1 M sodium phosphate buffer, pH 7.4 (PB) (Ripellino et al. 1985) or in neutral buffered formalin. The blocks were dehydrated and embedded in paraffin. Sections 3 μm thick were cut and deparaffinized according to normal protocols.
Immunostaining of CD44
The slides were incubated with the blocking buffer (1% fat-free milk powder in PB) for 30 min to block nonspecific binding. The generation of the anti-human CD44 antibody Hermes3 has been previously described (Jalkanen et al. 1987). This MAb recognizes an epitope in the common backbone of all CD44 forms, corresponding to codons 152–235 (Goldstein et al. 1989). Hermes3 MAb (kindly provided by Dr. Sirpa Jalkanen; Turku, Finland) was diluted with the blocking buffer (1:200 of the culture supernatant) and incubated with the sections overnight at 4C. After washes in PB, the sections were sequentially incubated for 1 hr at room temperature (RT) with biotinylated anti-mouse antibody (Vector Laboratories, Irvine, CA; 1:100), and avidin-biotin-peroxidase (ABC, Vector Laboratories; 1:200). After washes with buffer, the slides were incubated in 0.05% DAB (3,3′-diaminobenzidine; Sigma, St Louis, MO) and 0.03% hydrogen peroxide in PB at RT for 5 min. Finally, the sections were counterstained with Mayer's hematoxylin (Riedel-de Haën; Seelze, Germany) for 1.5 min, and mounted in DePeX (BDH Laboratory Supplies; Poole, UK). The specificity of the staining was controlled by incubating sections without the primary antibody, or the primary antibody was replaced with nonimmune IgG (IgG2a; Sigma).
The monoclonal antibody against the CD44 variant exon 3 was generated by immunizing mice with a recombinant fusion protein containing exons v3-v10 (Fox et al. 1994). One of the clones, 3G5, recognizes CD44 forms containing exon v3 (Fox et al. 1994). For the CD44v3 immunostaining, the sections were subjected to microwave treatment before incubation with the primary antibody (anti-human CD44v3, clone 3G5; R&D Systems, Oxon, UK, dilution 1:400). The dilutions of the biotinylated anti-mouse antibody and ABC (ABC Elite kit; Vector) were 1:200 and 1:25, respectively.
