
Abstract
This study demonstrates for the first time that human articular chondrocytes express osteogenic protein-1 (OP-1). OP-1 was originally purified from bone matrix and was shown to induce cartilage and bone formation. Both OP-1 protein and message were present in human normal and osteoarthritic (OA) cartilages. OP-1 mRNA was upregulated in OA cartilage compared with normal adult tissues. However, the level of mature OP-1 protein in the same OA tissues was downregulated, whereas the pro-OP-1 remained high. Moreover, these two forms of OP-1 were localized in an inverted manner. Mature OP-1 was primarily detected in the superficial layer, whereas the pro-form was mostly in the deep layer of cartilage. The presence of pro- and mature OP-1 in extracts of normal and OA cartilages was confirmed by Western blotting. These findings imply that articular chondrocytes continue to express and synthesize OP-1 throughout adulthood. The observed patterns of the distribution of pro- and mature OP-1 also suggest differences in the processing of this molecule by normal and OA chondrocytes and by the cells in the superficial and deep layers. Distinct distribution of OP-1 and its potential activation in deep zones and regions of cloning in OA cartilages may provide clues to the potential involvement of endogenous OP-1 in repair mechanisms.
Keywords
C
Bovine osteogenic protein was originally purified from bone matrix as a mixture of homodimers of two members of the BMP family. One component was identified as the protein product of the bovine equivalent of the human osteogenic protein-1 (OP-1; BMP-7) gene and the other was identified as the protein product of the bovine equivalent of the human BMP-2A gene (Sampath et al. 1990). Human recombinant OP-1 was purified from transfected CHO cells as a disulfide-linked homodimer with an apparent molecular weight range of 46-34 kD. On reduction, the protein migrates as 23-,19-, and 17-kD monomers. These monomers each contain amino acid residues 293–431 and differ only by the extent of glycosylation. This form of OP-1 is referred to as the mature domain of OP-1. Residues 29-292 from the amino acid sequence described by Özkaynak et al. (1990) are termed the pro-domain. The pro-OP-1 molecule consists of a complex of two pro-domains that, after the proteolytic processing, remain noncovalently associated with the disulfide linked mature dimer (Jones et al. 1994). Expression of full-length OP-1 cDNA in CHO cells results in translation of OP-1 as a pre-pro-protein, which undergoes a subsequent proteolytic processing at an Arg-Xaa-Xaa-Arg site, yielding a mature disulfide linked homodimer (Jones et al. 1994).
Recent studies from our department (Flechtenmacher et al. 1996; Huch et al. 1997; Koepp et al. 1999) have focused on the potential role of exogenous OP-1 in human and bovine cartilage homeostasis and repair. We showed that human recombinant OP-1 can (a) stimulate synthesis of aggrecan and collagen Type II by human articular chondrocytes (Flechtenmacher et al. 1996), (b) counteract the deleterious effects of interleukin-1 on the metabolism of human articular chondrocytes (Huch et al. 1997), and (c) block bovine cartilage damage mediated by fibronectin fragments (Koepp et al. 1999). In these studies, chondrocytes were either cultured as explants or were isolated from their matrix and cultured in alginate beads; OP-1 was added exogenously.
Members of the BMP family have also been identified in different connective tissues, but mostly during the process of development. OP-1 was detected in the ectodermal epithelium of the fetal mouse fore- and hindlimbs, heart, teeth, intestinal epithelium, perichondrium, hypertrophic chondrocytes, and the periosteum/osteoblast layer of developing human bones (Helder et al. 1995). BMP-2, −4, and −7 were colocalized in mature and immature chondrocytes surrounding the calcified zone of ossified ligamentum flavum (Hayashi et al. 1997). OP-1 was found during tooth development in rodents (Helder et al. 1998), and a 17-fold increase of OP-1 was reported in high-grade chondrosarcomas in comparison to low-grade and benign tumors (Weber et al. 1998). Similar results were obtained in our laboratory, where the expression of OP-1 (mRNA and protein) was detected only in high-grade human chondrosarcomas but not in low-grade or benign tumors (Chubinskaya et al. 1996b, 1998). All these findings prompted us to investigate if OP-1 can be detected in human adult articular cartilage and if there are changes in the patterns of expression and localization of OP-1 in cartilage of different pathophysiological states.
Materials and Methods
Reagents
Human recombinant mature and pro-OP-1, polyclonal antiserum against pro-domain of OP-1 (R2854), and anti-OP-1 monoclonal antibodies (12G3 and 1B12) were provided by Creative BioMolecules and Stryker Biotech (Hopkinton, MA; Sampath et al. 1992). Electrophoresis grade reagents were from Bio-Rad (Hercules, CA). Chemicals, either reagent or molecular biology grade, were purchased from Sigma Chemical (St Louis, MO) unless otherwise noted. Keratanase (Pseudomonas sp; EC 3.2.1.103), keratanase II (Bacillus sp), and chondroitinase ABC (Proteus vulgaris; EC 4.2.2.2) were obtained from Seikagaku (Tokyo, Japan). Pronase was purchased from Calbiochem (La Jolla, CA), collagenase P (Clostridium histolyticum) from Boehringer Mannheim (Indianapolis, IN), and fetal bovine serum (FBS) from Hyclone (Logan, UT). Low-viscosity alginate (Keltone, LV) was obtained from Kelco (Chicago, IL). Dulbecco's modified Eagle's medium/Ham's F12 medium (DMEM/F12) was obtained in a 1/1 powder from Gibco (Grand Island, NY).
Tissue Acquisition
Normal Cartilage. Full-thickness articular cartilage was dissected within 24 hr of death from the load-bearing regions of femoral condyles of human donors with no history of joint disease. Tissues (from both men and women, ranging from newborn to 84 years of age) were obtained through the Regional Organ Bank of Illinois.
Osteoarthritic Cartilage. Human OA cartilage adjacent to eroded areas was obtained as surgical specimens from patients (59-88 years of age, both men and women) undergoing total knee arthroplasty due to advanced OA. Within 3 hr after surgery, full-thickness noncalcified cartilage was dissected from the femoral condyle. Osteophytes were excluded from the studies because this tissue represents a completely different biology compared to primary OA cartilage. Adjacent pieces of the cartilage from both patients and donors were either processed for histology, immunohistochemistry, and in situ hybridization or they were used for total RNA extraction for RT-PCR or for protein extraction for Western blot analyses. Cartilages from four human donors (newborn and adult) were used for isolation and culture of chondrocytes in alginate beads.
Preparation of Chondrocytes and Alginate Beads
Chondrocytes were prepared for culture in alginate beads as previously described (Mok et al. 1994). Briefly, the cells were released from their native matrix by sequential digestion in pronase (0.2%) for 1 hr and then with collagenase P (0.025%) overnight in DMEM/F12 with 5% FBS at 37C. The isolated chondrocytes were suspended at a density of 4 times; 106 per ml in a 1.2% solution of sterile alginate in 0.15 M NaCl. The cell suspension was slowly expressed through a 22-gauge needle and dropped into a 102 mM CaCl2 solution. The beads, with approximately 40,000 cells/bead (diameter 3 mm), were allowed to polymerize for 10 min and washed two times with 0.15 M NaCl, followed by two washes in DMEM/F12. The beads were then transferred to medium (500 beads/25 ml/dish) containing DMEM/F12, 50 μg/ml gentamycin, 10% FBS, and 25 μg/ml ascorbic acid. The beads were cultured at 37C in a humidified atmosphere of 5% CO2 for 7 days, with medium replaced daily.
