
Abstract
We determined the distributions of the small proteoglycans biglycan and decorin and the glycoprotein matrilin-1 (cartilage matrix protein) during development and aging of articular cartilage in the rabbit knee joint. Before cavitation, the matrices of the interzone and the adjacent epiphyseal cartilage do not contain biglycan or decorin, but some chondrocytes express their mRNAs. Matrilin-1 is found only in the deeper epiphyseal cartilage. After cavitation, biglycan and decorin are detected in the presumptive articular cartilage, but there is no matrilin-1. All are present in the underlying epiphyseal cartilage. In the neonate, the epiphyseal cartilage is ossified and the articular cartilage becomes a discrete layer. Biglycan and decorin accumulate in the articular cartilage, but matrilin-1 remains confined to the residual epiphyseal cartilage. In the adult, the distributions of biglycan and decorin are highly variable. Decorin tends to be confined to the central region; matrilin-1 is absent. The findings indicate that the articular and epiphyseal cartilages are different from the earliest developmental stages. The epiphyseal cartilage can be identified by its possession of matrilin-1. Epiphyseal cartilage is removed during development to leave the articular cartilage. The relationships between the distributions of decorin and matrilin-1 and the fibrillar collagens are discussed.
Keywords
Biglycan and decorin have very similar core proteins which are substituted with two and one glycosaminoglycan (GAG) chains, respectively (Fisher et al. 1989; Oldberg et al. 1989). In articular cartilage, the GAG chains are mostly chondroitin, rather than dermatan, sulfate (Cheng et al. 1997).
Decorin may have a role in collagen fibrillogenesis. It binds to the fibrils in specific regions (Pringle and Dodd 1990; Hedbom and Heinegård 1993; Svennson et al. 1995). It also binds to Type VI collagen, which may act as an intermediary in the binding of decorin to Types I and II collagen (Bidanset et al. 1992). The amount of decorin present affects fibril development in chick tendon (Birk et al. 1995). It inhibits fibrillogenesis of Type I collagen in vitro and in vivo (Daniel-son et al. 1997; Sini et al. 1997).
The function of biglycan is less well understood. It may be involved in the promotion of matrix mineralization (Geerkens et al. 1995; Boskey et al. 1997). The core protein can self-associate and exists primarily as a hexamer (Liu et al. 1994).
Biglycan and decorin in human, bovine, and marsupial adult articular cartilages are mainly confined to the upper region (Heinegård et al. 1986; Poole et al. 1986; Bianco et al. 1990; Miosge et al. 1994; Archer et al. 1996). Biochemical studies indicate an increase in a non-PG form of biglycan with increasing age in human articular cartilage (Roughley et al. 1993). Expression of biglycan mRNA decreases, whereas expression of decorin mRNA increases, in human chondrocytes with increasing age of the donor (Roughley et al. 1994).
Matrilin-1 is a 148-kD glycoprotein with three disulfide-bonded monomers (Paulsson and Heinegård 1981). It binds to Type II collagen and decreases the rate of fibril formation in vitro so that the fibrils remain of small diameter (Chandrasekhar et al. 1986; Winterbottom et al. 1992). It can self-associate and form filaments independently or with Type II collagen (Aszodi et al. 1994; Chen et al. 1995). It also binds to aggrecan (Paulsson and Heinegård 1982; Hauser et al. 1996).
Matrilin-1 is not found in normal human and chicken articular cartilage at any stage in development (Paulsson and Heinegård 1981,1982; Saxne and Heinegård 1989; Stirpe and Goetnick 1989; Mundlos and Zabel 1994), but Okimura et al. (1997) found matrilin-1 in some articular chondrocytes from human adults.
In both osteoarthritic and rheumatoid cartilage there is increased degradation of biglycan and decorin compared to age-matched control tissue (Beesley et al. 1992; Cs-Szabo et al. 1995), but their synthesis is also increased in osteoarthritis (Cs-Szabo et al. 1997; Okimura et al. 1997). Matrilin-1 appears in osteoarthritic cartilage (Saxne and Heinegård 1989).
In the rabbit knee joint, precise changes with age in the fibrillar collagens indicate that the articular cartilage is unique and distinguished from the underlying epiphyseal cartilage from the earliest stages of joint development (Bland and Ashhurst 1996). To investigate the concomitant changes in the small PGs, the distributions of biglycan and decorin and the cells expressing their mRNAS have been determined. Matrilin-1 is not present in normal healthy articular cartilage. It is a constituent of the underlying epiphyseal cartilage in the rabbit (unpublished observation). Because matrilin-1 appears in diseased cartilage, its distribution was determined to establish its relationship with normally developing articular cartilage.
The aims of this study were, therefore, first to investigate the changes in the distribution of the small PGs during development and aging, second, to determine whether the results provide any further evidence for the uniqueness of articular cartilage, and finally to provide data that will enable a better understanding of the changes that occur with the onset of osteoarthritis.
Materials and Methods
All reagents and labeled antibodies were obtained from Sigma (Poole, UK) and all restriction enzymes and polymerases were obtained from Roche (Lewes, UK) unless otherwise stated.
Preparation of Tissue
Hindlimbs distal to the mid-femoral region were removed from New Zealand White rabbit fetuses aged 17, 20, and 25 days. Knee joints were dissected from newborn and 1-week-old rabbits. The tibial plateau and underlying bone were taken from 3-, 6-, and 12–14-week, 8-month, and 2-year-old rabbits. At least two rabbits were used at each stage. The tissues were fixed in 4% paraformaldehyde in 0.05 M Tris-Hcl buffer, pH 7.3, for 18 hr, then washed extensively in buffer. With the exception of 17-day joints, the tissue was decalcified in 14.3% EDTA, pH 7.0, until radiographically free from calcium. After washing, the tissue was dehydrated in graded ethanols, cleared in methyl salicylate, and embedded in paraffin. Sagittal longitudinal sections were cut at 7 μm. For histological observations, sections were stained with hematoxylin and eosin.
Immunohistochemistry
Antibodies to Biglycan and Decorin. Chicken polyclonal antibodies to purified bovine biglycan and purified bovine decorin (a kind gift from Prof. D. Heinegård) were used. The total IgG was purified as described by Heinegård et al. (1985).
Sections were dewaxed and rehydrated before the following standard pretreatments: (a) 0.1% trypsin in 0.05 M Trissaline, pH 7.8, with 0.1% CaCl2 at 37C for 1 hr (to reduce crosslinking and aid tissue permeability); (b) 2% hyaluronidase in PBS, pH 7.3, at 37C for 1 hr (to remove GAG chains and unmask antigenic sites); and (c) 2% L-lysine in PBS for 15 min (to reduce charged sites). The sections were washed in PBS between each treatment. To block nonspecific binding, the sections were incubated in heat-inactivated normal rabbit serum (Harlem Sera Labs; Loughborough, UK) with the addition of 4% bovine serum albumin (BSA) and 0.3% Triton X-100 for 30 min. This was drained from the slide and the primary antibody was added at a dilution of 1:1000 in 1% BSA. The sections were incubated overnight at 4C. After washing, the sections were incubated in alkaline phosphatase-labeled rabbit anti-chicken IgG antibodies for 90 min. The alkaline phosphatase label was localized using the following substrate: 5 μg naphthol AS-BI phosphate was dissolved in 1 drop dimethylformamide and added to 5 μg Fast Red TR in 10 ml veronal acetate buffer, pH 9.2. Levamisole (1 μg/ml) was added to inhibit endogenous alkaline phosphatase activity. The sections were incubated in the substrate for 20 min before being washed and mounted in glycerin jelly.
For negative controls, the primary antibody was replaced with normal chicken serum; at no time was there any binding (see Figure 6). For positive controls, using the same protocol, biglycan and decorin antibody binding was localized in skin and ligaments, which are known to contain biglycan and decorin (not shown).
Antibodies to Matrilin-1. Rabbit polyclonal antibodies to purified bovine tracheal matrilin-1 (a kind gift from Prof. M. Paulsson) were used. They were prepared as described by Hauser and Paulsson (1994). Because the antibody to matrilin-1 was raised in rabbit, precautions were taken to prevent the secondary antibody (alkaline phosphatase-labeled goat anti-rabbit IgG) from binding to the tissue sections. The method of Hierck et al. (1994) was used, with some modifications.
Anti-matrilin-1 antibody (4 μl) and goat anti-rabbit IgG (1 μl) were mixed together in 400 μl PBS and stirred overnight at 4C. One μl of heat-inactivated normal rabbit serum was added to remove the excess anti-rabbit IgG and stirred at 4C for at least 2 hr.
After the same pretreatments used for biglycan and decorin antibodies, the complexes were placed on the sections and incubated at room temperature for 4 hr. The sections were then washed with PBS and the labeled antibodies were localized as above. The sections were mounted in glycerin jelly.
For negative controls, the primary antibody was replaced with normal rabbit serum. At no time was there any binding in the negative controls. Matrilin-1 is found only in cartilage. The binding of the complex by all other appropiate cartilaginous regions, e.g., growth plate and axial skeleton, served as a positive control. At no time was there any binding in noncartilaginous tissues.
In Situ Hybridization
Generation of Riboprobes. cDNA probes to human biglycan and decorin were a gift from Dr. L. Fisher (NIH; Bethesda, MD). These probes recognized the appropiate rabbit mRNAs on Northern blots (Demoor-Fossard et al. 1998). Antisense and sense probes were generated by linearizing with Kpn I and T3 polymerase and Xba I and T7 polymerase, respectively, for biglycan and Bam HI and T7 polymerase and Kpn I and T3 polymerase, respectively, for decorin. Both probes were labeled with digoxigenin using a kit (Roche). The riboprobes are 1600 bp (decorin) and 1685 bp (biglycan) long. The transcripts were hydrolyzed at 60C for 40 min in 100 nM bicarbonate buffer to yield fragments of ∼200 bp and were stored at −20C until use.
Procedure. The sections were dewaxed and rehydrated. The following pretreatments were performed: (a) 0.2 N HCl for 20 min; (b) 6% H2O2 for 30 min; (c) proteinase K (20 μg/ml) for 10 min at 37C; (d) 4% paraformaldehyde in PBS for 20 min at 4C; (e) 0.1 M glycine twice for 10 min; and (f) 0.25% acetic anhydride in triethanolamine (pH 8) for 10 min. Between each treatment, the sections were washed in PBS. They were then dehydrated through graded ethanols and allowed to dry. Fifteen μl of hybridization solution was applied to each section. The hybridization solution (low stringency) contained 50% formamide, 10 mM Tris-HCl (pH 7.4), 1 mM EDTA, 1 × Denhardt's, 0.5% SDS, 600 mM NaCl, 10% dextran sulfate, 0.5 μg/ml yeast tRNA (Roche), and labeled antisense or sense probe at a final concentration of 1.7 ng/ml. Hybridization was for 18 hr at 45C for decorin mRNA and 50C for biglycan mRNA. After hybridization, the coverslips were removed in 2 × SSC before rinsing in Tris-EDTA buffer. The sections were then treated with 20 μg/ml RNase in Tris-EDTA-NaCl buffer, pH 8.0, for 20 min at 37C. The sections were washed twice in 2 × SSC for 15 min, then 1 × SSC for 10 min at 55C; a final wash in 1 × SC was at room temperature. The digoxigenin label was detected using the Roche kit, except that 0.3% Triton X-100 was added to the antibody solution. This blocks the nonspecific binding by cartilaginous matrices that arises after treatment with proteolytic enzymes (Bland et al. 1991). The sections were mounted in glycerin jelly.
