
Abstract
The ovarian steroids estrogen and progesterone are important in directing the normal growth and development of the mouse mammary gland. Previously, we have demonstrated that the majority of proliferating mammary epithelial cells do not express estrogen receptor-α (ERα). In this study we examined the relationship between progesterone receptor (PR) expression and proliferation in mammary epithelial cells using simultaneous immunohistochemistry for progesterone receptor (PR) and tritiated thymidine [3 H]-Tdr) autoradiography. Results showed that the majority (>80%) of mammary epithelial cells labeled with [3 H]-Tdr were PR-positive in the terminal end buds (TEBs) of pubertal mice and the ducts of pubertal and adult mice. Whereas the majority of mammary epithelial cells were also PR-positive, the basal cell population, which comprises the minority of mammary epithelial cells in the mammary ducts, was predominantly PR-negative. Nevertheless, the PR-positive phenotype remained the major proliferating cell type in the basal population. These findings suggest that the progesterone signaling pathway is involved in the proliferation of basal cell populations, potentially directing formation of tertiary side branching during pubertal development and alveolar bud formation in adult glands. A proportion of the basal cells exhibited weak expression of ERβ suggesting that the role of ERβ in mediating normal estrogen-induced responses should be further studied.
T
In a previous study (Zeps et al. 1998), we demonstrated that although estrogen has an established role in stimulating mammary epithelial proliferation, the majority of epithelial cells do not express ERα in both pubertal and adult mice. Progesterone has also been ascribed a significant role in stimulating the proliferation and differentiation of mammary epithelial cells during the normal development and function of the female mouse mammary gland (Nandi 1958; Bresciani 1971; Haslam 1988; Shyamala et al. 1997), and it is feasible that it may be in part responsible for directing proliferation in these ERα-negative cells. Several lines of evidence indicate that the actions of progesterone are directly mediated by the progesterone receptor (PR), a member of the steroid receptor superfamily of nuclear transcription factors (Graham and Clarke 1997; Shyamala et al. 1997). Although PRs are present in a large proportion of mammary epithelial cells, particularly in the highly proliferative end buds of the pubertal mouse mammary gland (Silberstein et al. 1996; Shyamala et al. 1997), the functional significance of progesterone and PR expression in mammary gland development is not known. However, evidence from studies using transgenic mice with a disrupted PR (PRKO) (Lydon et al. 1995) or mice that constitutively overexpress the isoform of PR, PR-A (PR-A-transgenic) (Shyamala et al. 1998) indicates that PR expression is important in both the normal growth of mammary ducts and in the side branching that arises from them during puberty. In adult mice, it has been proposed that progesterone is the principal mammary mitogen during estrus and pregnancy, its effects mediated by estrogen-induced PR in the mammary epithelial cells (Shyamala 1997; Fendrick et al. 1998). The significantly impaired lobulo-alveolar development in PRKO mice supports the importance of PR in mammogenesis (Lydon et al. 1995).
PR expression has been reported in the highly mitotic cap cell population of the TEBs during puberty and in the majority of the luminal cells of mammary ducts during lobulo-alveolar development in pregnant adult mice (Silberstein et al. 1996). Although it is possible that progesterone directly stimulates proliferation of these PR-positive epithelial cells, the precise relationship between PR expression and proliferation is not known. In this study we have investigated PR expression in proliferating mammary epithelial cells during puberty and during pro-estrus (quiescent stage) and estrus (proliferative stage) in virgin adult mice. Using dual immunohistochemistry for PR and [3 H]-Tdr autoradiography, we observed that whereas the majority of proliferating mammary epithelial cells were PR-positive, PR expression appeared to be associated specifically with proliferation only in the basal cell population in the mammary ducts of pubertal and adult mice. Furthermore, although basal cells were uniformly ERα-negative, ERβ was detected in low levels in this cell population, introducing the possibility that estrogen-induced PR expression or proliferation might be mediated through this receptor.