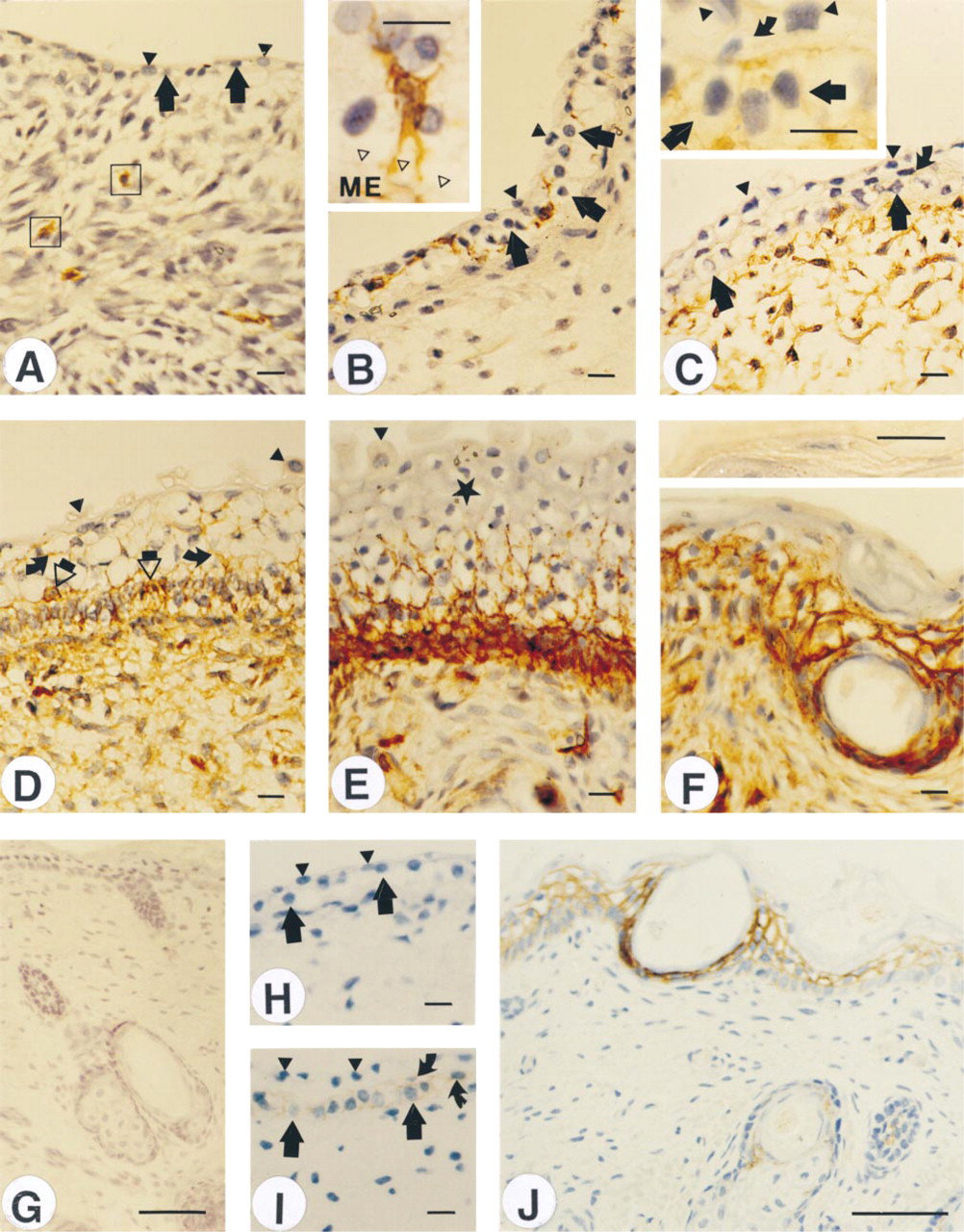
Expression of CD44 is associated with maturation of mesenchymal tissue and stratification of the epidermis. Skin from the lower back (A,D) or from the scalp (B,C,E,F,H-J) of fetuses with estimated gestational ages of 8 (A), 9 (B,H), 11 (C,I), 12 (D), 17 (E), and 20 (F,G,J) weeks was fixed and stained for CD44 using Hermes3 antibody. Although most cells in the embryonic skin are CD44-negative, a few cells show strong positive staining in both tissue compartments (A,B). The inset in B shows a high-power view of such a cell inside the epithelium. At the transition from the embryonic to the fetal period (C), most dermal cells became CD44-positive, and epidermis shows at first just faint staining in the basal cell layer (inset) but later exhibits a strong signal (D-F). The terminally differentiated keratinocytes, including the upper intermediate cells at 17 weeks (E) and granular cells and corneocytes at 20 weeks (F, inset), are unstained. H-J are stained using anti-variant 3 MAb. The epithelium and the mesenchyme were totally negative in H, while after the eleventh week a signal was detected in basal and intermediate layers (I). When keratinization started, the signal became stronger in the epidermis and hair follicle (J). Isotypically matched nonimmune IgG produced no signal (G). Closed triangles, periderm; thick arrows, germinative layer; curved arrows (C,D), intermediate cells; asterisk (E), upper intermediate cells; ME (inset in B), mesenchyme; open triangles (inset in B), basal lamina; open arrows (D), dendritic cells; open squares (A), CD44-positive stellate cells. Bars: A-F,I and insets = 10 μm; G,J = 50 μm.