Histological Processing of Cartilage Tissue
Cartilage from newborn donors and articular cartilage from adult donors (~3 times; 3 mm2 pieces) were removed from the joints free from subchondral bone or calcified cartilage and processed for in situ hybridization and immunohistochemistry using RNase-free solutions as previously described (Chubinskaya et al. 1996a).
Processing of Alginate Beads for In situ Hybridization and Histology
The beads were processed as previously described (Petit et al. 1996) except that RNase-free solutions were used. The beads were fixed in 4% paraformaldehyde, 0.1 M cacodylate buffer, pH 7.4, containing 10 mM CaCl2 for 4 hr at 20C and were then washed overnight at 4C in 0.1 M cacodylate buffer (pH 7.4) containing 50 mM BaCl2. The beads were dehydrated through alcohols and xylene and embedded in paraffin.
Sections of cartilage tissue or alginate bead (6 μm) were utilized for in situ hybridization and immunohistochemistry as described (Chubinskaya et al. in press).
Oligonucleotide Probes and PCR Primers for OP-1
The OP-1 sequence has about 30% homology to the sequence of TGF-β and about 60% homology to BMP-2. Therefore, PCR primers and oligonucleotide probes for in situ hybridization were primarily designed in the most conserved and specific area of the sequence in the nontranslated region of the OP-1. Four antisense and sense oligonucleotide probes and primers were designed on the basis of OP-1 sequence (accession X51801) to fulfill requirements of both in situ hybridization and RT-PCR: (a) 21-mer, antisense, location 1810-1830, 5′-TTTTCCTTTCGCACAGACACC-3′; (b) 24-mer, antisense, location 1499-1522, 5′-GGGAAGGTCTCACAAAAGGCAGTT-3′; (c) 20-mer, sense, location 1328-1347, 5′-TGCCATCTCCGTCCTCTACT-3′; and (d) 23-mer, sense, location 1518-1540, 5′-TTCCCCTCCCTATCCCCAACTTT-3′. The specificity of these probes was compared with sequence data available from PC/GENE on the human DNA database. Sequences were checked using Oligo software program, Version 5 for Windows. All oligonucleotide probes were synthesized and purified by Research Genetics (Huntsville, AL). For in situ hybridization, probes were 3′-end-labeled with 5′-[α-thiol-35S]-dCTP (NEN Du Pont; Boston, MA) using terminal deoxynucleotidyl transferase (Promega; Madison, WI). Hybridization was done under conditions of highest stringency.
In Situ Hybridization
In situ hybridization was done according to Sandell et al. (1991) as adapted by our laboratory (Chubinskaya et al. 1996a). The major modification for chondrocytes in alginate beads was the presence of divalent ions Ca+2 or Ba+2 in the fixation or buffer washing solutions respectively (Chubinskaya et al. in press). After in situ hybridization, the slides were dipped in Kodak NTB2 emulsion at 42C. The emulsion was exposed for 3-7 days at 4C, developed with Kodak D19, and fixed with Kodak fixative. The sections then were stained with 0.2% aqueous cresyl violet to visualize the cells and matrix. Coverslips were mounted with Acrytol. To enhance the visualization of the binding of radiolabeled oligonucleotide probes to mRNA within the cells, the tissue sections were viewed by darkfield microscopy (Nikon, Microphot-FXA).
In Situ Hybridization Controls
Positive Controls. Human chondrosarcoma tissue (Weber et al. 1998) and sequencing of the PCR products were used as positive controls.
Negative Controls. Sense primers and competitive inhibition control served as negative controls. Competitive inhibition control (cold hybridization) was performed by mixing radiolabeled probe with unlabeled (cold) probe in a ratio of 1:1 and 1:2 (amount of isotope was kept constant) as previously described (Chubinskaya et al. 1996a).
Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Total RNA was extracted directly from cartilage tissue with acid-guanidinium thiocyanate as previously described (Cs-Szabo et al. 1997). Specific primer pairs were constructed for OP-1 (Primers a and d, listed above) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). These primer pairs were designed to yield PCR products of different sizes (319 bp for GAPDH and 313 bp for OP-1). RT-PCR was done using 1 μg of total RNA. Reverse transcription was done as described by Cs-Szabo et al. (1997). Five μl of the mixture was subjected to PCR using Taq DNA polymerase (Promega) in the presence of specific upstream and downstream primers (15 pM each). To perform RT-PCR at the optimal conditions and to stay within the logarithmically linear product formation, 33 cycles were chosen (45 sec at 95C, 30 sec at 60C of annealing temperature and 45 sec at 72C for the primers used), followed by the final extension for 6 min at 72C. PCR products were separated in 3% Metaphor agarose gels (FMC BioProducts; Rockland, ME) and visualized by ethidium bromide staining. Densities of the bands were measured using a Fluor-S MultiImager (Bio-Rad) with attached software program. The densities of the OP-1 bands were normalized to the densities of the GAPDH bands to control variability among samples. In addition, PCR products of the OP-1 gene from one normal cartilage and one OA cartilage were sequenced and identified (Ana-Gen Technologies; Palo Alto, CA). For each experimental group (newborn, normal adult, and OA), the result is reported as the mean ± SD of analysis of three different cartilages. The ratio for each cartilage was a result of triplicate determinations.
Cartilage Extraction
Articular cartilage was briefly washed with 0.9% saline, homogenized, and extracted in 1 M guanidine hydrochloride in the presence of proteinase inhibitors (0.1 M ∊-aminocaproic acid, 0.05 M sodium acetate, 0.005 M EDTA, 0.005 M benzamidine chloride, 0.01 M N-ethylmaleimide, 0.0005 M phenymethanesulfonyl fluoride) at pH 7.4. Samples were extracted for 3 hr on ice and the extracts were dialyzed against water for 48 hr at 2-8C, lyophilized, and solubilized in sample buffer [0.01 M Tris-HCl, pH 6.5, 1% sodium dodecyl sulfate (SDS), 10% glycerol, 0.016% bromphenol blue, 8 M urea, 0.2 M sodium chloride, no dithioerythritol (DTT), pH 6.8].Protein concentrations were determined by a colorimetric assay using BSA as a standard (Pierce; Rockford, IL).