Materials and Methods
Animals and Tissues
This study was performed in compliance with Australian State and Federal animal protection laws under a permit issued by the Animal Experimentation and Ethics Committee of the University of Western Australia. Mammary glands from virgin female Balb/c mice (Animal Resource Centre; Perth, Western Australia) 5 weeks of age (pubertal) and 10-12 weeks of age (adult) were used in these studies. To investigate the role of ovarian hormones during mammary gland growth in adult mice, animals between 10 and 12 weeks of age were staged for estrus by vaginal smear over several days (Zeps et al. 1996). Mice identified as being in pro-estrus or estrus were used in this study, and stage in estrus was confirmed histologically after sacrifice. Forty minutes before sacrifice, all mice received a single
Immunohistochemistry
Sections were dewaxed in xylene, rehydrated, and washed in Tris-buffered saline (TBS, pH 7.4). Slides were immersed in 5 mM ethylenebis (oxyethylenenitrilo) tetraacetic acid (EGTA), pH 8.0 (Sigma), and boiled for 15 min in a pressure cooker in a microwave. After antigen unmasking, sections were incubated for 20 min each in avidin and biotin blocking solutions (DAKO Biotin blocking kit; Carpinteria, CA). They were then incubated in 1.5% H2O2 (BDH, Merck; Victoria, Australia) in methanol (BDH) for 10 min, then blocked with 20% normal horse serum (NHS) in TBS for 30 min.
Sections were incubated overnight in one of the following antisera: rabbit polyclonal anti-PR (2 μg/ml in blocking solution; SC-538, Santa Cruz Biotechnology, Santa Cruz, CA), rabbit polyclonal anti-ERβ (5 μg/ml in blocking solution; Affinity Bioreagents, Golden, CO), rabbit polyclonal anti-ERα (1 μg/ml in blocking solution; SC-542, Santa Cruz Biotechnology). Sections were incubated in swine anti-rabbit biotinylated secondary antibody (Dako) diluted 1:200 in blocking solution, followed by streptavidin-peroxidase (Silenius; Melbourne, Australia) diluted 1:200 in TBS. All incubations were carried out at RT for 50 min. Between incubations, sections were washed three times for 5 min in PBS containing 0.2% polyoxyethylene sorbitan monolaurate (Tween-20). Immunoreactivity was detected by incubation of sections in metal-enhanced 3,3′-diaminobenzidine tetrahydrochloride (DAB; Pierce, Perth, Australia) for 1 min. The sections were washed in dH2O for 10 min, then transferred to 70% ethanol overnight before being dipped in Kodak NTB-2 autoradiographic emulsion as described previously (Zeps et al. 1996). For negative control sections, either primary antibody was replaced with 1% BSA in TBS or control peptide (Santa Cruz Biotechnology) was used in preincubation with primary antisera in 10-fold excess. No false-positive staining was observed in controls.
Data Collection and Analysis
At least 15 randomly chosen TEBs and/or ducts were counted per animal in serial sections of the mammary glands, representing >2000 epithelial cells for each mouse, using an Olympus BX-40 light microscope at × 1000 (oil immersion). Cells were scored according to location, receptor expression (+ or -), and [3 H]-Tdr labeling [background grain counts were low in all sections and nuclei with three or more silver grains above them were counted as positive (Zeps et al. 1996)]. Basal cells were discriminated on the basis of their elongated appearance and their location within and lying parallel to the basement membrane (Chepko and Smith 1997). Luminal cells contained large, pale-staining round nuclei, condensed small nuclei, or a “gourd”-like appearance as previously described (Smith and Medina 1988; Chepko and Smith 1997). Luminal cells lay within and perpendicular to the basement membrane. Data are summarized as the average of the means of three to six mice ± SEM. Data were compared using Student's t-test.
Results
Progesterone Receptor Expression in Pubertal Mice (5 Weeks)
PR-positive cells were recognizable by a well-defined brown nuclear stain (Figure 1A) and were present in mammary epithelial cells during all stages of pubertal development. The PR and proliferation status in the terminal end buds (TEBs) and the ducts of 5-week-old mice are presented separately below. Similarly, ERα immunoreactivity was detected as a nuclear stain in a proportion of mammary epithelial cells, as previously described (Zeps et al. 1998).
Terminal End Buds
PR immunoreactivity was observed in all TEBs and was heterogeneously distributed in both the cap and body cells (Figure 1A). Of the 63 TEBs scored (representing 7435 cells), the majority of both the outer cap cell layer (90 ± 2.9%) and the inner population of body cells (83 ± 0.5%) were PR-positive (Figures 2A and 2B, respectively). All cap cells were ERα-negative (Figure 2A), and in the body of the TEBs 62 ± 0.2% of epithelial cells were ERα-positive (Figure 2B).