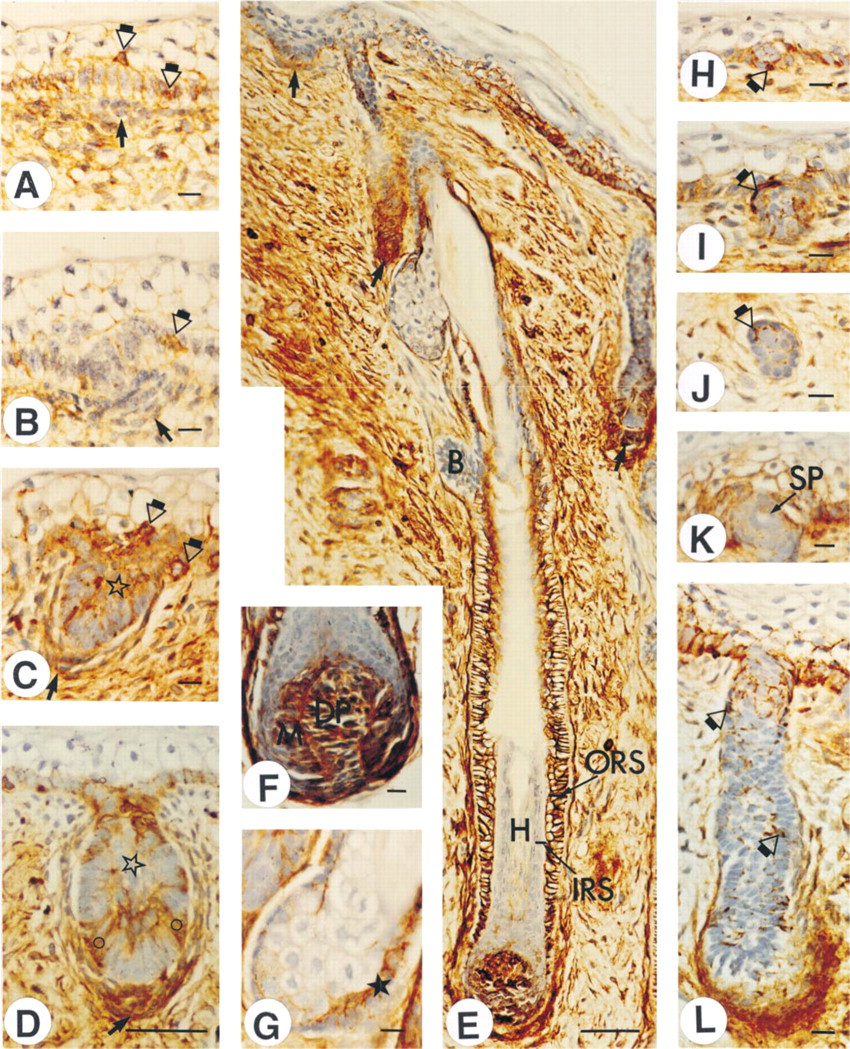
Initiation of hair follicle and sweat gland development is associated with a transient decrease in epithelial CD44. Fetal skin specimens at the estimated gestational ages of 12 (A,B), 17 (C,D,H-L), and 20 (E-G) weeks were obtained from the lower back (A-D,H,I) or scalp (E-G,J-L). CD44 expression is low in the pre-germ hair follicle anlagen (A). During the development of the follicle (B-D), CD44 expression increases in the follicle epithelium and in the dermal condensation and becomes compartmentalized in the fully developed follicle (E,F). A sebaceous gland (G) shows a signal in the progenitor cell layer. Expression in the hair bulge is low (E). The sweat gland anlagen (H-L) shows a relatively low level of CD44 staining. Small arrows in A-D, dermal condensation in primary follicles; small arrows in E, dermal condensation in secondary follicles; open circle in D, intensely stained distal part of the hair peg; open asterisk in D, inner core cells in hair peg; ORS (E) outer root sheath; IRS (E), inner root sheath; H (E), hair; B (E), bulge; M (F), hair matrix; DP (F), dermal papilla; closed asterisk (G), progenitor cells in sebaceous gland; open arrows (A-C,H-J,L), intensely stained cells; SP (K), intraepidermal sweat pore. Bars: A-C,F,L = 10 μm; D,E = 50 μm.