Western Blot Analyses
Aliquots of samples (30 μg of protein/lane) were analyzed by electrophoresis and immunoblotting without prior digestion. Samples were heated in boiling water for 5 min and loaded onto a 12% SDS-polyacrylamide gel (PAGE) in a buffer containing 0.2 M glycine, 25 mM Tris, and 3.5 mM SDS. Proteins were transferred from SDS-PAGE gels onto nitrocellulose transfer membranes overnight at 20 V, 4-8C in a buffer containing 12 mM Tris (pH 7.4), 6 mM Na-acetate, and 0.3 mM EDTA at pH 7.4. Immunoblot analyses were done either with polyclonal antiserum against pro-domain OP-1 (pro-domain primary antibody; R 2854) or with two anti-OP-1 (mature-OP-1) monoclonal antibodies, 12G3 and 1B12 (Vukicevic et al. 1994). Nonspecific binding sites were blocked with 5% skim milk powder (Carnation). The blots were then incubated with 12G3 or 1B12 primary antibodies at 1:2000 dilution or pro-domain, R2854, antibody at 1:250 dilution, and a horseradish peroxidase-conjugated goat anti-mouse IgG or anti-rabbit IgG (Cappel, Organon Teknika; West Chester, PA) as a secondary antibody (1:4000 dilution). To increase the sensitivity of Western blots, Enhanced ChemiLuminescent kits (Amersham Life Science; Poole, UK) were applied. As positive controls, human recombinant pro-OP-1 and mature OP-1 were used. Sample buffer, secondary antibodies alone, or primary antibody preabsorbed with recombinant mature or pro-OP-1 served as negative controls.
Immunohistochemistry
For immunohistochemistry, cartilage sections (6 μm) were fixed in 4% paraformaldehyde, dehydrated, and embedded in paraffin. Before incubation with primary antibodies, tissue sections were digested with keratanase (0.01 U/ml), keratanase II (0.0001 U/ml), and chondroitinase ABC (0.01 U/ml) in 100 mM Tris/50 mM Na-acetate buffer (pH 6.5) at 37C for 90 min to increase the penetration of antibodies into cartilage. Primary antibodies were applied at a 1:100 dilution. Secondary antibodies, peroxidase-labeled goat anti-mouse or anti-rabbit immunoglobulin G (Organon Teknika-Cappel), or biotinylated antibodies from ImmunoPure ABC Alkaline Phosphatase Mouse or Rabbit IgG Staining Kits (Pierce) were used at the same, 1:100 dilution. For controls, the primary antibody was replaced with either normal serum, secondary antibody alone, or primary antibody preabsorbed with OP-1. The colored reaction product was developed either with diaminobenzidine/H2O2 (Sigma) or with alkaline phosphatase-based 1 step-NBT/BCIP substrate (ImmunoPure ABC kit; Pierce). To inhibit endogenous alkaline phosphatase activity, ImmunoPure Phosphatase Suppressor (levamisole) was added to NBT/BCIP substrate in 1:100 dilution.
Results
Expression of OP-1 mRNA
We investigated the expression of OP-1 mRNA in human articular cartilage using comparative RT-PCR and in situ hybridization. By RT-PCR mRNA bands for OP-1 in newborn, normal adult, and OA cartilages were detected (Figures 1A and 1B). PCR bands with specific OP-1 primers revealed the same molecular size products (313 bp) as expected (Figure 1A). When densities of OP-1 mRNA bands were normalized to densities of the GAPDH bands, the relative levels of OP-1 expression in newborn and normal adult cartilage (Figure 1A, Lanes 2-7) were almost the same. However, in OA cartilage (Figure 1A, Lanes 8-10), mRNA expression of OP-1 appeared to be upregulated by at least twofold (p<0.01; Figure 1B). Actual values of the PCR bands are represented in Table 1. Sequence analyses of the final PCR products from one normal adult cartilage and one OA cartilage showed 100% homology with human OP-1 mRNA (accession number X51801; Özkaynak et al. 1990).
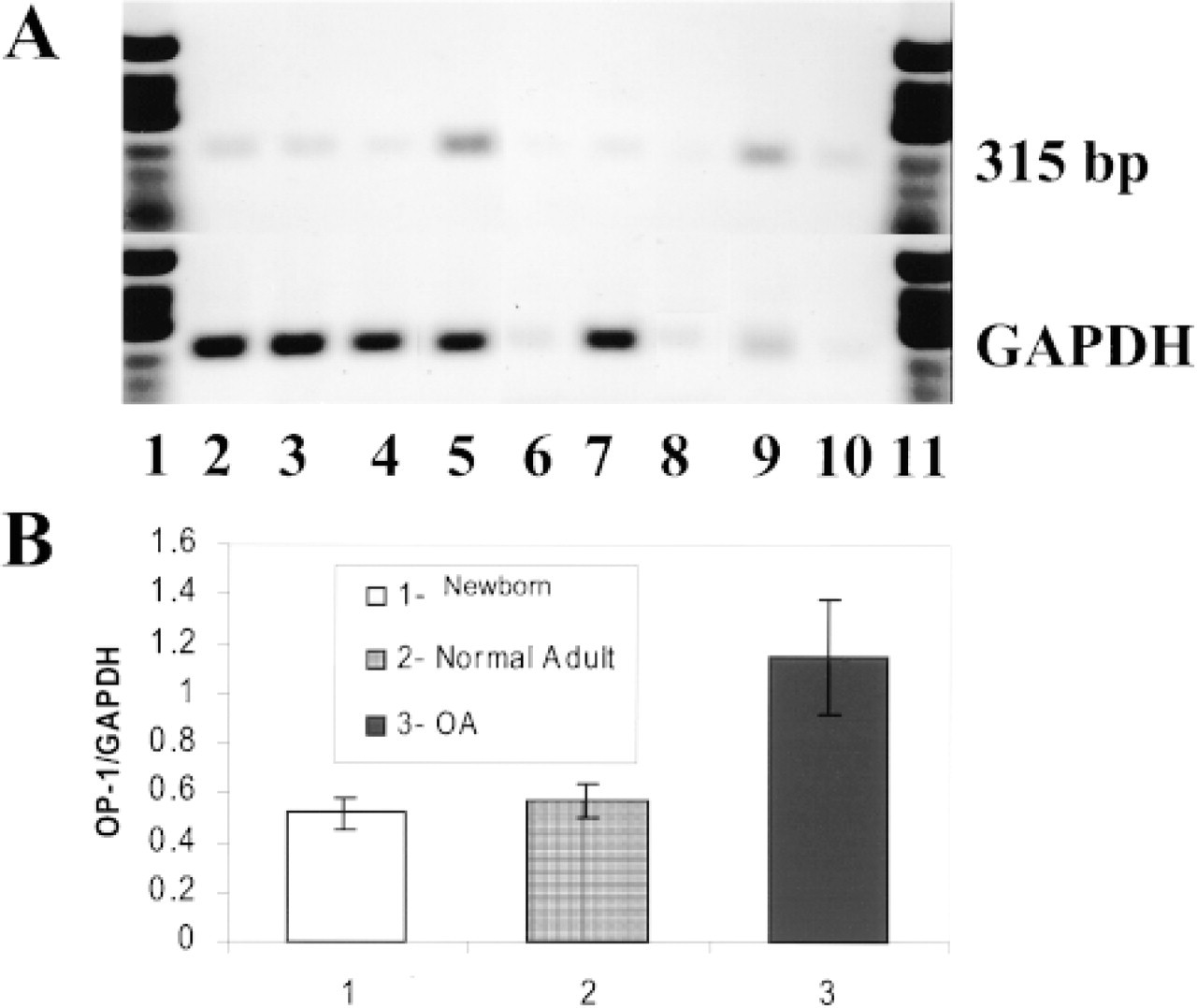
RT-PCR analyses of mRNAs of normal and OA cartilage samples for the presence of OP-1 and GAPDH. (
Densities of the PCR-derived bands for OP-1 and GAPDH mRNA products a
OA, osteoarthritic cartilage.