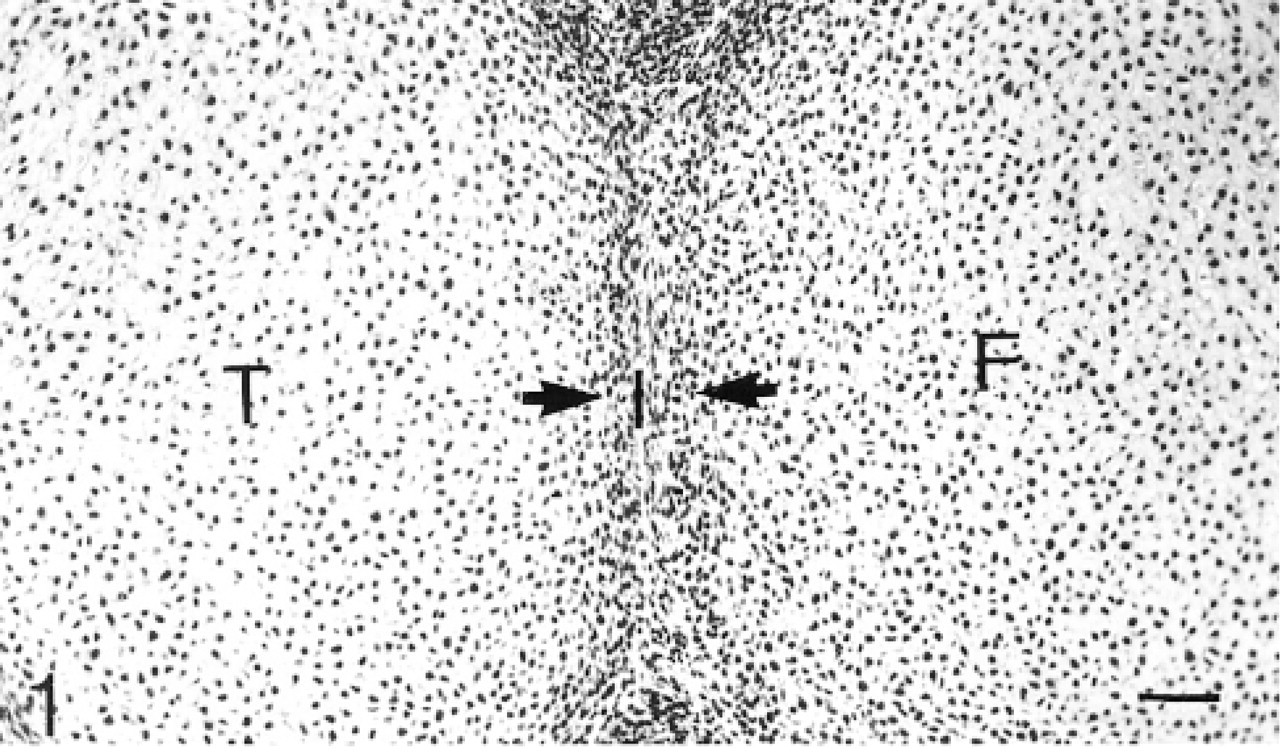
Cavitation has not yet occurred. The chondrogenous layers of the interzone are clearly seen. Hematoxylin and eosin. Bar = 50 μm.
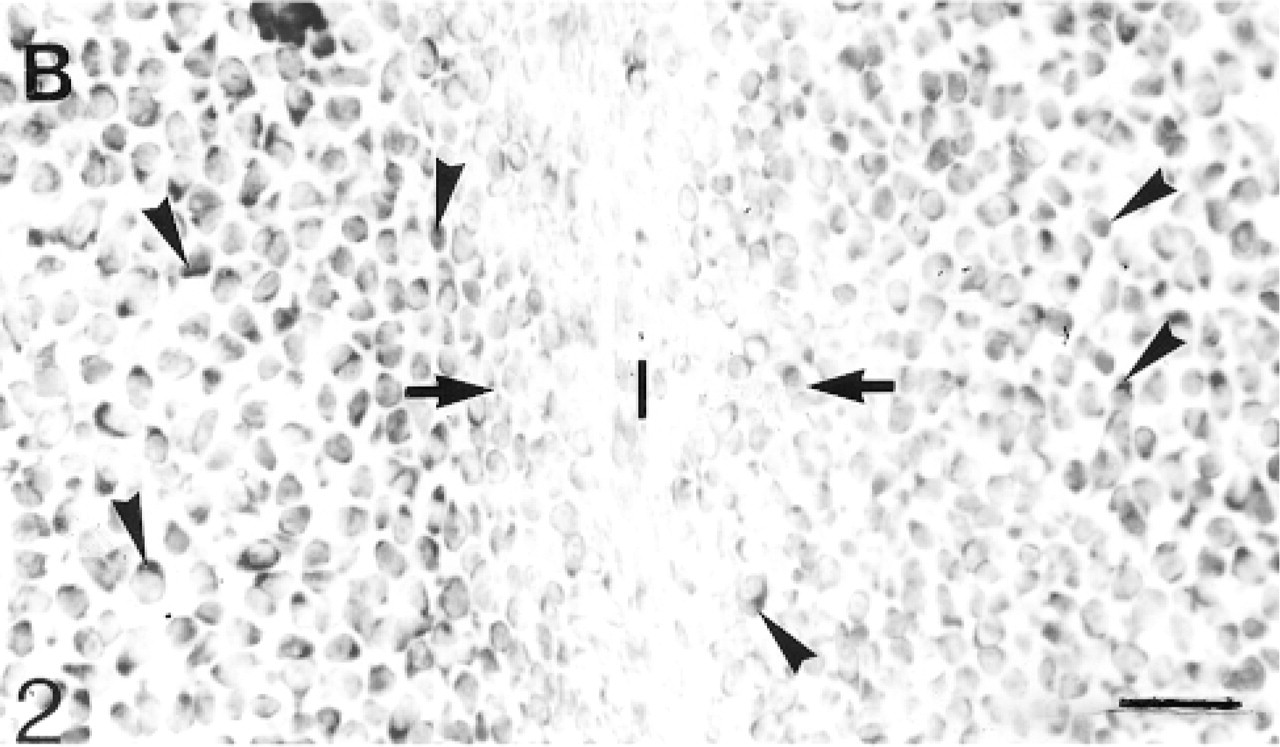
In situ hybridization for biglycan (B) mRNA: Many cells (arrowheads) in the epiphyseal cartilage but very few in the inter-zone express the mRNA. Bar = 30 μm.
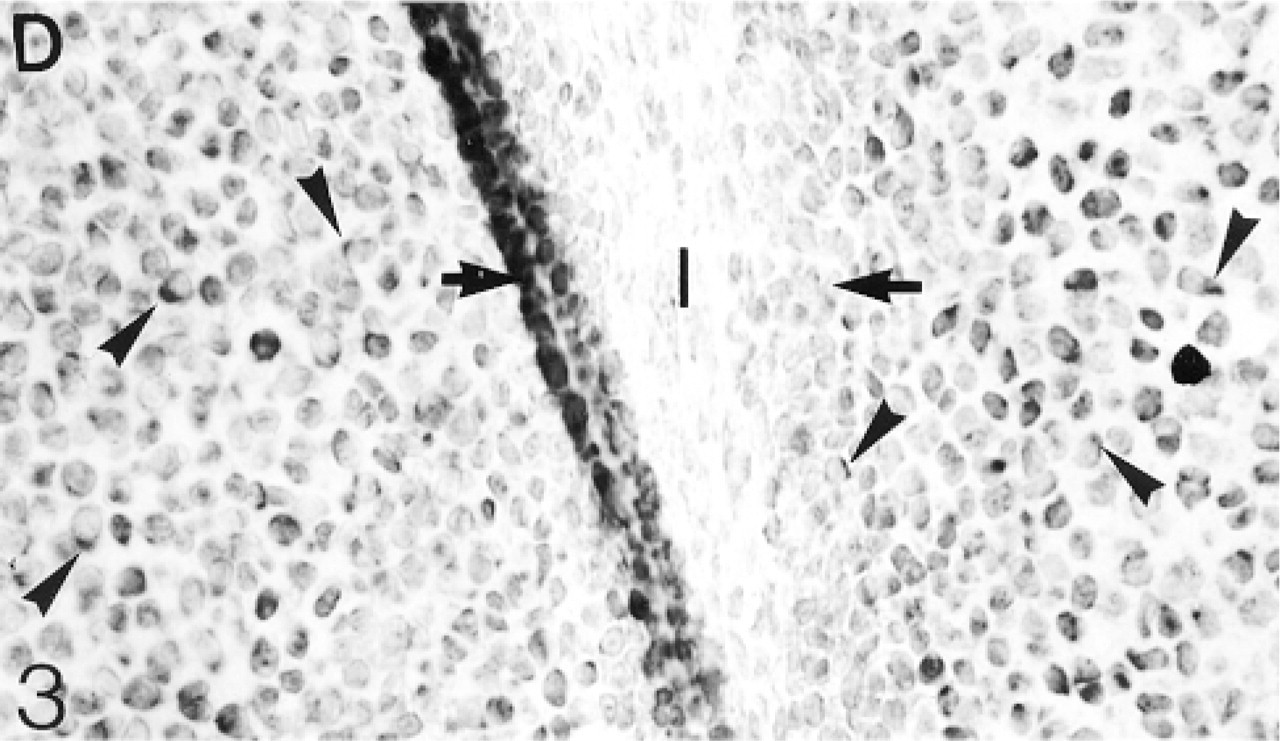
In situ hybridization for decorin (D) mRNA: Many cells (arrowheads) in the epiphyseal cartilage but very few in the inter-zone express this mRNA.

Distributions of biglycan, decorin, and matrilin-1 in fetal and neonatal kneejoints a

a+, antibodies bound; w+, antibodies bound weakly; -, antibodies not bound.
At no time was there any reaction with the sense, i.e., negative control, probes (see Figure 13). For positive controls, hybridization was performed on rabbit tissues, including ligaments, skin, and blood vessels. The cells in all these tissues are positive with both probes.
Results
Nomenclature
The term “epiphyseal cartilage” is used here to denote the cartilage in the epiphysis that will be removed by ossification to leave the articular and growth plate cartilages. In 17- and 20-day fetuses, the interzone comprises the two chondrogenous layers and the thin intermediate layer (Andersen 1961). The chondrogenous layers are distinguished from the underlying epiphyseal cartilage by their cell density (Figure 1) and the presence of Types I, III, and V collagen in the matrix (Bland and Ashhurst 1996). The characteristics of this layer of closely packed cells and its distinctive matrix persist as it develops into articular cartilage.