Results
Dermal Mesenchyme
Most of the primitive mesenchymal cells in the embryonic skin were negative for CD44. Only occasional strongly CD44-positive stellate or elongated cells were present when the fetus was about 6 weeks old, first in the lower part of the dermal mesenchyme (Figure 1A). Three weeks later, cells with similar morphology and CD44 signal emerged also in the upper dermis and close to the epidermal-dermal junction (not shown).
In the eleventh week, most of the dermal cells had acquired at least a low level of CD44 expression (Figure 1C). However, at 12 weeks (Figure 1D), 17 weeks (Figures 1E, 2C, and 2D), and 20 weeks (Figure 2E) the signal was also associated with the fibrillar matrix. This matrix-associated CD44 was less intense in the deep connective tissue below the dermis (Figure 2E) and did not contain variant exon 3 (Figure 1J).
Nonkeratinocytes
The germinative epithelial cells and the peridermal cells in the embryonic skin did not express CD44 (Figures 1A, and 1B). However, as early as 6 weeks, occasional strongly Hermes3-positive but v3-negative cells with dendritic morphology were seen above the germinative cells (not shown). Two weeks later, cells with similar morphology resided more frequently between the periderm and the germinative layer (Figure 1B) and subsequently also within the basal layer (Figures 1D and 2A-2C).
Epidermal Keratinocytes
The formation of the first intermediate cell layer at 11–12 weeks coincided with the first signs of CD44 expression in the germinative (basal) layer (Figure 1C, inset), whereas the newly formed intermediate cell layer and the peridermal layer remained CD44-negative (Figure 1C). With increasing epidermal thickness, the CD44 signal grew stronger in the basal layer and the intermediate cells became CD44-positive (Figures 1D and 2A-2C). From the beginning, the stratifying epidermal cells were also positive for variant exon 3 (Figure 1I).
Maturation of epidermis was associated with the loss of CD44 expression from the upper intermediate cells at the age of 17 weeks (Figure 1E). With complete keratinization of epidermis (EGA 20 weeks), the granular cells (Figure 1F), and the corneocytes were CD44-negative (Figure 1F, inset). At this stage, the lower cell layers (basal and spinous cells) were intensely CD44-positive (Figure 1F).
Hair Anlagen and Dermal Condensation
The tightly packed cylindrical basal epithelial cells in the initial hair anlagen (pre-germ) stained less intensely for CD44 than the surrounding interfollicular basal cells or the intermediate cells (Figure 2A). This was even more evident in a late pre-germ follicle (Figure 2B). When the hair germ was formed, its outer cells showed less intense staining for CD44 than the inner cells (Figure 2C). Among the latter, occasional strongly positive cells with dendritic morphology were present (Figure 2C). The mesenchymal cell aggregation below the epithelial germ, which at first displayed faint CD44 expression (Figure 2B and 2E), became strongly CD44-positive during elongation of the follicle (Figures 2C-2E).
The outer epithelial cells on the deep part of the hair peg were more strongly CD44-positive than those in the more superficial end and at the base of the peg (Figure 2D and 2E). Formation of bulbous pegs was associated with the compartmentalization of CD44, with strong expression in the ORS (outer root sheath) and loss of CD44 from the terminally differentiated part of IRS (inner root sheath) and the hair (Figure 2E and 2F). In the matrix of the hair and IRS, the progenitor cells remained CD44-positive (Figure 2F).
Interestingly, the putative stem cells in the hair bulge exhibited staining from faint to negative, clearly different from the rest of the ORS (Figure 2E).
The mesenchymal cells in the dermal papillae were strongly CD44-positive (Figure 2F). In addition, the cells forming the connective tissue sheath around the follicle showed a strong signal (Figure 2E).