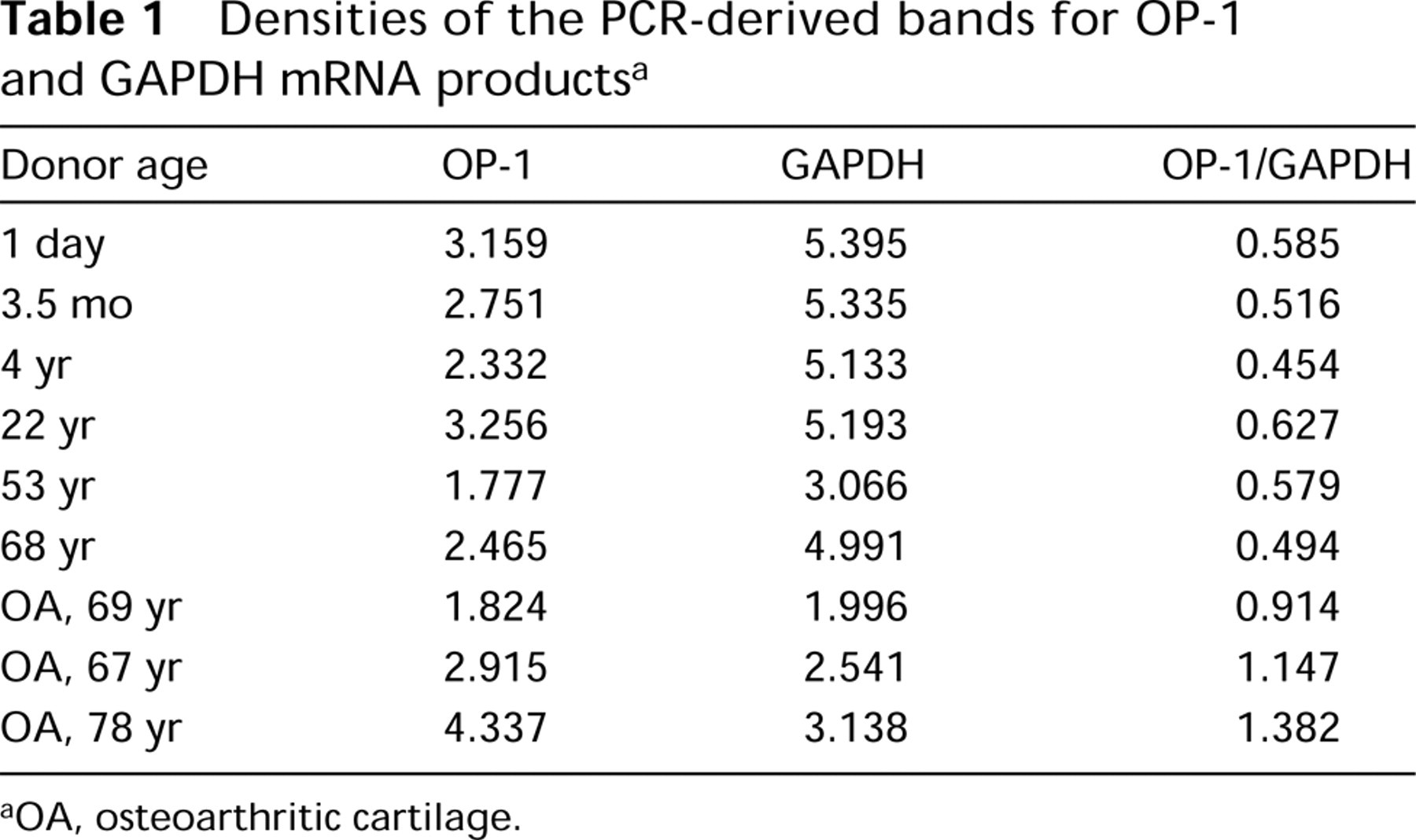
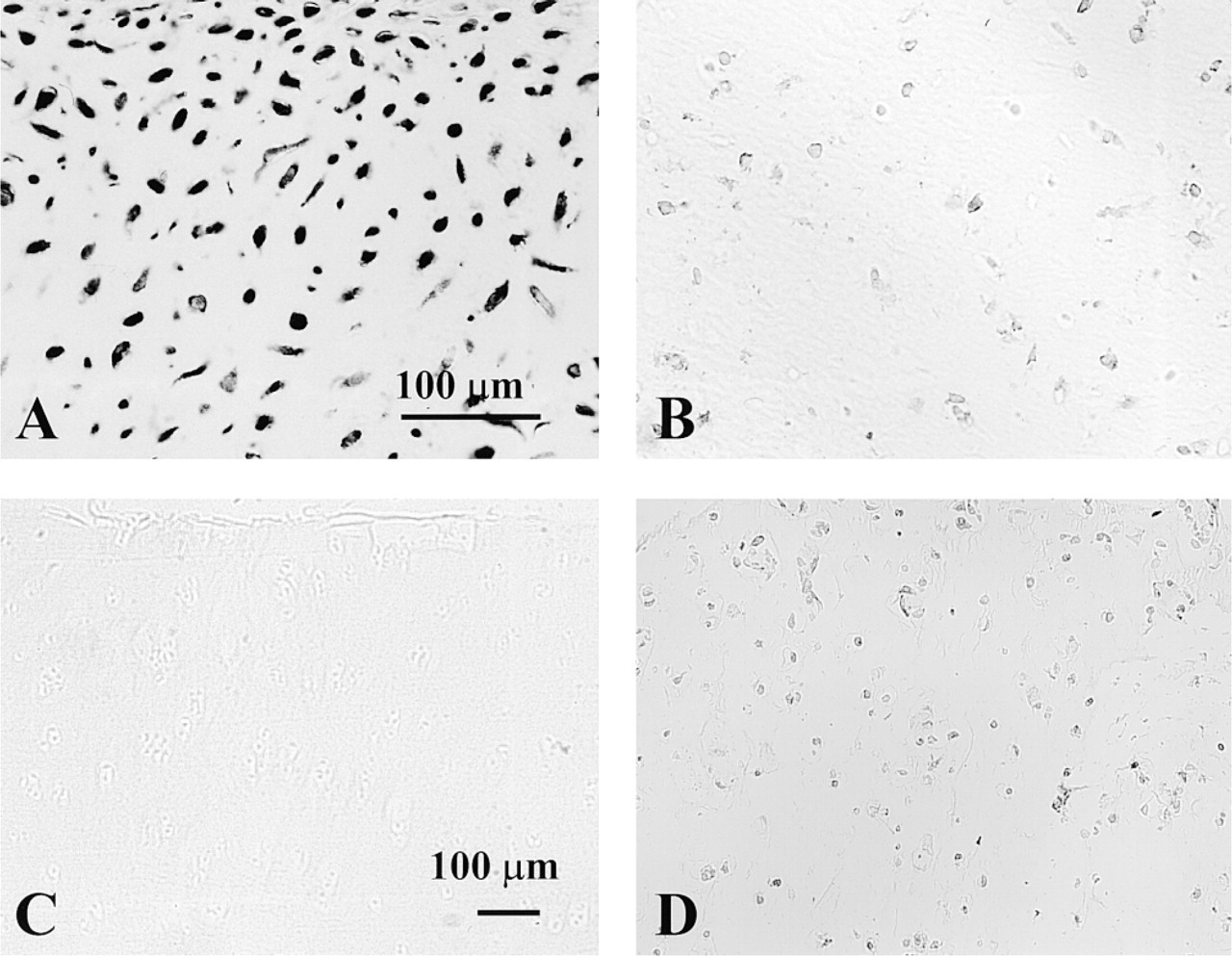
Sequencing of the PCR products served as a positive control for in situ hybridization because the same PCR primers were applied for both methods. Sense primers and the inhibition control were used as negative controls. Sense oligonucleotide probes (or upstream primers) revealed no specific binding to the chondrocytes (not shown). Addition of unlabeled probe to the radiolabeled oligonucleotide probe at a ratio of 1:1 was able to inhibit specific binding of the probe to the mRNA within the chondrocytes (Figures 2A and 2B), thus demonstrating the high specificity of the probes.