Fetal Knee Joints
Cavitation has not yet occurred in the 17- and 20-day fetal knee joints (Figure 1). Antibodies to biglycan are not bound by the matrix of the interzone. The antibodies are bound quite strongly by the hypertrophic cartilage in the diaphysis, but towards and into the epiphysis the antibody binding becomes progressively weaker until, in the region adjacent to the interzone, there is no detectable binding (not shown; Table 1). Many chondrocytes in the epiphyseal cartilage, but few in the interzone, express the mRNA for biglycan (Figure 2). The binding of decorin antibodies follows an identical pattern to that of biglycan (not shown; Table 1). Most chondrocytes in the epiphyseal cartilage, but only very few in the interzone, express decorin mRNA (Figure 3). Antibodies to matrilin-1 are not bound by the interzone or the epiphyseal cartilage immediately below, but they are bound strongly in the rest of the epiphyseal cartilage (Figure 4; Table 1).
By 25 days, cavitation has occurred. The chondrocytes in the future articular cartilage are more densely packed than those of the epiphyseal cartilage (Figure 5). The chondrocytes on the surface are flattened. Antibodies to biglycan bind weakly to the matrix of the developing articular cartilage but more strongly to the matrix of the epiphyseal cartilage below (Figure 7; Table 1). Many chondrocytes in both the articular and epiphyseal cartilages express the mRNA for biglycan (Figure 8). Decorin antibodies are bound very weakly in the developing articular cartilage, whereas the epiphyseal cartilage matrix binds the antibodies more strongly (Figure 9; Table 1). Many chondrocytes in both regions are expressing the mRNA for decorin (Figure 10). Antibodies to matrilin-1 are not bound by the matrix of the developing articular cartilage. The epiphyseal cartilage binds the antibodies very strongly (Figure 11; Table 1).
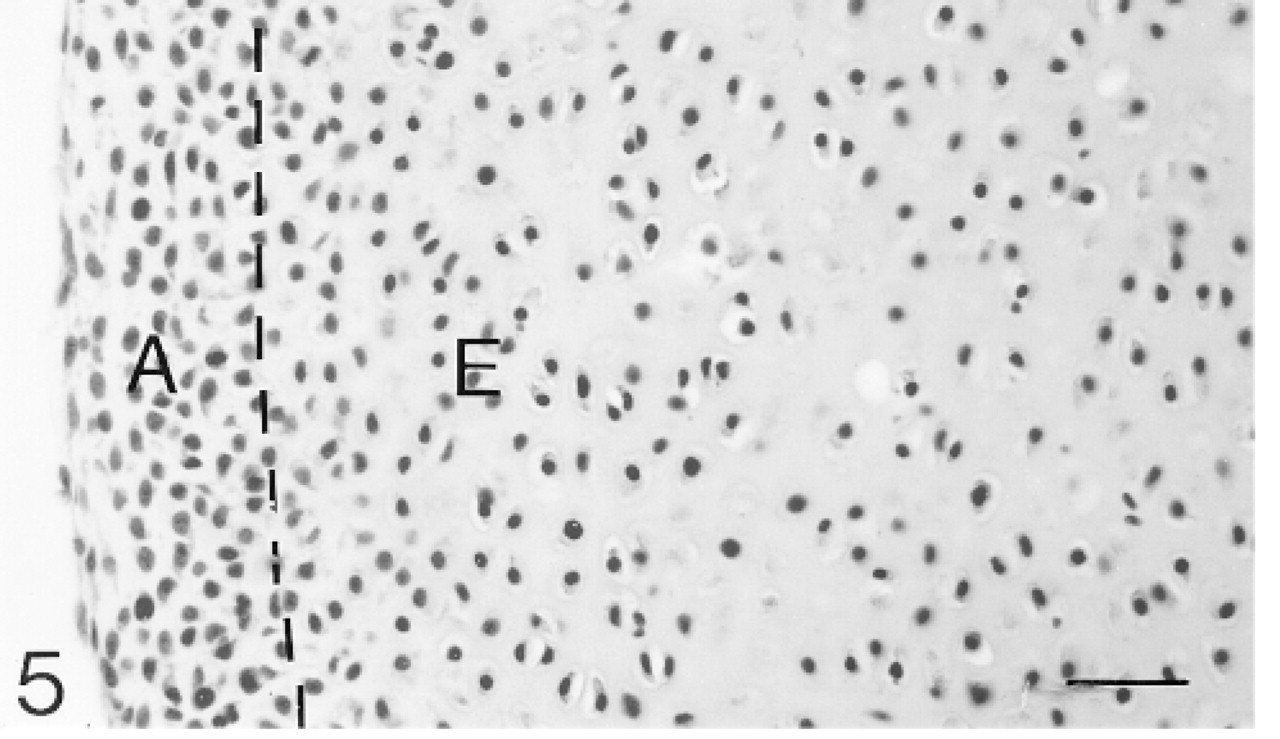
Cavitation is complete. A dashed line marks the boundary between the future articular (A) and epiphyseal (E) cartilages. Hematoxylin and eosin. Bar = 30 μm.

Negative control for immunohistochemistry; there is no evidence of nonspecific binding.

Binding of antibodies to biglycan (B). The epiphyseal cartilage binds the antibodies more strongly than the future articular cartilage

In situ hybridization for biglycan (B) mRNA.
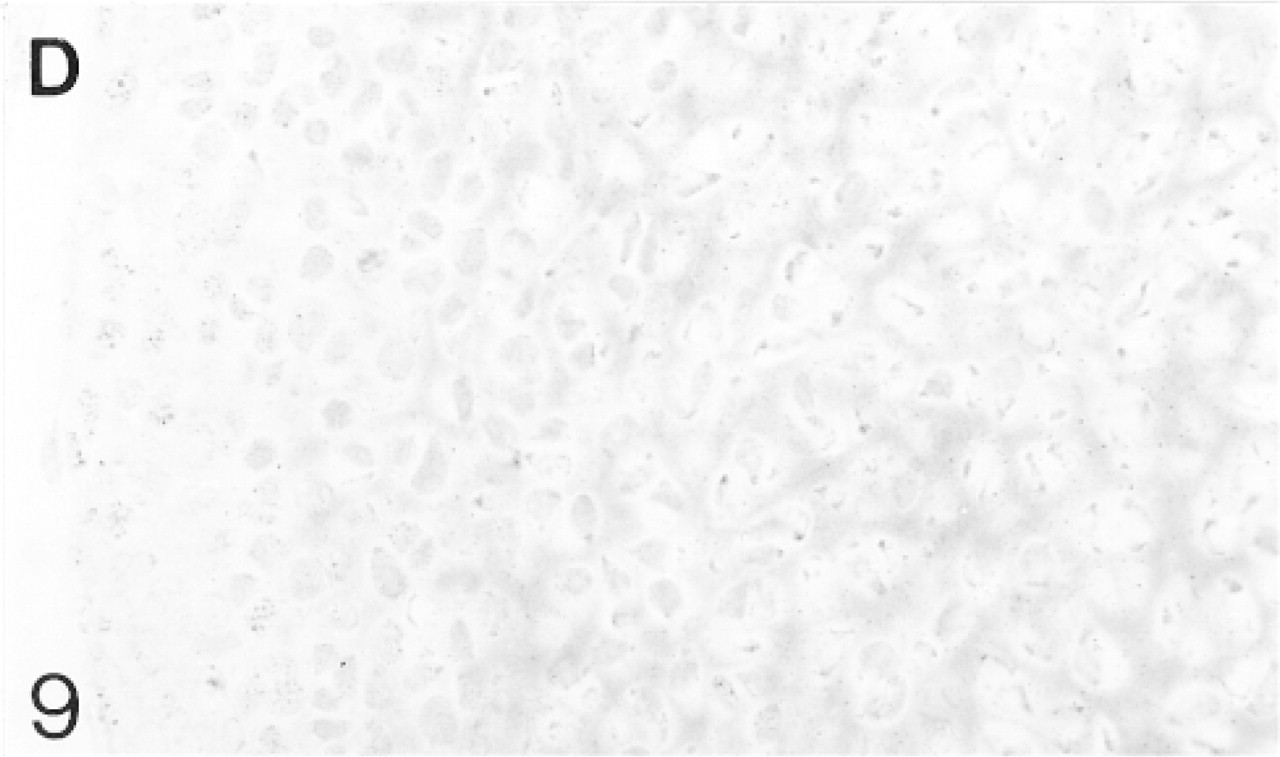
Binding of antibodies to decorin (D). The epiphyseal cartilage matrix binds the antibodies more strongly than the future articular cartilage.

In situ hybridization for decorin (D) mRNA.
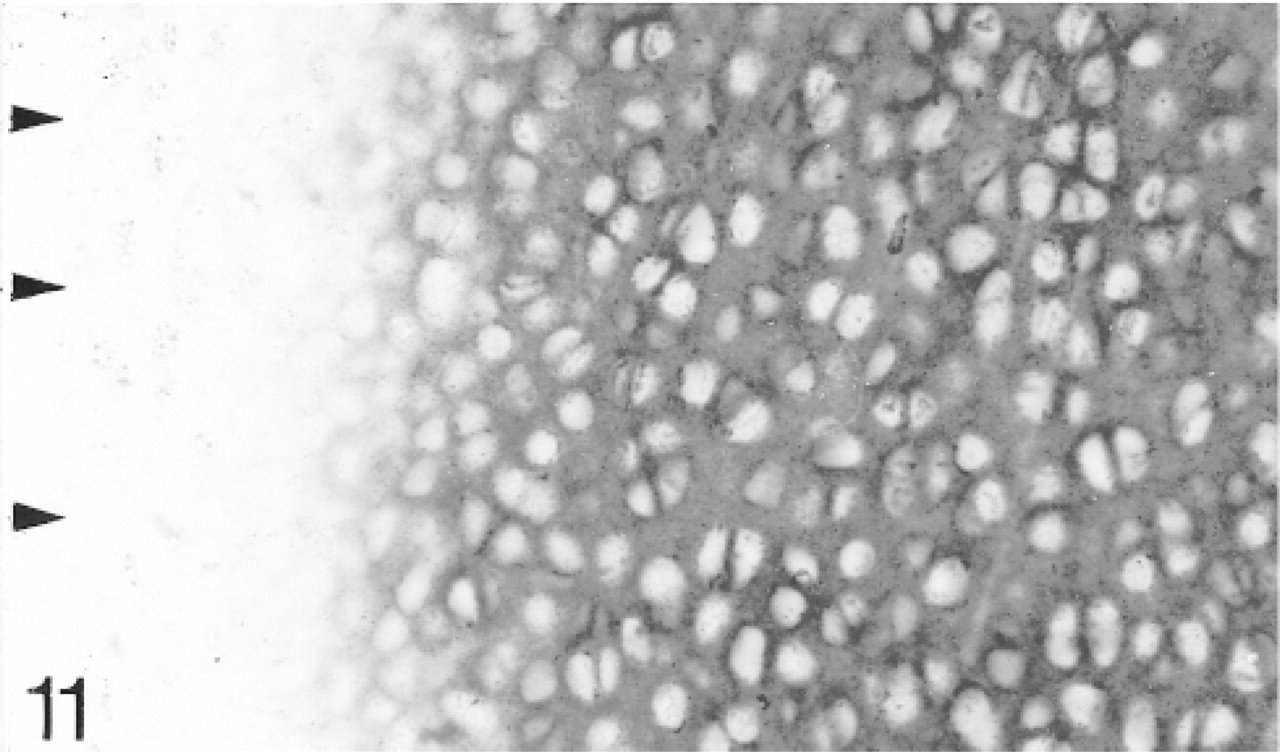
Binding of antibodies to matrilin-1; only the epiphyseal cartilage binds the antibodies. The articular surface is indicated by arrowheads.