Sebaceous Cells
The undifferentiated progenitor cells in the sebaceous gland primordium were CD44-positive, whereas the maturing sebocytes were only occasionally positive and fully mature cells expressed no CD44 (Figure 2E and 2G).
Sweat Gland
In the sweat gland bud, the outer keratinocytes were CD44-positive, whereas the inner cells were almost negative (Figure 2H). A strongly positive cell with dendritic morphology was often present in the bud (Figure 2H and 2I). The more elongated sweat gland ducts exhibited a very low level of CD44 expression, most of the outer and inner cells being negative (Figure 2J and 2L). Occasional intensely stained cells and cell processes were seen among the inner cells (Figure 2J and 2L). Cells surrounding the intraepidermal sweat pore were CD44-negative, in contrast to the surrounding basal and intermediate keratinocytes (Figure 2K).
CD44v3 Expression
Early fetal skin showed no signal with the monoclonal antibody specific for v3 (Figure 1H). When epidermis started to stratify, a faint v3 signal appeared in basal and suprabasal layers (Figure 1I), similar to that of Hermes3 (Figure 1C), and increased in intensity with epidermal maturation and particularly with the onset of true cornification (Figure 1J). The signals for v3 in the epithelial cells of developing hair follicles were also virtually superimposable with those obtained with Hermes3 (not shown). In a contrast, all mesenchymal cells and cells with dendritic morphology remained negative, indicating restriction of this variant to the keratinocyte lineage in human skin (Figures 1H-1J). The v3 signal was also absent from the dermal extracellular matrix, suggesting that this CD44 did not originate from epidermis.
Discussion
Epidermis
The present study showed that CD44, with its variant exon v3, first appears in the basal cells of epidermis during the transition to a stratified epithelium. At the same time, the basal cells adopt columnar morphology, contain less glycogen, and express new keratin types (reviewed by Dale et al. 1985; Holbrook 1991). A week later, the intermediate cells begin to express CD44, simultaneously with an altered, more mature suprabasal keratin pattern (Dale et al. 1985; Lane et al. 1985). These findings indicate that CD44, with the v3 exon, belongs to a set of proteins that mark the differentiation lineage of stratifying keratinocytes.
The importance of CD44 is largely due to its ability to bind and immobilize hyaluronan in the epidermis (Kaya et al. 1997) and to organize hyaluronan in the keratinocyte pericellular matrix (Tammi et al. 1998). The space provided by hyaluronan, in turn, allows the exchange of metabolites and waste products between the multilayered epithelium and the circulation. Hyaluronan may also offer a protective coating against various noxious agents. Perhaps also important in the present context, the increased amount of keratinocyte surface hyaluronan by CD44 is believed to facilitate the upward movement and shape change that occur in differentiating keratinocytes. This continuous cell reorganization is probably fundamental for stratification and wound healing (Oksala et al. 1995; Kaya et al. 1997).
Interestingly, peridermal cells and early epithelial cells in human epidermis display hyaluronan (Ågren et al. 1997) but not CD44. This suggests that, in a primitive epidermis lacking CD44, hyaluronan may be bound directly to the hyaluronan synthase (Weigel et al. 1997) or to another as yet unknown receptor (s). Thus far, none of the other known hyaluronan receptors has been described in early fetal epidermis (reviewed by Tammi et al. 1994). In a contrast to human skin, murine peridermal cells express CD44 (Underhill 1993).
Epidermal Nonkeratinocytes
The strongly CD44-positive but v3-negative cells with dendritic morphology in the early epithelia probably represent Langerhans cells, melanocytes, or Merkel cells (Holbrook 1991; Kim and Holbrook 1995). All these cell types are CD44-positive in normal adult skin (Penneys and Shapiro 1994; Tammi et al. 1994; Osada et al. 1995; Danen et al. 1996) and do not express v3 (Herbold et al. 1996; Weiss et al. 1997). The present data indicate that the nonkeratinocytes display CD44 earlier than the epithelial cells. The expression of CD44 probably facilitates their migration into the epidermis through the hyaluronan-rich embryonic matrix (Ågren et al. 1997). Cells may use CD44 as an endocytosis receptor to invade into a viscous gel formed by hyaluronan, or migrate on hyaluronan using CD44 as an adhesion receptor. In addition, CD44 may mediate signals to enhance cell motility (Ohta et al. 1997).