When cells were scored for S-phase, 95 ± 1.6% of the [3 H]-Tdr-positive cap cells were PR-positive and all of these cells were ERα-negative (Figure 2C). In the body of TEBs, 83 ± 0.5% of proliferating cells were PR-positive compared to 63 ± 1.3% ERα-positive proliferating cells (Figure 2D). The percentage of body cells labeled with [3 H]-Tdr (thymidine labeling index, TLI) was 13 ± 0.7%, significantly less (p=0.03) than that of the cap cells, which had a TLI of 24 ± 2.9%. Nevertheless, despite the relatively lower TLI of the body cells, because they constitute the majority of cells in TEBs (76 ± 1.1%) they also comprised the majority of the proliferative compartment (64 ± 0.7%).
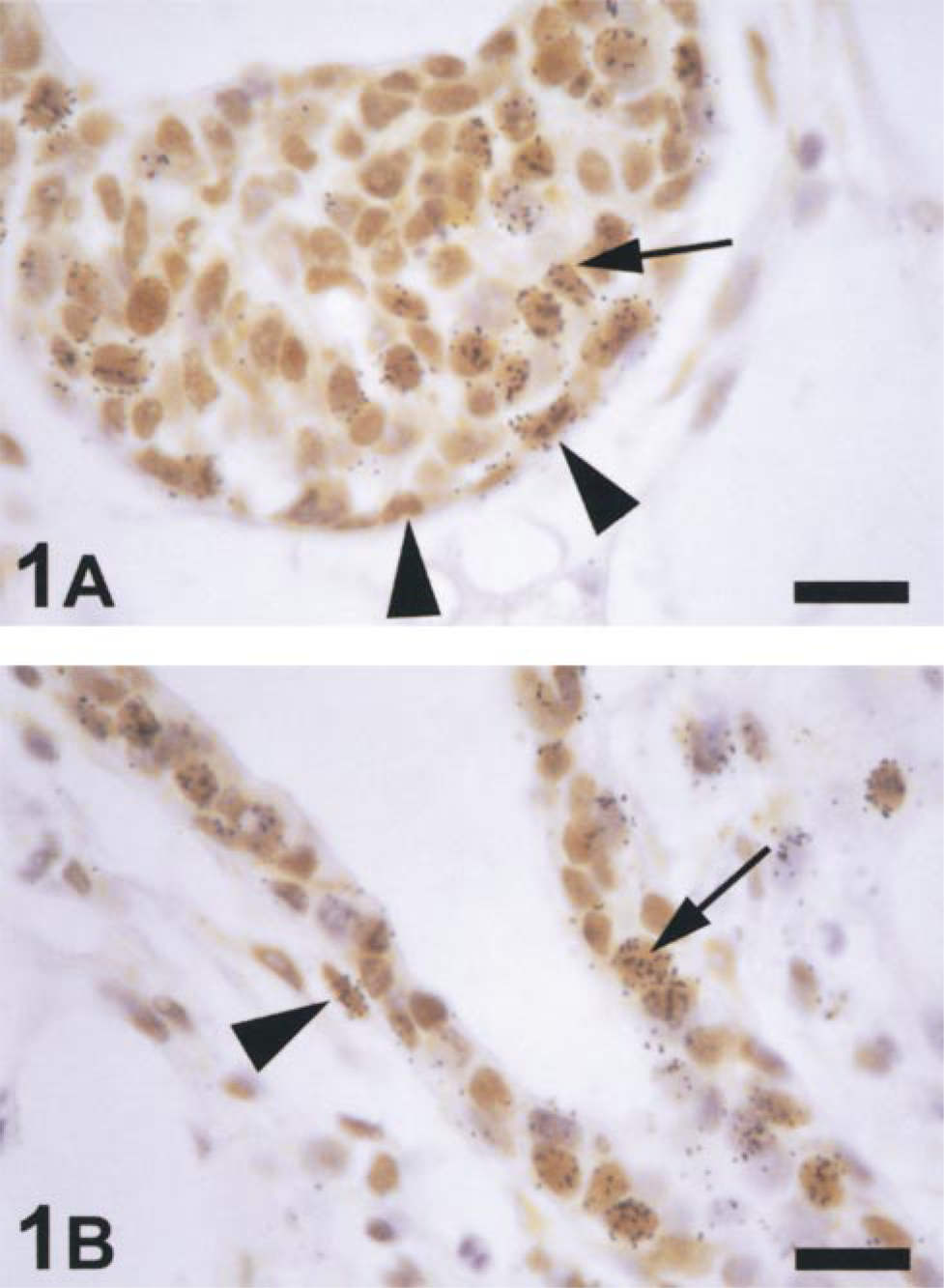
Light micrographs of immunohistochemical staining for PR and [3 H]-Tdr autoradiography in the mammary gland from a 5-week-old mouse sacrificed 40 min after injection with [3 H]-Tdr. The cap ▴ and body ↑ cells in TEBs (A) had strong nuclear staining for PR and a proportion of these were labeled with [3 H]-Tdr. PR-positive basal ▴ and luminal ↑ cells labeled with [3 H]-Tdr were present in the ducts (B). Bars = 16 μm.