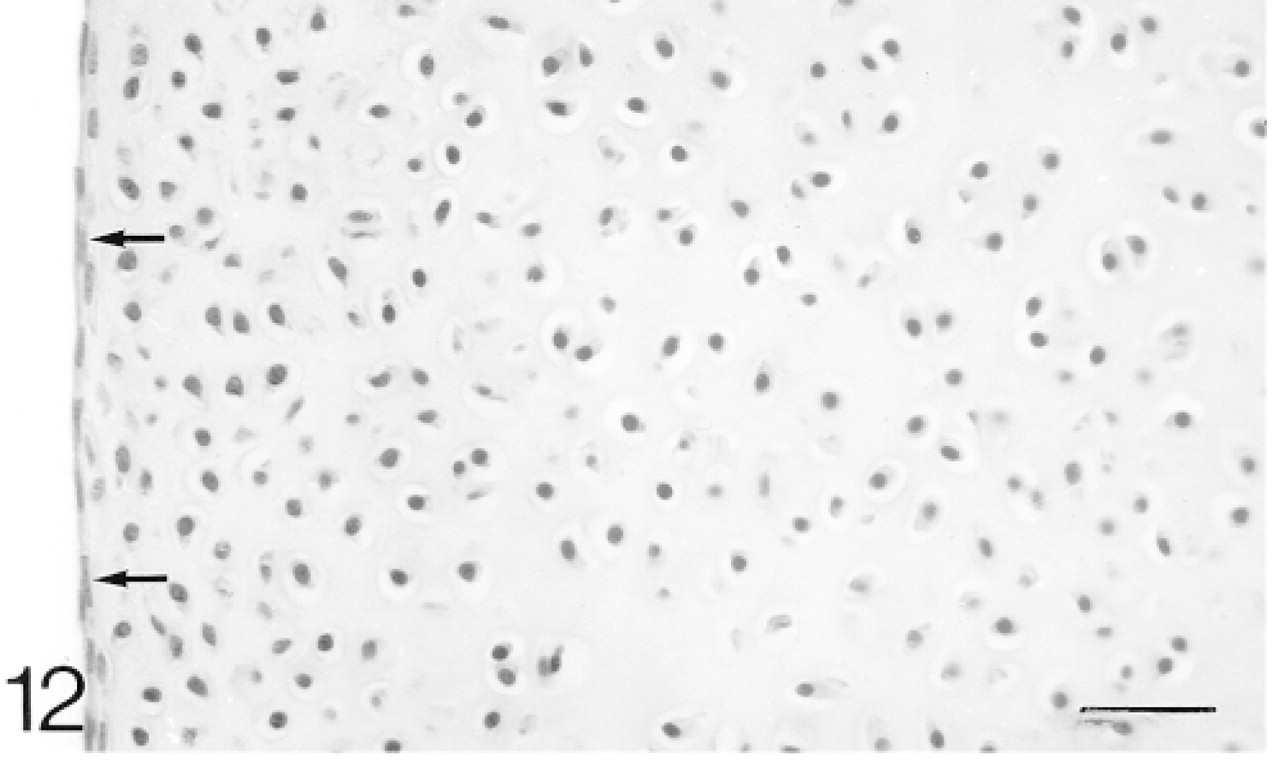
The chondrocytes in the articular cartilage are closer together than those in the epiphyseal cartilage. There is a layer, two cells thick, of flattened cells on the surface (arrows). Hematoxylin and eosin. Bar = 30 μm.
Distributions of biglycan, decorin, and matrilin-1 in articular cartilage of juvenile and adult rabbit knee joints a

aw +, antibodies bound weakly; +, antibodies bound; + +, antibodies bound strongly; -, antibodies not bound; p, pericellular.
bFrom 12–14 weeks, this region is below the tidemark and is calcified.
Neonatal Articular Cartilage
At 1 postnatal week the chondrocytes in the articular cartilage are still closer together than those in the epiphyseal cartilage. There is a layer, about two cells thick, of flattened cells on the surface (Figure 12). The secondary ossification center is developing in the center of the epiphysis. Biglycan antibodies are bound by both the articular and epiphyseal matrices (Figure 14; Table 1) and most chondrocytes are expressing the mRNA for biglycan (Figure 15). Antibodies to decorin are now bound by the articular and epiphyseal matrices, except in the surface layer (Figure 16; Table 1). Most chondrocytes express the mRNA for decorin (Figure 17). Antibodies to matrilin-1 are not bound by the articular cartilage but they are bound throughout the epiphyseal cartilage matrix (Figure 18; Table 1).
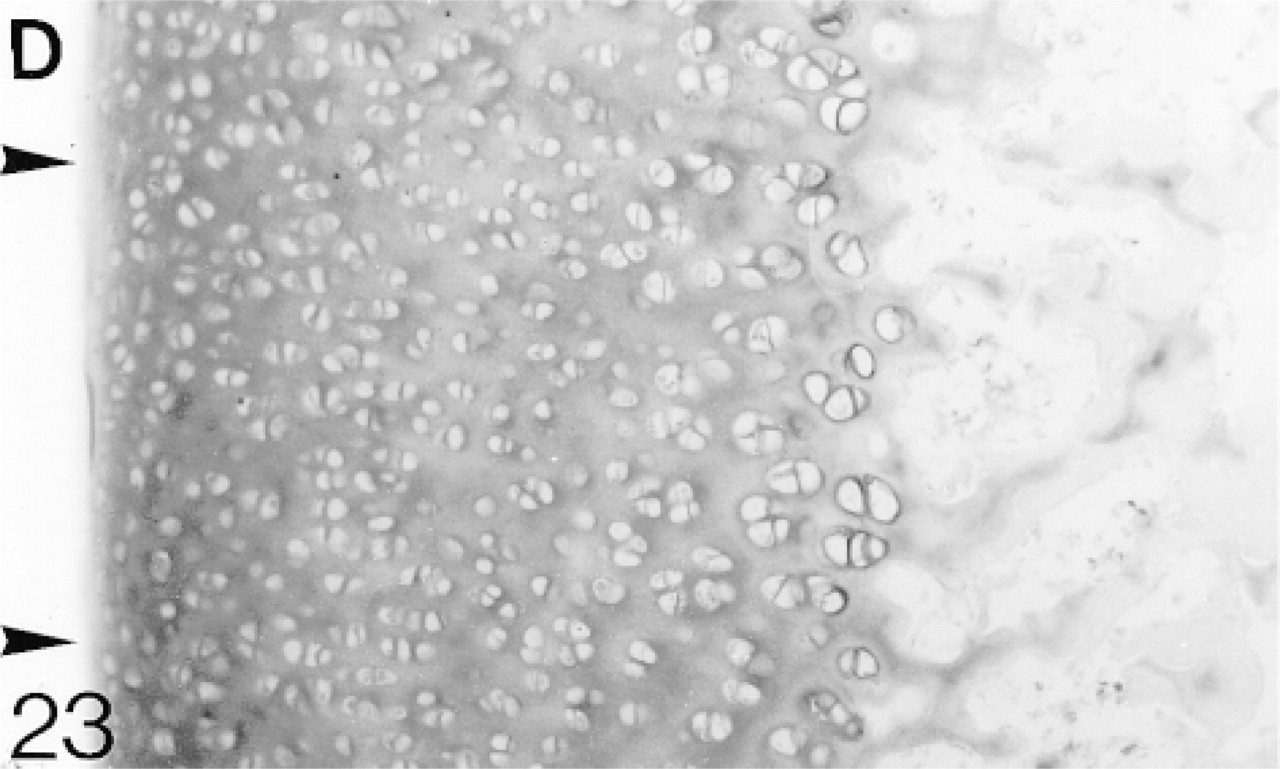
At 3 weeks the articular cartilage, with some underlying epiphyseal cartilage, forms a discrete layer, and by 6 weeks ossification is almost complete (Figure 19). Immediately below the surface layer the cells are randomly arranged, but below this they are beginning to form columns. The surface layer is that containing the flattened cells (one to two cells deep); the upper, middle, and deep regions are indicated in Figure 19 (Poole et al. 1990). From 1 to 6 weeks, biglycan antibody binding gradually increases throughout the articular cartilage (Figure 21; Table 2). At 6 weeks, binding is stronger in the deep matrix and there is prominent pericellular binding around the chondrocytes (Figure 21). Many chondrocytes, particularly in the middle region, express the mRNA for biglycan (Figure 22). Decorin antibody binding is absent from the surface layer. Binding in the upper region of the cartilage is strong, but it decreases slightly towards the deep layer, in which there is strong pericellular binding (Figure 23; Table 2). Many chondrocytes express the mRNA for decorin (Figure 24). Antibodies to matrilin-1 are bound pericellularly in the deep region (Figure 20; Table 2).