As a first step with in situ hybridization, the mRNA expression of OP-1 was tested in isolated chondrocytes cultured in alginate beads. These cells showed strong expression of OP-1 mRNA (Figure 3A). In situ hybridization on cartilage sections from different donors and patients verified that all types of analyzed cartilage were positive for OP-1 message. In newborn cartilage, expression was evident from the upper epiphyses to the hypertrophic area (Figure 3B). In normal adult cartilage, OP-1 expression was primarily found in the superficial and middle layers of the cartilage (Figure 3C), whereas in the deep layer levels of expression were very low or below the detection limit. In OA cartilage, OP-1 message was detected in all cartilage layers, with the strongest expression in chondrocyte clusters (Figure 3D).
Expression of OP-1 Protein
To determine if OP-1 protein can be detected in human articular cartilage, extracts from normal adult and OA cartilages were analyzed by Western blotting. Human recombinant mature and pro-OP-1 were used as positive controls (Figures 4A and 4B, Lanes 1 and 2). As expected, antibody to pro-domain (R2854) did not recognize mature recombinant OP-1 (Figure 4A, Lane 1) but did recognize the pro-form of the molecule (Figure 4A, Lane 2). Antibodies to mature domain (12G3) detected both mature OP-1 (Figure 4B, Lane 1) and pro-OP-1 (Figure 4B, Lane 2). In extracts from all cartilages, anti-mature and anti-pro-domain antibodies recognized immunoreactive bands. Under nonreduced conditions, pro-domain antibody (Figure 4A) recognized three major bands in the pro-OP-1 control at about 36-40 kD, 70 kD, and 110 kD. Two of these immunoreactive bands (about 36-40 kD and 70 kD) were also present in the cartilage extracts. In addition, two other major bands at 50 kD and above 220 kD were identified. The band at 70 kD may correspond to a mature dimer associated with one pro-domain, called a hemidimer, and bands between 45-55 kD could represent the monomer of the pro- and mature domains. According to Jones et al. (1994), the band at 110 kD corresponds to a soluble OP-1 complex with one mature OP-1 dimer (35 kD) noncovalently associated with two pro-domains (40 kD each).
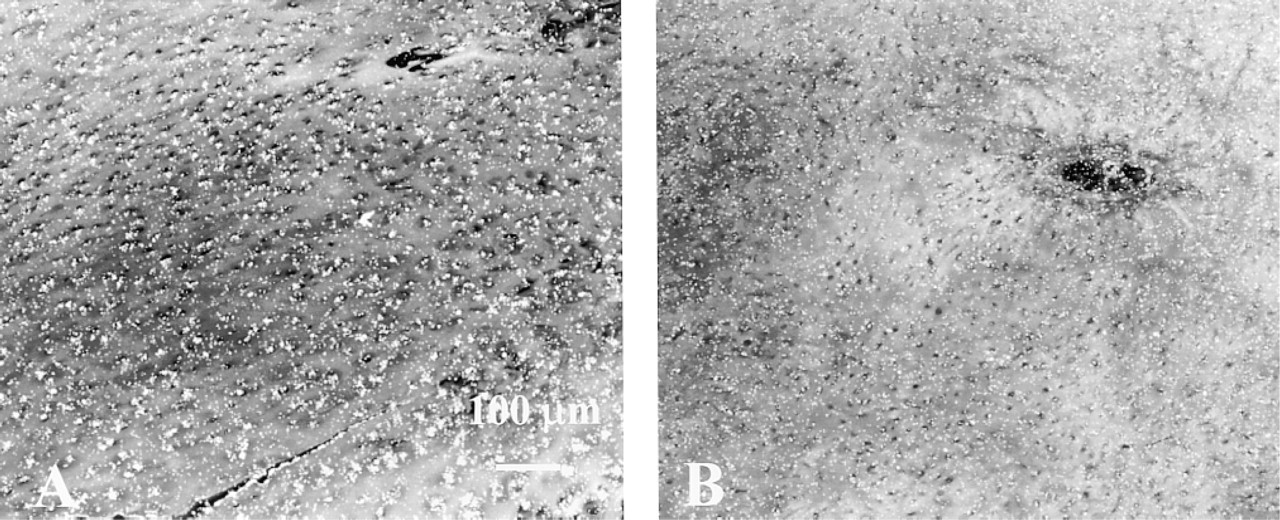
Darkfield photomicrographs of human cartilage hybridized to the OP-1 oligonucleotide probe. (
Darkfield photomicrographs of human cartilage hybridized to the OP-1 oligonucleotide probe. (
Immunoblot probed with antibody to mature domain (Figure 4B) also showed strong immune bands in both controls and in the cartilage extracts. In controls, the same major band at about 36-40 kD was recognized (Figure 4B, Lanes 1 and 2). Furthermore, two other bands at about 50 and 84 kD were found in the mature OP-1 control. This antibody revealed specific OP-1 bands in the pro-OP-1 preparation at the same molecular weight (about 70 and 110 kD) as anti-prodomain antibody. Extracts from human cartilages showed the strongest immunoreactive band at about 70 kD with antibody to mature domain. Other bands that could be identified on the Western blot with the same antibody are 36-40 kD, above 220 kD, and a few minor bands between 45 and 55 kD (Figure 4B). It is important to note that in extracts from OA cartilages an additional low molecular weight band at about 17 kD was detected (Figure 4B, Lane 5). This band could also be seen in some of the donor tissue with signs of asymptomatic degenerative morphological changes (Figure 4B, Lane 4). A band at about 220 kD probably indicates high molecular weight aggregates of OP-1 with matrix molecules, such as collagens or fibronectin, because high affinity of OP-1 binding to different collagens was previously reported (Vukicevic et al. 1994). The 36-kD band most likely represents the mature OP-1 dimer alone or associated with cleaved pro-domains, and the band at 17 kD presumably represents the mature OP-1 monomer. The 17-kD band may also be a fragment or component of the entire OP-1 molecule. However, we assume that this band represents only the OP-1 monomer because its molecular weight is closer to the reported one and this band is not recognized by anti-pro-domain antibody (Sampath et al. 1992; Jones et al. 1994).