Ducts
PR was expressed in both the basal and the luminal cells of mammary ducts in 5-week-old mice (Figure 1B). A total of 55 ducts were examined representing 4302 cells. PR was expressed in a minority (43 ± 1.9%) of basal cells (Figure 3A), all of which were ERα-negative. In the luminal cell population of the ducts, 86 ± 4.1% of cells were PR-positive (Figure 3B), and only 55 ± 0.4% of luminal cells were ERα-positive (Figure 3B). The proportion of ERα-positive luminal cells in the ducts was significantly less than the proportion of ERα-positive body cells in the TEBs (p=0.004).
The majority (78 ± 8.6%) of proliferating basal cells in the ducts were PR-positive, and all were ERα-negative (Figure 3C). Almost all proliferating luminal cells (94 ± 2.0%) were PR-positive (Figure 3D); only 54 ± 5.0% of luminal cells labeled with [3 H]-Tdr expressed ERα. Although the relative TLI in the luminal cell population (TLI = 7 ± 0.4) appeared to be less than that in the basal cell population (TLI = 10 ± 2.6%) this difference was not statistically significant (p=0.19). However, like the body cells of the TEBs, the inner luminal cells comprise the largest proportion of the duct epithelium (80 ± 1.9%) and hence contain the majority of duct proliferating cells (73 ± 4.4%).

Histograms comparing the mean proportions of cells ▪ positive and () negative for PR and ERα in all cap (A) and body (B) cells and in only those labeled with [3 H]-Tdr (proliferating) in the cap (C) and body cell (D) populations in the TEBs of mammary glands from 5-week-old mice.
Progesterone Receptor Expression in Adult Mice (10-12 Weeks Old)
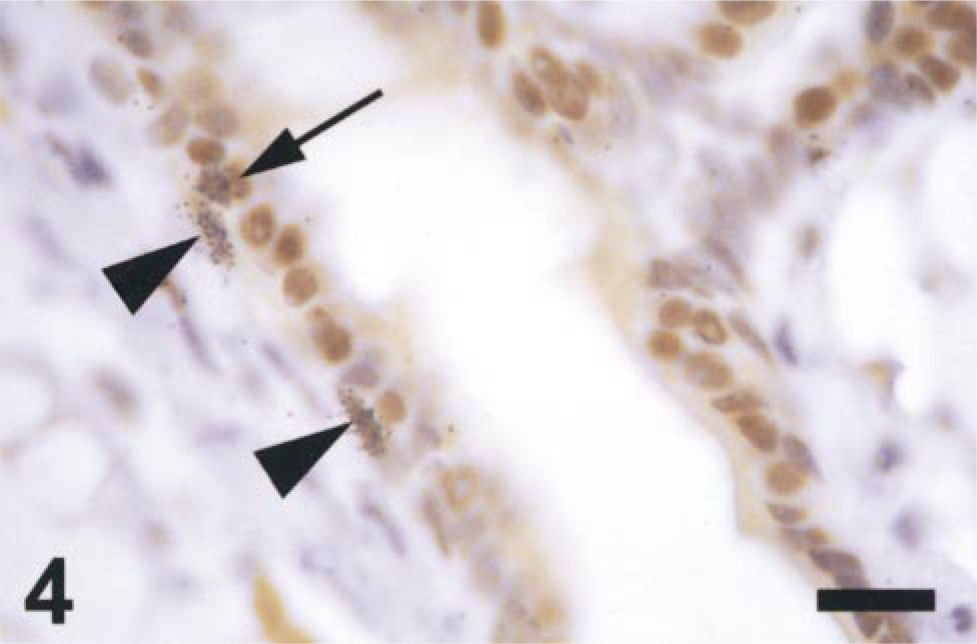
Staining of adult mammary tissues revealed that both basal and luminal cells expressed detectable levels of PR protein (Figure 4). In these ducts, as in those from pubertal animals, the majority of basal cells do not express PR either during pro-estrus, when 25 ± 1.4% of cells were PR-positive, or during estrus, when 32 ± 8.3% of basal cells were PR-positive (Figures 5A and 5B). The majority of luminal cells have detectable levels of PR during both pro-estrus (83 ± 0.8%) and estrus (80 ± 4.1%) (Figure 5B). All basal cells were ERα-negative, but approximately half of the luminal cells expressed ERα during both pro-estrus (50.6 ± 3.9%) and estrus (43.0 ± 4.4%) (Figure 5A). There was no significant difference in the proportions of PR-positive basal (p=0.47) or luminal cells (p=0.43), nor of ERα-positive luminal cells (p=0.21) during the different stages of estrus.