Juvenile Cartilage
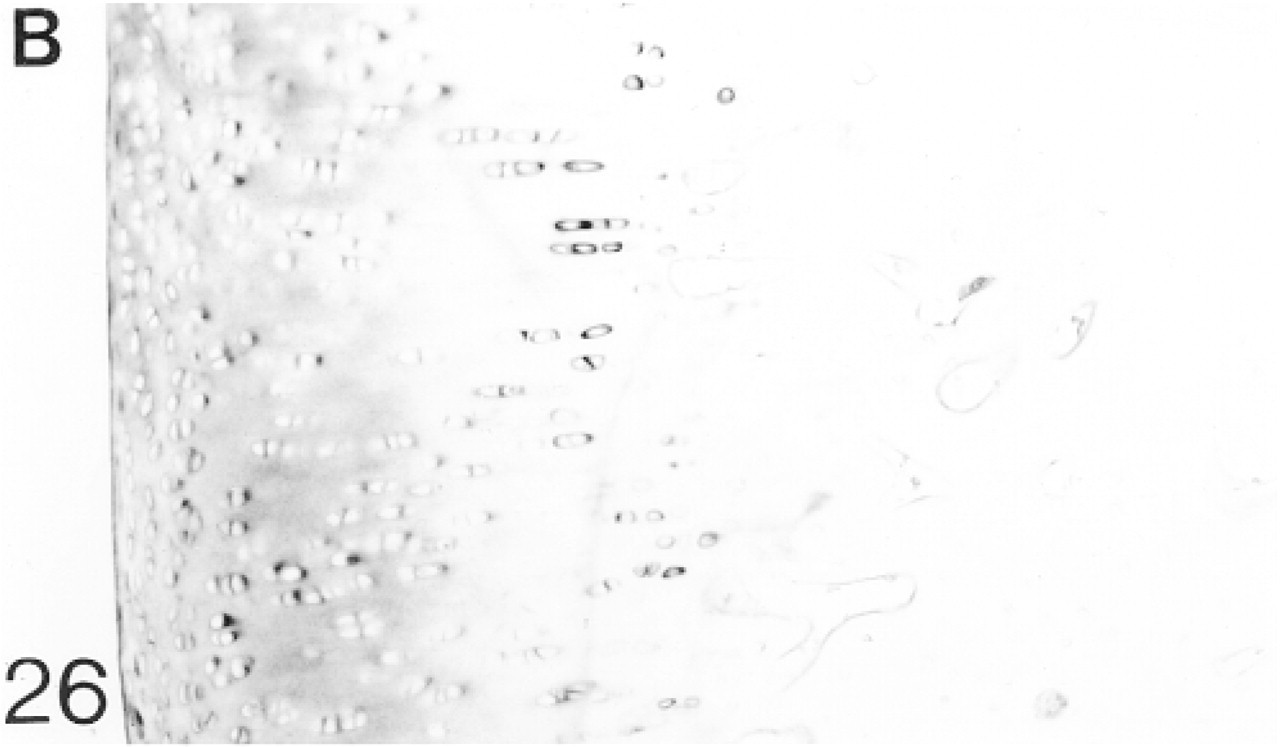
At 12–14 weeks the articular cartilage lies on a layer of compact subchondral bone and the tidemark can now be distinguished (Figure 25). Below the flattened surface cells there is a region in which the cells are irregularly arranged. Below this the cells are arranged in columns of four to six cells. Under the tidemark, the arrangement is again irregular. Biglycan antibody binding is variable at the surface but is stronger below, especially in the territorial matrix. Below the tidemark, binding is pericellular (Figure 26; Table 2). A few chondrocytes express the mRNA for biglycan (not shown). Decorin antibody binding is variable, especially in the upper region of the articular cartilage; it is weak in the surface layer. Pericellular binding occurs throughout the cartilage (Figure 27; Table 2). A few chondrocytes express the mRNA for decorin (not shown). Antibodies to matrilin-1 are not bound by the matrix of the articular cartilage (not shown; Table 2).
Adult Articular Cartilage
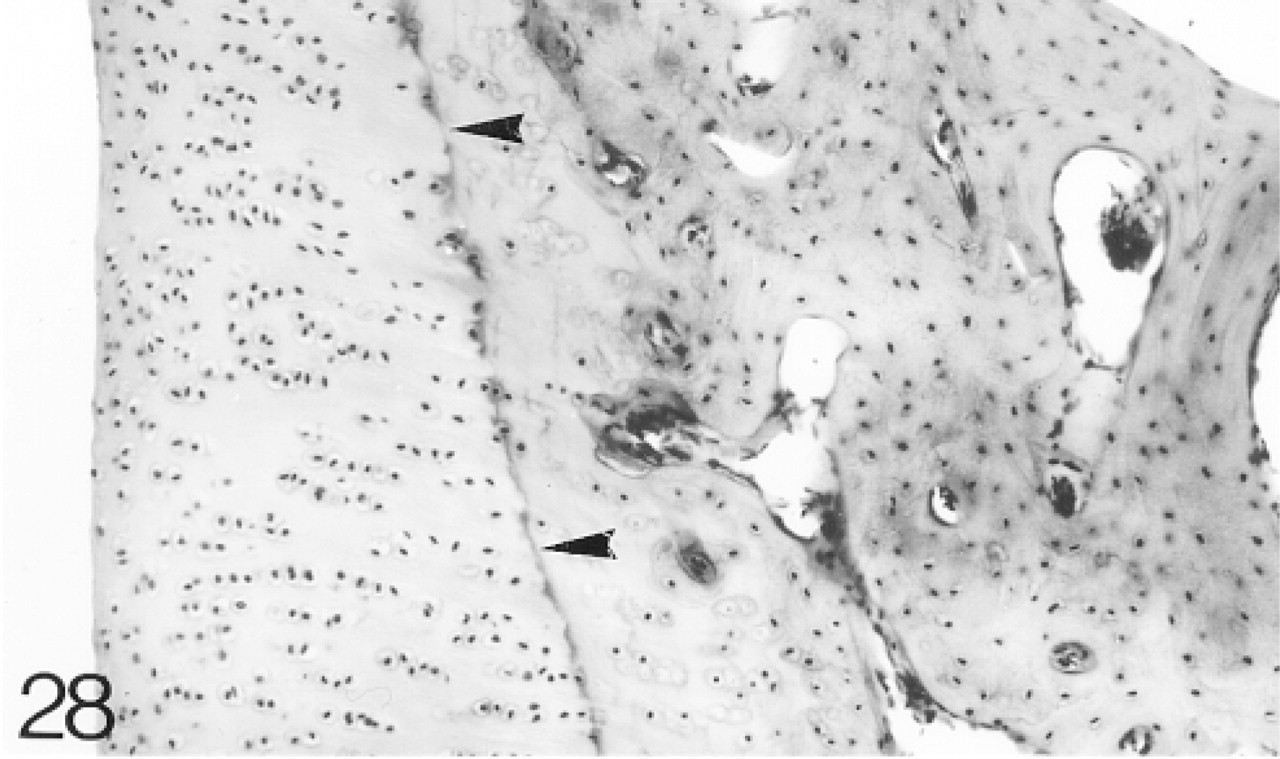
By 8 months the growth plates have closed and the animal is skeletally mature. The articular cartilage is very similar to that at 12–14 weeks (Figure 28) and it does not change appreciably between 8 months and 2 years. Biglycan antibody binding above the tidemark is highly variable and decreases with depth, although there is strong pericellular binding below the tidemark (Figure 29; Table 2). Decorin antibody binding is confined to the interterritorial matrix of the middle and upper regions of the cartilage. Below this region, peri-cellular binding is seen around some chondrocytes (Figure 30; Table 2). A few chondrocytes still express biglycan and decorin mRNAs to 2 years (not shown). Antibodies to matrilin-1 are not bound by the articular cartilage (not shown; Table 2).

Negative control for in situ hybridization using a digoxigenin-labeled sense probe to biglycan.
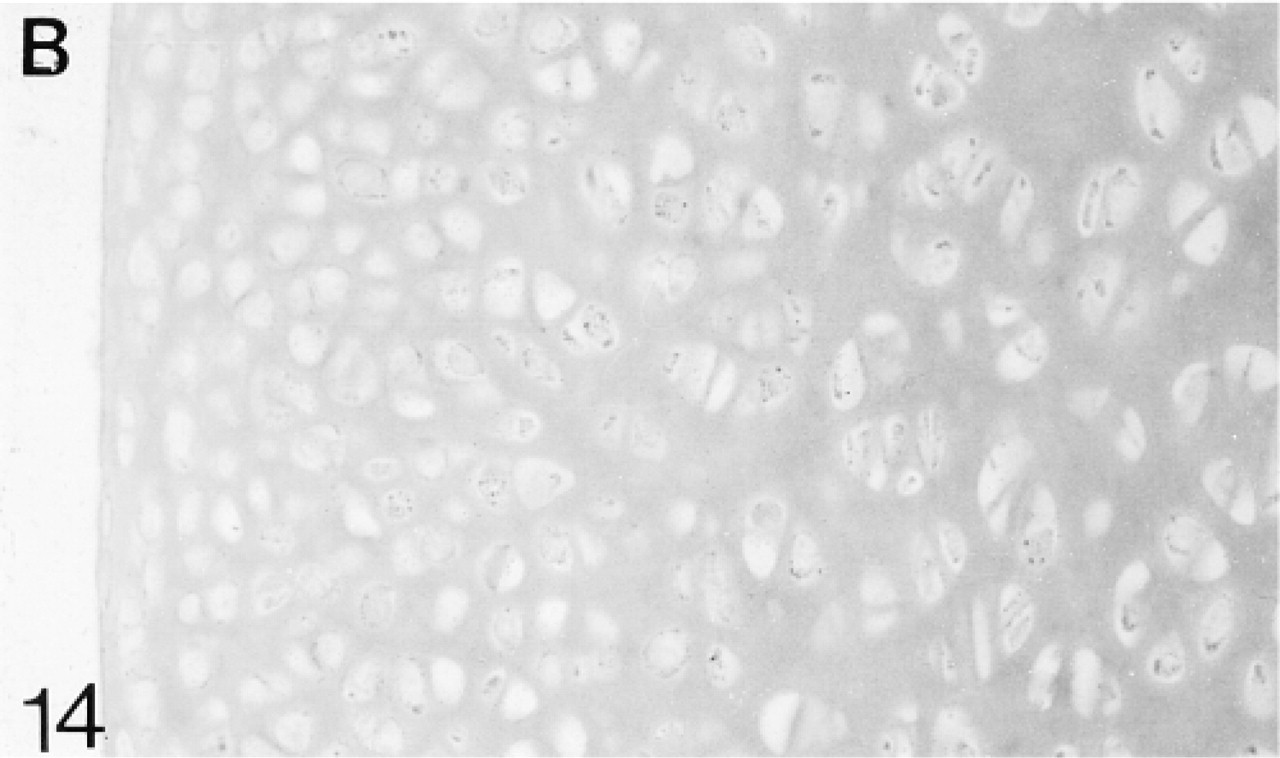
Binding of antibodies to biglycan (B).
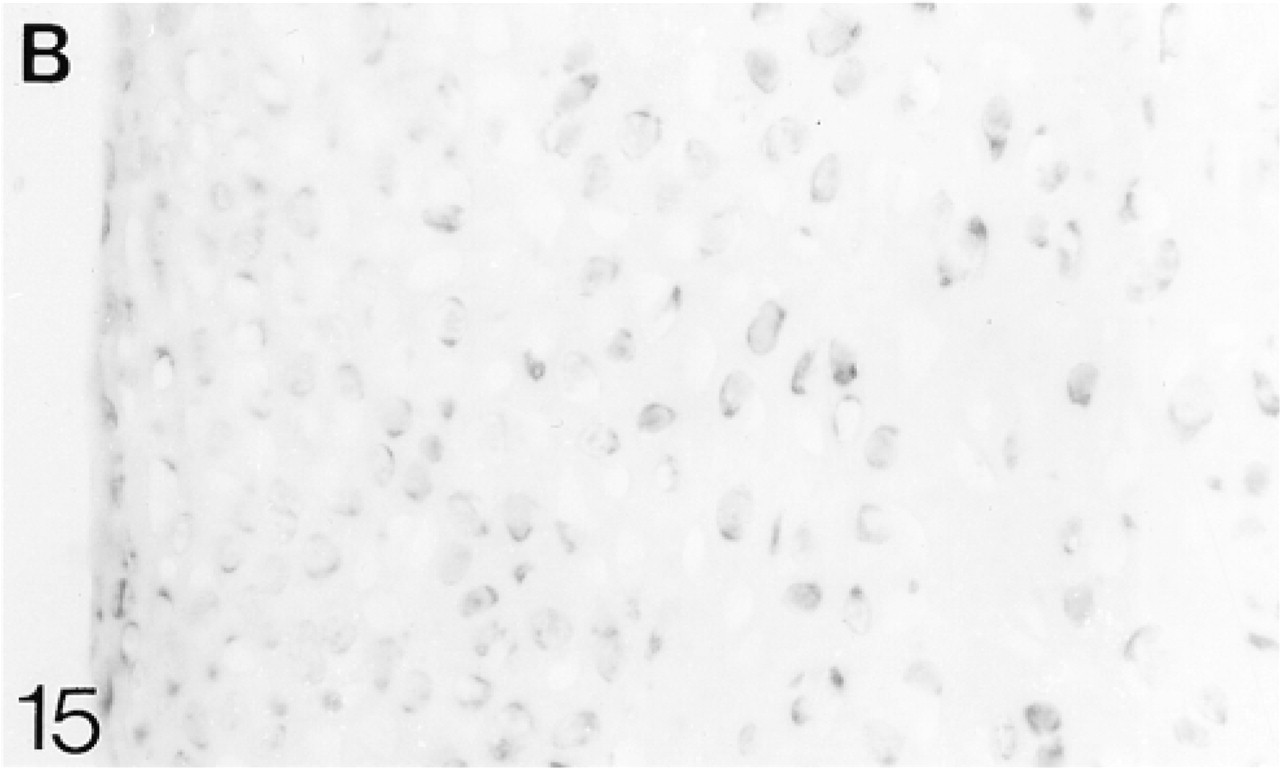
In situ hybridization for biglycan (B) mRNA.