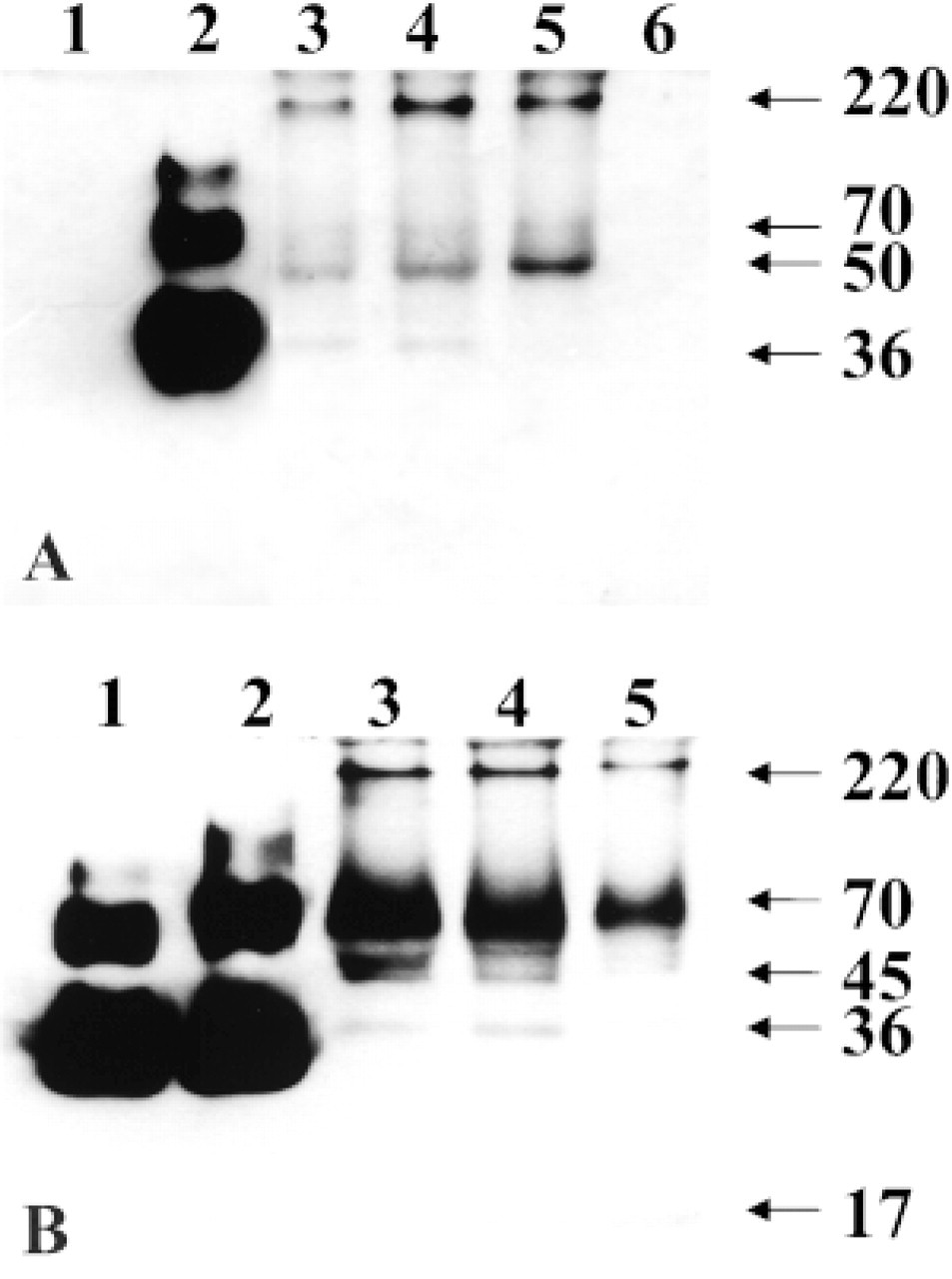
Representative Western blots of knee cartilage extracts with antiserum against OP-1 pro-domain (
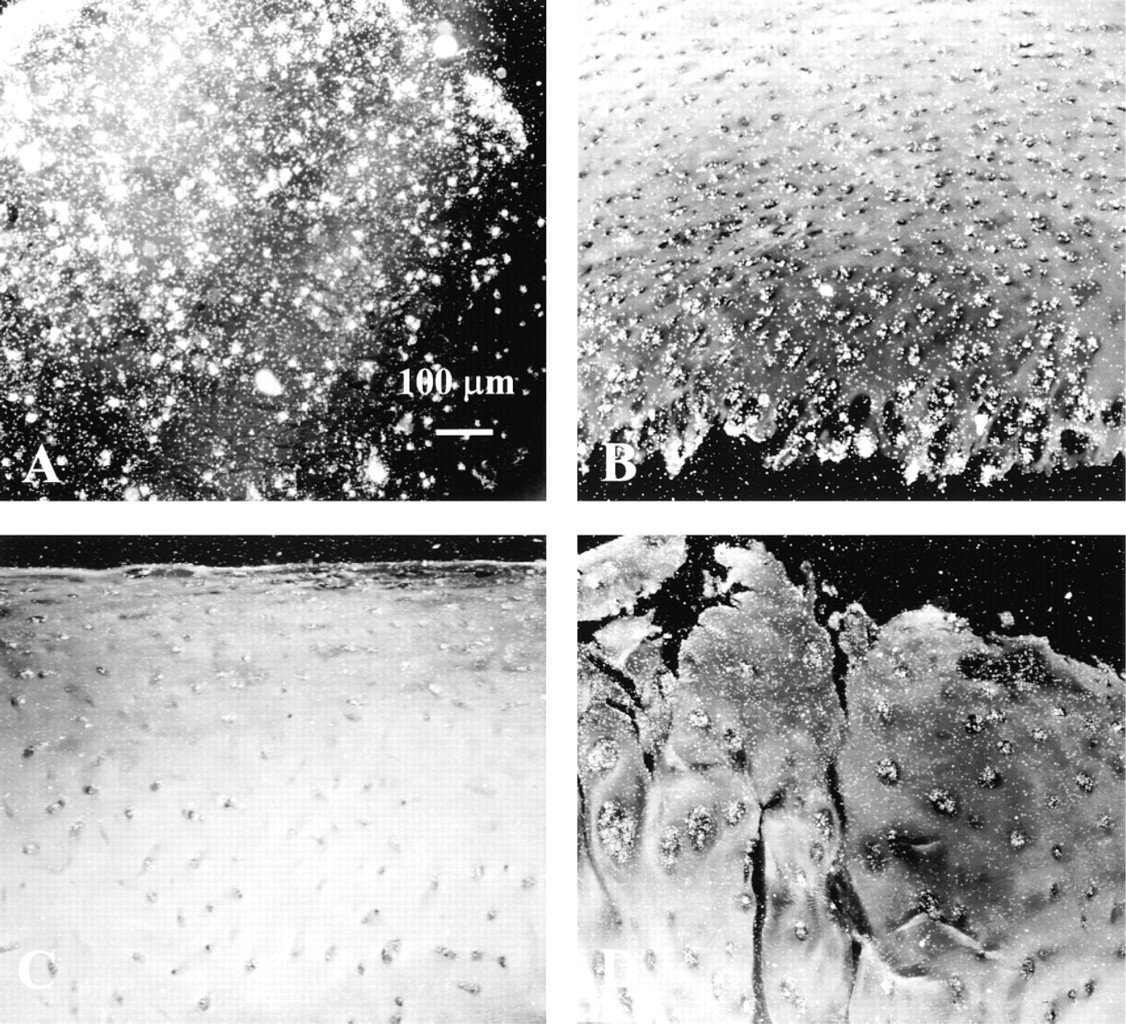
Immunohistochemical studies were performed on newborn chondrocytes cultured in alginate beads and on cartilage sections. The specificity of antibodies was demonstrated with an inhibition control when primary antibodies were preabsorbed with human recombinant OP-1. The addition of recombinant mature OP-1 decreased the intensity of anti-mature OP-1 staining (Figures 5A and 5B). The same type of control was used to demonstrate the specificity of pro-OP-1 immunostaining with antibody to the pro-domain (not shown). When immunostaining was carried out in the absence of primary antibody (negative control), no positive staining of the cartilage section was visible (Figure 5C). Chondrocytes from newborn donors cultured in alginate beads showed positive staining with anti-mature OP-1 antibody (Figure 5D). The positive immunostaining of human cartilage with antibodies to mature and pro-domains is shown in Fig. 6.