Dermis
CD44 appeared relatively late in the mesenchyme, apparently being expressed with the gene set associated with the transition of dermis from a cellular to a fibrous tissue (Holbrook 1991). Interestingly, this newly formed fibrous tissue soon gained strong matrix staining for CD44. Matrix-associated CD44 exists in malignant stroma (Furuta et al. 1998) and in adult human skin (unpublished observations). CD44 may be shed from the cell surface by proteolysis (Bazil and Strominger 1994) or may represent a variant form completely lacking the membrane-spanning and intracellular domains, as described in developing muscle and cartilage (Yu and Toole 1996). The matrix-associated CD44 in dermis may bind to its other matrix ligands, fibronectin and collagens (Jalkanen and Jalkanen 1992; Yu and Toole 1996), or to osteopontin (Weber et al. 1996a), which is associated with elastin fibers (Baccarani et al. 1995), as also suggested by the fibrillar appearance of the signal. Although the significance of the matrix-bound CD44 remains unknown at present, it may serve as an antagonist of the cell surface form (Yu and Toole 1996). Soluble CD44 exists in normal serum, and its concentration increases in inflammation and various malignancies (Ristamäki et al. 1996; Kittl et al. 1997).
The cells in the developing dermal papilla showed strong CD44 expression, as reported earlier in murine skin (Underhill 1993; Yu and Toole 1997). The CD44 in the dermal papillar cells was suggested to act as an endocytosis receptor for hyaluronan, to make room for the growing follicle (Underhill 1993), but it may also enhance migration of the cells (Yu and Stamenkovic 1999).
Skin Appendages
The basal cells at the initiation site of the hair follicle (pre-germ) expressed little CD44 and hyaluronan (Ågren et al. 1997). The removal of CD44, with its highly hydrated ligand, may be necessary for the tight packing of this cell aggregate. During elongation of the hair follicle CD44 expression was reacquired by the follicular cells and was most intense close to the lower end of the follicle, suggesting its involvement in penetration into the mesenchyme. It was recently reported that CD44 with associated hyaluronan binds and activates matrix metalloproteinase 9, which may aid in the invasion through the matrix (Yu and Stamenkovic 1999). In mature fetal hair follicles, CD44 was strictly compartmentalized, as in adult skin (Wang et al. 1992), and was co-distributed with hyaluronan (Ågren et al. 1997).
The fetal hair bulge differed from the rest of the ORS by its lower CD44 signal. The bulge cells have a morphology characteristic of undifferentiated keratinocytes (Akiyama et al. 1995), and it is assumed that the stem cells for the follicle reside in the bulge (Costarelis et al. 1990). This supports the idea that undifferentiated cells in the keratinocyte lineage show a lower level of CD44 expression than their more differentiated counterparts. Interestingly, the low CD44 expression of the bulge cells was in a contrast to their strong hyaluronan signal (Ågren et al. 1997).
The developing sweat gland anlagen exhibited only little CD44 and hyaluronan (Ågren et al. 1997). The high level of CD44 expression observed in the clear cells of adult sweat gland acini (Wang et al. 1992) therefore develops during maturation of the gland later in fetal life (Hashimoto et al. 1966).
Footnotes
Acknowledgements
Supported by a grant from the Hulda Tossavainen Fund of the North Savo Cultural Foundation to A-LT.
We thank Ms Eija Rahunen, Ms Arja Venäläinen, Mr Kari Kotikumpu, Mrs Aija Parkkinen, Ms Riikka Eskelinen, and Ms Tiina Räsänen for skillful technical assistance.