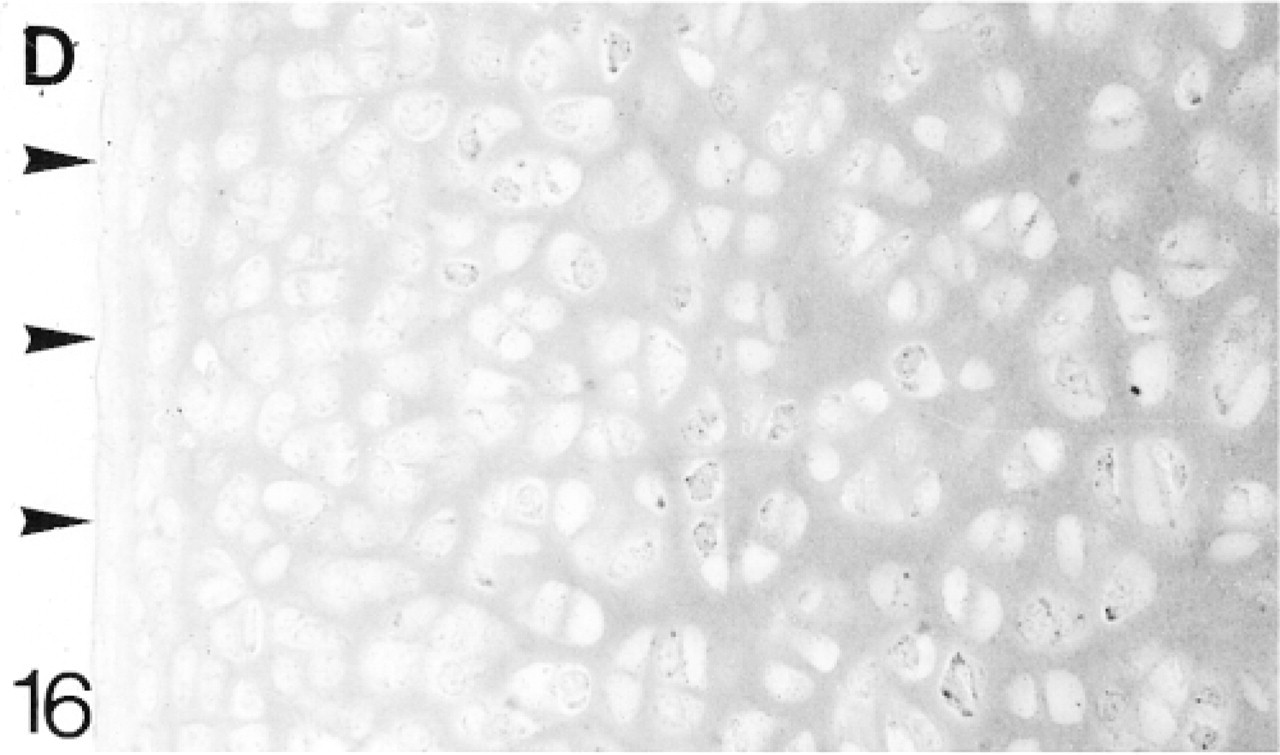
Binding of antibodies to decorin (D). There is no binding in the surface layer (arrowheads indicate the surface).
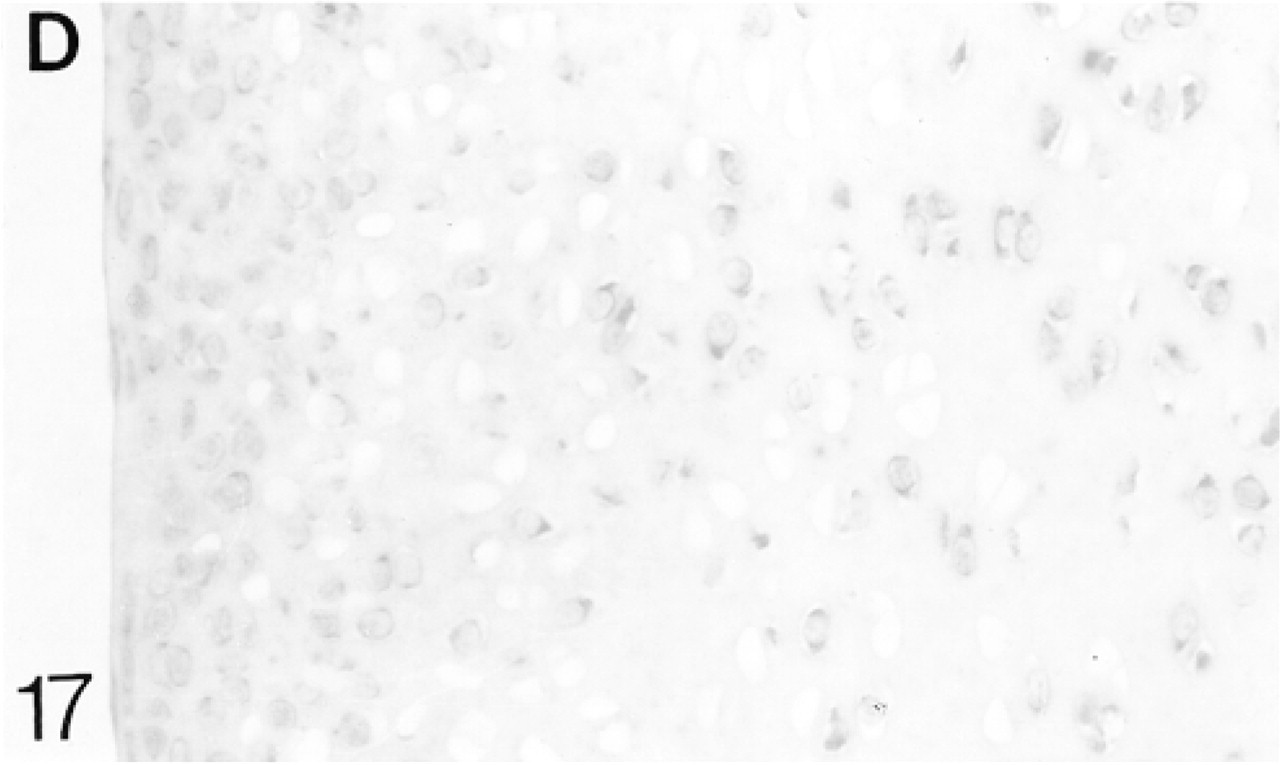
In situ hybridization for decorin (D) mRNA.
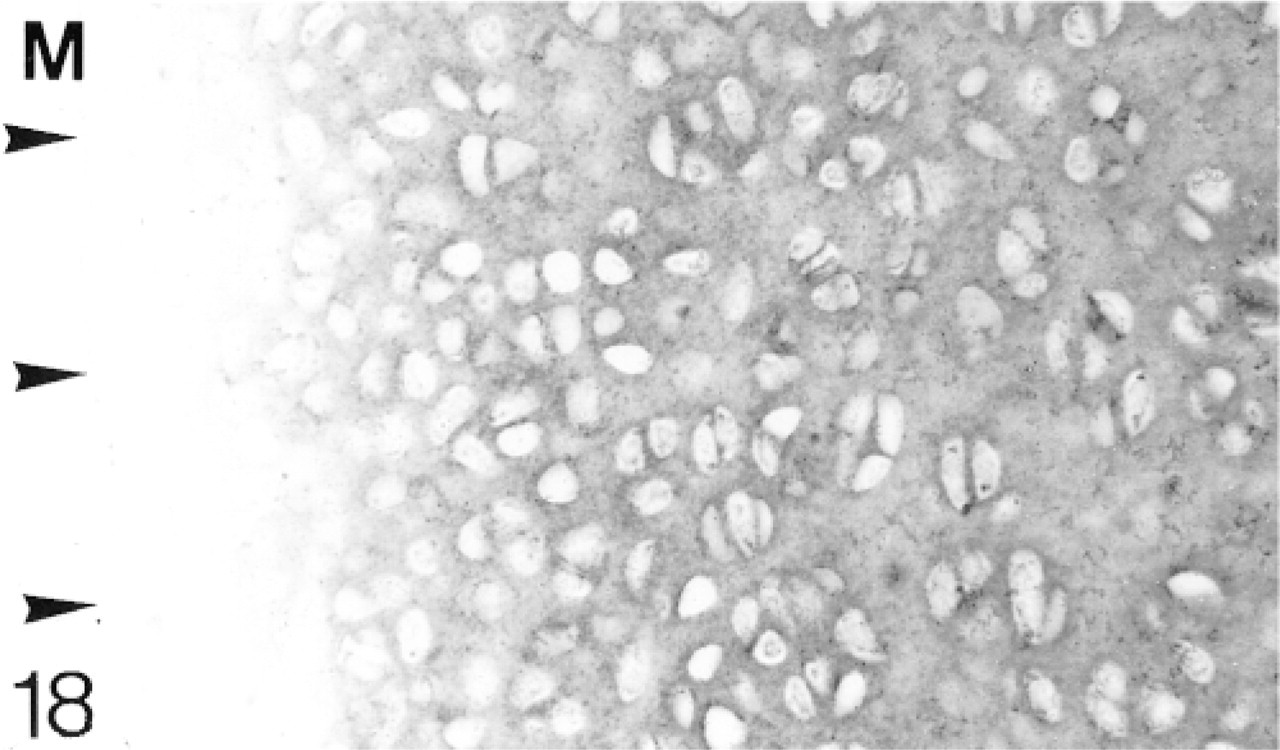
Binding of antibodies to matrilin-1 (M). The articular surface is indicated by arrowheads.