The most striking differences in the distribution of pro- and mature OP-1 were found by immunostaining of cartilage sections. Furthermore, there were distinct staining patterns among newborn, normal adult, and OA cartilage. In newborn growth plate cartilage, very strong intracellular staining with antibodies to mature domain was evident throughout the tissue in the epiphysis, proliferative, and hypertrophic regions (Figures 6A and 6B). The extracellular matrix showed no staining. On the other hand, both the perichondrial matrix and the cells entrapped within it were intensively stained (Figure 6A). When the same newborn cartilage was introduced to the antibody to pro-domain, the strongest staining was detected in the lower hypertrophic zone of growth plate cartilage (Figures 6C and 6D). In the resting and proliferating zones, the intensity of pro-OP-1 staining was lower than in the hypertrophic region. Pro-OP-1 was mostly localized intracellularly and was almost undetectable within matrix. In normal adult cartilage, mature OP-1 was found predominantly in the vicinity of the chondrocytes (within the cells and in the surrounding pericellular matrix) primarily in the superficial and upper middle layers of the tissue (Figure 6E). Some staining was evident in the extracellular matrix. Deep-layer chondrocytes showed very low or practically no mature OP-1 (Figure 6F). In contrast to the distribution of mature OP-1, the majority of pro-OP-1 was localized in the deep layer of tissue, with decreased amounts of pro-OP-1 in the middle and superficial zones of the cartilage (Figures 6G and 6H). The same trend in layer distribution of mature and pro-forms of OP-1 was described for OA cartilage. In the chondrocytes from the upper (superficial) part of OA cartilage, little anti-mature OP-1 staining appeared to be cell-associated, with low staining of lacunae walls. In addition, some staining was evident in the territorial and interterritorial matrix of the upper and middle layers of OA cartilage (Figure 6I). The deep layer of OA cartilage, as was seen in normal adult cartilage, showed very little or no anti-mature OP-1 staining (Figure 6J). The overall level of anti-mature OP-1 staining in OA cartilage appeared to be less intense than in newborn and normal adult cartilage. However, when the same OA cartilage was stained with the anti-pro-domain antibody, deep-layer chondrocytes showed very strong immunoreactivity. The intensity of staining was gradually decreased from the deep zone to the upper zone (Figures 6K and 6L). Moreover, pro-OP-1 staining was detected in the territorial matrix of the deep and middle layers, and granular staining of pro-OP-1 was observed in the middle and upper (superficial) layers of OA cartilage.
Immunostaining of cartilage and alginate bead sections with anti-OP-1 antibody (12G3). (
Discussion
By in situ hybridization, RT-PCR, Western blotting, and immunohistochemistry, we have demonstrated for the first time that human adult articular chondrocytes at different ages and different histopathogical stages synthesize OP-1, a bone morphogenetic protein that has been reported to be involved in a variety of physiological and pathophysiological processes (Sam-path and Rueger 1994; Cook and Rueger 1996; Urist 1997; Reddi 1998).
The majority of OP-1 studies have focused on the exogenous application of OP-1 and the resulting induction of anabolic response in a variety of connective tissues including bone, cartilage, and disks (Chen et al. 1993; Dieudonné et al. 1994; Flechtenmacher et al. 1996; Huch et al. 1997; Koepp et al. 1999; Masuda et al. 1999; Takegami et al. 1999). However, very little was known about the endogenous expression of OP-1 by connective tissue cells, and particularly by adult articular chondrocytes. The data from our present study and from previously published work (Özkaynak et al. 1990; Vukicevic et al. 1994; Helder et al. 1998; Weber et al. 1998) prompts us to hypothesize that OP-1 may be an endogenous growth factor that plays a role not only in the development and repair of different tissues and organs but also in maintaining adult tissue homeostasis under normal and pathological conditions. To our knowledge, it is presently the only member of the BMP family, along with cartilage-derived morphogenetic proteins, found to be expressed by adult articular chondrocytes.