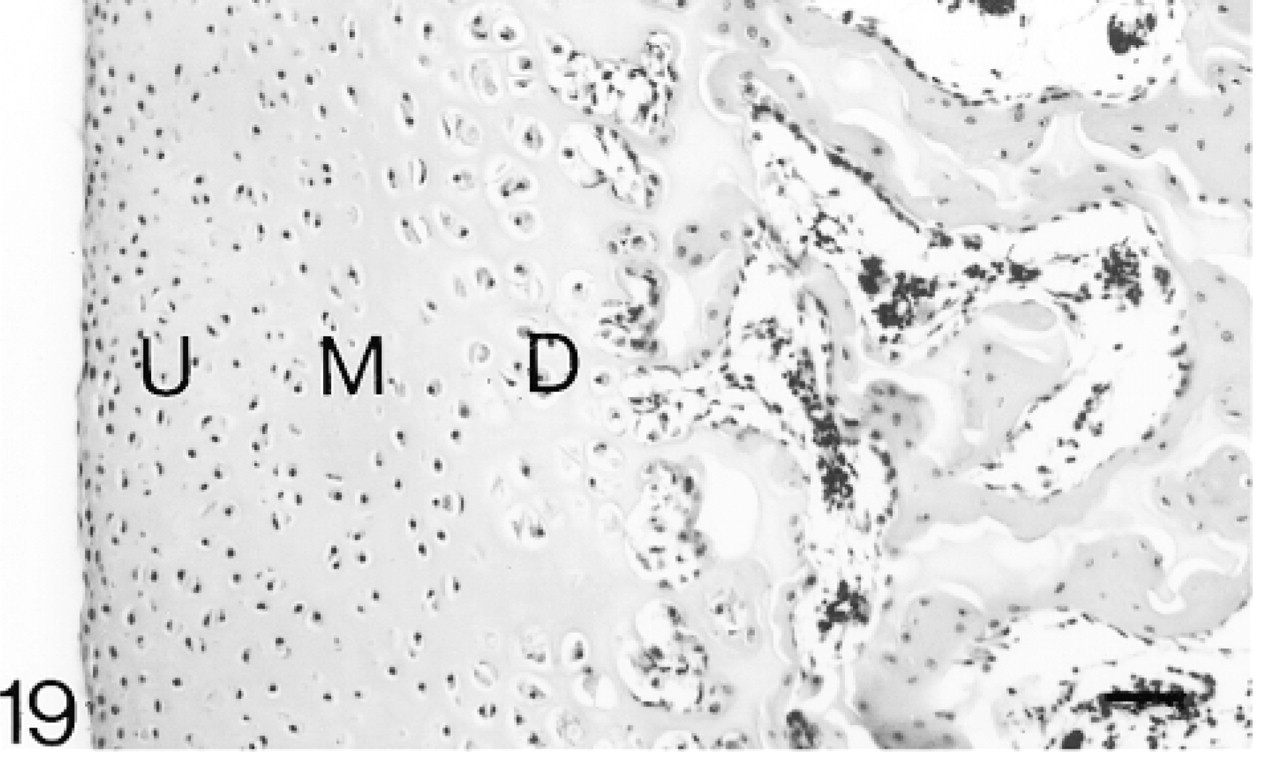
The articular cartilage is a discrete layer and ossification is almost complete. The upper (U), middle (M), and deep (D) regions are indicated. Hematoxylin and eosin. Bar = 50 μm.
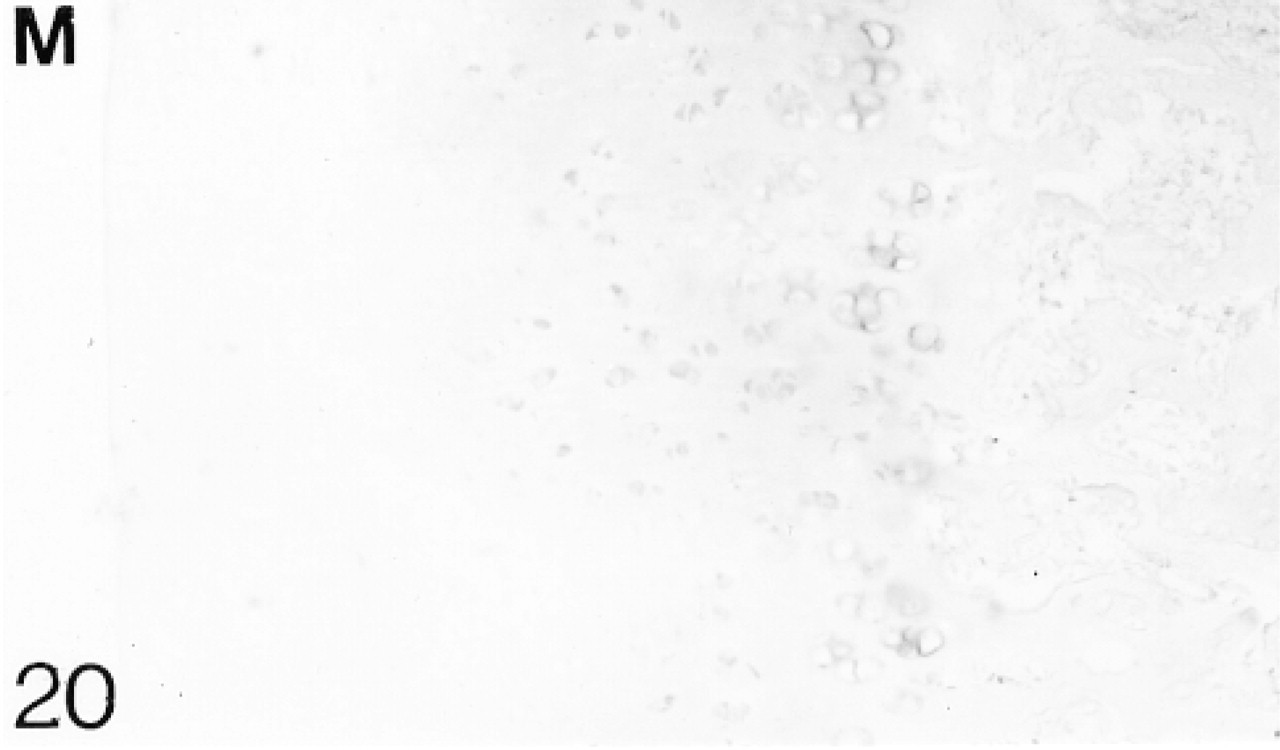
Antibody binding to matrilin-1 (M).
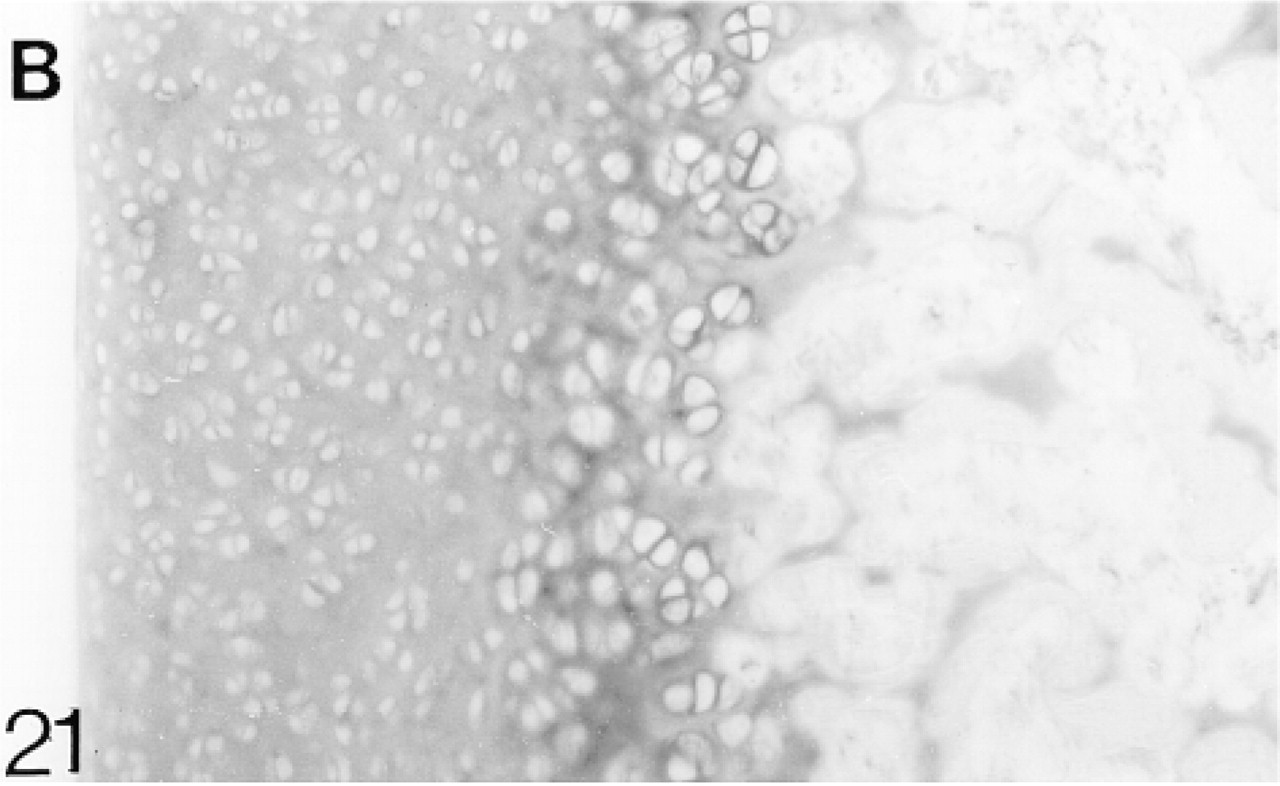
Binding of antibodies to biglycan (B). There is strong pericellular binding in the deep region.
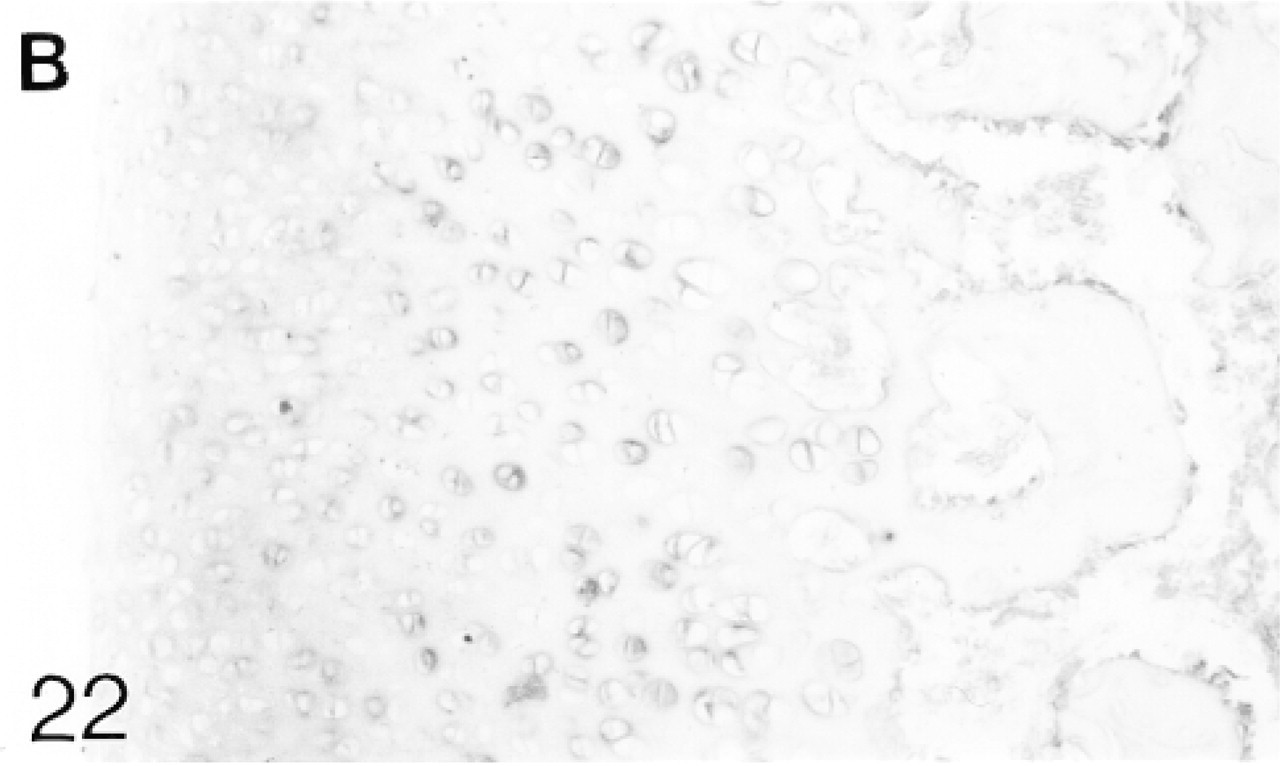
In situ hybridization for biglycan (B) mRNA.
Antibody binding to decorin (D); arrowheads indicate the surface.

In situ hybridization for decorin (D) mRNA.
Discussion
The distributions of the small PGs biglycan and decorin and the glycoprotein matrilin-1, during development and aging of the rabbit knee joint have been examined.

The articular cartilage lies on a layer of compact subchondral bone and the tidemark (arrowheads) can now be distinguished. Hematoxylin and eosin. Bar = 50 μm.
Antibody binding to biglycan (B).

Antibody binding to decorin (D).
The articular cartilage is very similar to that at 12–14 weeks and the tidemark (arrowheads) is clearly visible. Hematoxylin and eosin.

Antibody binding to biglycan.

Antibody binding to decorin.
Biglycan
Before cavitation, biglycan could not be detected in the interzone, i.e., the region from which the articular cartilage develops. Biglycan is present in the deeper regions of the epiphyseal cartilage. A similar distribution was observed in Monodelphis (Archer et al. 1996). Its absence suggests that it is unlikely to be involved in cavitation. Immediately after cavitation in the 25-day rabbit fetus, biglycan is present in the future articular cartilage and the epiphyseal cartilage. The same distribution is found in bovine and Monodelphis joint cartilage at this stage (Bianco et al. 1990; Archer et al. 1996).
At 6 postnatal weeks the articular cartilage forms a discrete layer on the subchondral bone but ossification is incomplete. Biglycan is found throughout the cartilage but antibody binding is stronger near the hyper-trophic cells. As the skeleton matures, biglycan becomes increasingly confined to the pericellular matrix, a distribution similar to that reported previously in bovine and human articular cartilage (Bianco et al. 1990; Miosge et al. 1994). In the adult rabbit (8 months onwards) the distribution of biglycan is variable. In general, it is concentrated in the surface and upper regions and decreases towards the tidemark in a similar manner to that found in human and Monodelphis (Miosge et al. 1994; Archer et al. 1996).
In the 20-day fetus the expression of biglycan mRNA in the epiphyseal and presumptive articular cartilages suggests that the synthesis of biglycan is just beginning. Few chondrocytes in the interzone express the mRNA at this stage, which correlates with the weak antibody binding in the 25-day fetal articular cartilage. The number of articular chondrocytes that express the mRNA for biglycan is higher in the 25-day fetus and in the 1- and 6-week neonate, but the number gradually decreases to 2 years. This suggests that synthesis reaches a maximum in the late fetus and neonate and then declines with age. Studies on isolated human chondrocytes showed that the level of mRNA expression for biglycan decreased with age (Roughley et al. 1994).
The pericellular localization of biglycan in the hypertrophic region of the juvenile rabbit (6 weeks) may be associated with its possible role in the initiation of mineralization (Geerkens et al. 1995; Boskey et al. 1997), but its pericellular distribution at other ages and in other regions suggests another unknown function. Biglycan may be involved in the organization of the matrix, in cell-matrix interactions, and in binding of growth factors (Hildebrand et al. 1992; Buckwalter and Mankin 1997). The strong pericellular antibody binding around the hypertrophic chondrocytes may be attributed either to larger amounts of biglycan in the matrix or to the increased availability of epitopes exposed by the partial breakdown of this matrix in preparation for its removal during endochondral ossification.
Decorin
Before cavitation, decorin is not detected in the matrix or cells of the interzone, but some decorin is present in the deeper regions of the epiphyseal cartilage of the rabbit. In contrast, decorin was found in the chondrocytes and matrix of the interzone of Monodelphis (Archer et al. 1996). After cavitation, decorin gradually accumulates until, in the 6-week neonate, it is evenly distributed in the articular cartilage. A similar pattern is seen in bovine and Monodelphis articular cartilage, but decorin was not identified in human phalangeal joints (Poole et al. 1986; Bianco et al. 1990; Archer et al. 1996). By 12–14 weeks decorin has become concentrated in the upper region of the cartilage but remains in the pericellular matrix of most chondrocytes, irrespective of their position in the cartilage. These observations agree with previous reports of human, bovine, and Monodelphis articular cartilage (Poole et al. 1990; Miosge et al. 1994; Archer et al. 1996). In adult articular cartilage, decorin is localized in the interterritorial matrix of the middle region but is pericellular, below the tidemark. An interterritorial distribution was described previously in human and bovine articular cartilage (Bianco et al. 1990; Poole et al. 1990).
In the 20-day fetus, the expression of decorin mRNA by cells in the epiphyseal cartilage and inter-zone suggests that the synthesis of decorin is just beginning. Very few chondrocytes in the interzone express the mRNA at this stage, which correlates with the very weak antibody binding in the presumptive articular cartilage of the 25-day fetus. The number of articular chondrocytes that express the mRNA is higher from the 25-day fetus to the 6-week neonate, but they then decrease in number to 2 years. Thus, synthesis reaches a maximum in the late fetus and neonate and then declines with age.
Decorin binds to Type II collagen and may be involved in the control of fibrillogenesis (Hedbom and Heinegård 1993). Type II collagen is not present in the interzone of the developing rabbit joint (Bland and Ashhurst 1996) but, after cavitation, it gradually accumulates in the cartilage matrix at the same time as decorin. The similar distributions of decorin and Type II collagen throughout development and aging support the suggested interactions of decorin with fibrillar collagens. In addition to its role in the organization of the matrix, decorin may also act as an intermediary in the binding of growth factors and in cell-matrix interactions (Hildebrand et al. 1992; Buckwalter and Mankin 1997).
Matrilin-1
Matrilin-1 is absent from the interzone and therefore from the presumptive articular cartilage, but it is widely distributed in the epiphyseal and diaphyseal cartilages at 20 days. Its presence in this cartilage precedes that of the small PGs. At 6 postnatal weeks it is present only in the pericellular matrix of the hypertrophic layer, i.e., the remnants of the epiphyseal cartilage. On completion of endochondral ossification, only articular cartilage, which never contained matrilin-1, remains. This distribution agrees with that in developing human and mouse articular cartilage (Aszodi et al. 1994; Mundlos and Zabel 1994).
Matrilin-1 protein and mRNA accumulate during skeletogenesis in the growth plate of human, mouse, and chicken (Aszodi et al. 1994; Mundlos and Zabel 1994). Its mRNA is expressed at high levels in the late proliferative and upper hypertrophic zones (Aszodi et al. 1994; Chen et al. 1995).
The distributions of matrilin-1 and Type II collagen are very similar before cavitation in the chicken and rabbit (Winterbottom et al. 1992; Bland and Ashhurst 1996). Matrilin-1 is associated with two types of fibril. One type contains only matrilin-1, whereas the other is a mixture of matrilin-1 and Type II collagen (Winterbottom et al. 1992). The distribution of matrilin-1 cannot be correlated with any other fibrillar collagen.
Matrilin-1 enables the articular and epiphyseal cartilages to be distinguished in a manner analogous to the pericellular distribution of Type V collagen in the interzone and in developing and aging articular cartilage (Bland and Ashhurst 1996). The localization of matrilin-1 in the hypertrophic cartilage confirms that only epiphyseal cartilage is removed during ossification. Once ossification is complete, only the cartilage formed from the presumptive articular cartilage in the fetus remains. The absence of matrilin-1 from cartilaginous structures exposed to strong compressive forces indicates that this protein is clearly different from other cartilage proteins, and its role remains unknown.
Conclusions
Neither biglycan nor decorin was detected in the inter-zone of the developing rabbit knee joint. Therefore, the chondroitin sulfate found in the interzone of developing human knee joints by Andersen (1961) is part of another PG. The large PG aggrecan is a possible candidate, but its presence has not been reported in the interzone. Hyaluronan is found in the interzone and believed to play a role in the formation of the cavity, but it is not sulfated (Edwards et al. 1994; Pitsillides et al. 1995; Dowthwaite et al. 1998).
In the adult articular cartilage, the variable distributions of biglycan and decorin could reflect the different weight-bearing properties in different regions of the knee joint. Both biglycan and decorin synthesis are increased in load-bearing regions of canine, equine, and ovine articular cartilage (Adams et al. 1995; Dourado et al. 1996; Little and Ghosh 1997; Little et al. 1996,1997).
Loss of biglycan and decorin occurs in human osteoarthritis (Carroll 1989; Pettipher et al. 1989; Witson-Prehm et al. 1992; Cs-Szabo et al. 1997; van de Loo et al. 1998). In contrast, Poole et al. (1990) found no change in the overall content of biglycan and decorin, but changes in their distributions leads to their loss from the superficial layers and their increase in the deep cartilage. Although matrilin-1 is not present in normal articular cartilage it appears in osteoarthritic and rheumatoid cartilage (Saxne and Heinegård 1989; Okimura et al. 1997). Its presence indicates another property that distinguishes diseased from normal articular cartilage.
The results presented here and those of Bland and Ashhurst (1996) show how the distributions of both collagens and small PGs vary during development and aging in rabbit articular cartilage. In addition, they suggest that the articular chondrocytes are a distinct population of cells from the earliest stages of joint development. Such data are important for a greater understanding of the changes that take place during arthritic disease processes. That the arthritic cartilage acquires some of the constituents, e.g., matrilin-1, of epiphyseal cartilage is an intriguing enigma. The question remains: When do changes, such as the synthesis of matrilin-1, occur in relation to the onset of the disease? The present study provides some basic information essential for studies of the development of disease.
Footnotes
Acknowledgements
Supported by the Arthritis Research Campaign (grant AO526).
We wish to acknowledge Prof D. Heinegård and Prof M. Paulsson for their kind gifts of antibodies. The technical expertise of Ms Y. Bland has been invaluable.