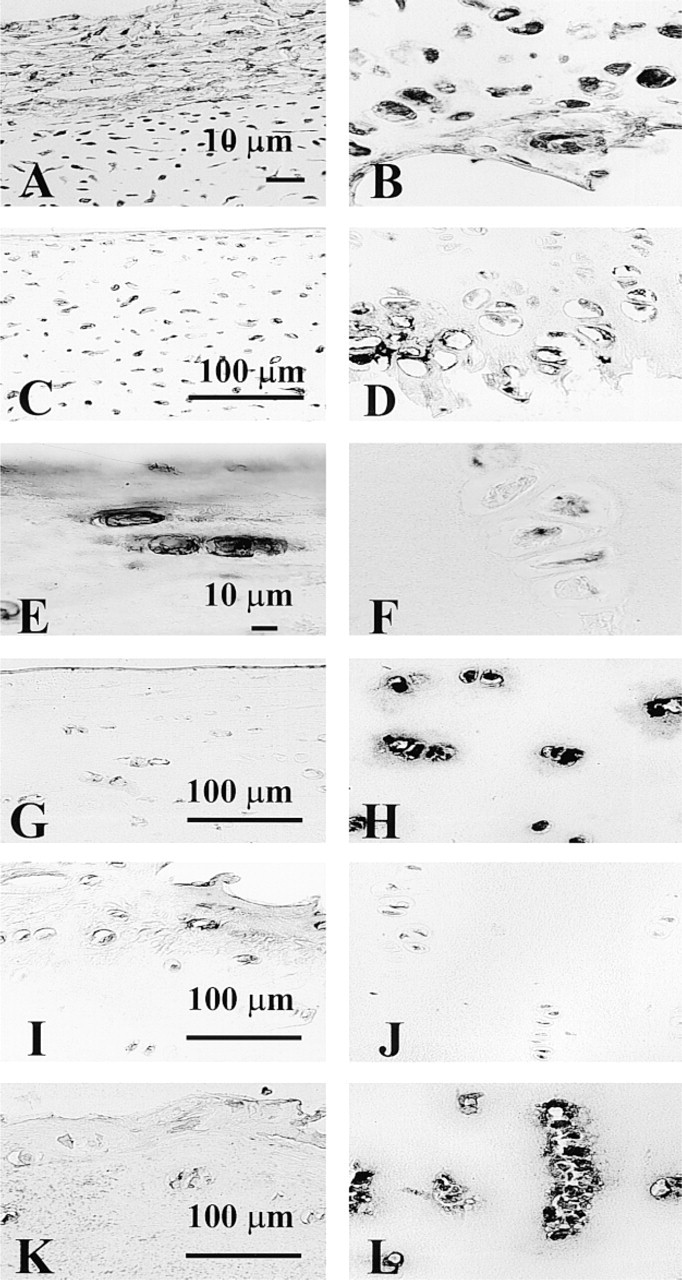
Immunostaining of cartilage sections with anti-OP-1 antibody 12G3 (
Interestingly, we detected two forms of the OP-1 molecule—the inactive pro-form and the active mature form—simultaneously present in human adult cartilage. The distinction between the two forms of the protein was possible because of the availability of two types of antibodies. The polyclonal antibody (R2854) was raised in rabbits against the entire pro-domain of the OP-1 sequence (amino acids 29 to 292; Jones et al. 1994; Helder et al. 1998) and recognizes only the pro-OP-1. Two mouse monoclonal antibodies (12G3 and 1B12; Vukicevic et al. 1994) were raised against the entire mature domain (amino acids 293-431) and recognize both forms of OP-1, pro-OP-1 and mature OP-1. 1B12 and 12G3 antibodies react only with the oxidized dimeric OP-1 (Vukicevic et al. 1994). Therefore, for the current studies, only nonreduced conditions were applied. Immune bands seen by both antibodies (R2854 or 12G3) presumably represent unprocessed or partially processed OP-1 in which the pro-domain units remain attached to the mature OP-1 dimers. Additional bands identified by Western blotting with 12G3 antibody may represent only the monomers or fragments of the mature form of OP-1. The heterogeneity of bands exhibited on both Western blots could be explained by the complexity of the multistep process of OP-1 activation described by Jones et al. (1994) for CHO cells. Nevertheless, the most important observation was that in normal and OA cartilage extracts all forms of OP-1, monomeric, dimeric and, possibly, oligomeric, were detected. Moreover, oligomerization could have occurred between the same domains (either pro- or mature), between pro- and mature domains together, or among the entire molecule.
Very intriguing data obtained by immunohistochemistry was the inverted pattern in the localization of mature and pro-OP-1. Here it is important to mention that adult articular cartilage is a heterogeneous tissue in which the cells and matrix are arranged in layers (Hunziker 1992) that have been designated as superficial (tangential), middle (transitional), and deep (radial). Mature OP-1 was highly visible in the superficial layer of cartilage, whereas pro-OP-1 was primarily detected in the deep zone of tissue. For some reason, anti-mature antibodies 12G3 and 1B12 did not recognize the pro-molecule epitope when it is within the tissue, even though these antibodies were raised against the entire mature molecule and were expected to recognize both forms of OP-1. The possible explanation could be that the pro-molecule has a protein conformation that is inaccessible to the anti-mature OP-1 antibodies (personal communications, Dr. Sampath). However, they do see the pro-molecule when it is extracted. Furthermore, anti-mature and anti-pro-OP-1 antibodies could have different strengths, different specificities, and different affinities. As previously reported (Helder et al. 1998), anti-mature antibodies reacted much more weakly (both intra- and extracel-lularly) than anti-pro-OP-1 antibody. To confirm that the real biological phenomenon was observed by immunohistochemistry and that the pro-OP-1 epitope is not masked in the tissue, cartilage sections were pretreated with hyaluronidase (unpublished data). However, the patterns of staining with both types of antibodies did not differ from those discussed above for the sections pretreated with a chondroitinase cocktail. Nevertheless, the observed localization of pro- and mature forms of OP-1 suggests that the processing of pro-OP-1 into mature OP-1 occurs primarily in the superficial chondrocytes. When the superficial layer is disrupted or absent, as it is in OA cartilage, the chondrocytes from the middle and deep cartilage zones apparently do not replenish the active OP-1 by further processing of pro-OP-1. In this case, the majority of OP-1 is probably still stored intracellularly as an inactive protein. An understanding of the mechanisms of OP-1 processing in human cartilage may yield new insights into the problem of the protection of cartilage from degenerative processes or even in the induction of cartilage repair.
Unlike the protein data, expression of OP-1 mRNA was upregulated in diseased cartilage compared to normal newborn and adult tissues. Moreover, in OA cartilage, chondrocytes of all layers demonstrated positive expression of OP-1, whereas in normal adult cartilage the superficial chondrocytes were primarily positive for OP-1 message. Furthermore, OP-1 expression was the strongest in cell clusters (areas of OA cartilage with some cell division). These data were supported by the results of RT-PCR showing that OP-1 mRNA expression was more than twofold higher in OA cartilage than in normal newborn and adult cartilage.
The observed results indicate, first, that the OP-1 gene is inducible in the deep-layer chondrocytes when the superficial layer is destroyed and, second, that in diseased cartilage chondrocytes attempt to withstand catabolic events by upregulation of mRNA for OP-1. However, this upregulation is limited to the OP-1 message and does not appear to take place at the level of protein translation or activation.
Collectively, the data presented here suggest a role for OP-1 in cartilage homeostasis/protection and/or repair. These data further imply that the regulation of the activity of OP-1 at the level of protein may occur through the control of the enzymatic cleavage of pro-OP-1 into the mature form, which could be a key mechanism governing the function of OP-1 in cartilage pathophysiology.
Footnotes
Acknowledgments
Supported by NIH grant 2P50-AR-39239-11.
We thank Dr A. Hari Reddi for valuable suggestions and discussions, and Drs Klaus Huch, Wolfgang Eger, and Arcady Margulis for providing cartilage samples. We would especially like to thank Dr Carol Muehleman for helpful comments on the manuscript. Collaboration with Allan Valdellon, MD, of the Regional Organ Bank of Illinois and his staff, as well as the surgeons of the Department of Orthopedic Surgery at Rush-Presbyterian-St Luke's Medical Center, is gratefully acknowledged.