Although proliferation of mammary epithelial cells is mainly observed during the estrous phase of the murine estrous cycle, cells in S-phase, detected by [3 H]-Tdr labeling, were also observed during pro-estrus. During the latter, only 10 of 981 basal cells counted were labeled with [3 H]-Tdr, and seven of these were stained positively for PR (Figure 6A). In the luminal cell population, 34 of 36 [3 H]-Tdr-labeled cells were PR-positive (from 4735 cells counted), compared to only seven of 40 ERα-positive [3 H]-Tdr-labeled cells (from 6258 luminal cells counted) (Figure 6B).
During estrus, both basal and luminal cells were stimulated to undergo DNA synthesis. As in pro-estrus, the majority of [3 H]-Tdr-labeled basal cells were PR-positive (34 of 49 basal cells labeled with [3 H]-Tdr from 1007 counted). During estrus there was an approximately ninefold increase in the number of proliferating luminal cells, the majority of which were PR-positive (281 of 316 luminal cells labeled with [3H]-Tdr from 5323 counted). The proportion of ERα-positive proliferating luminal cells also increased during estrus to comprise approximately 60% of all proliferating cells observed compared with approximately 20% during pro-estrus.
Histograms comparing the mean proportions of cells ▪ positive and () negative for PR and ERα in all basal (A) and luminal cells (B) and in those labeled with [3 H]-Tdr (proliferating) in the basal (C) and luminal cell (D) populations in the ducts of mammary glands from 5-week-old mice.
Estrogen Receptor-β (ERβ) Expression
ERβ mRNA was detected in the mammary glands of both pubertal and adult mice in pro-estrus and estrus using RT-PCR (not shown) (Tremblay et al. 1998). Staining for ERβ revealed strongly positive ERβ immunoreactivity in granulosa cells of ovaries (Figure 7A), in contrast to the weak expression in mammary glands (Figures 7B and 7C). At high magnification, the cap and body cells of the TEBs in pubertal mice were very weakly positive for ERβ (Figure 7B), whereas in the ducts of adult mice (Figure 7C) the staining was more intense. In adult mice the majority of mammary epithelial cells that stained positively for ERβ were in the luminal cell population whereas only a few ERβpositive basal cells were observed. Cells in the fatty stroma were also weakly stained for ERβ, but those in the condensed fibroblast layer in the periductal regions were negative (Figure 7C). Lymphocytes, abundant in the mammary gland, were uniformly ERβ-negative.
Light micrograph of an adult (12-week) mammary duct from a mouse in estrus stained for PR. PR-positive basal cells labeled with [3 H]-Tdr are seen parallel to and within the basement membrane ▴. A PR-positive luminal cell labeled with 3H-Tdr is adjacent to one of these basal cells ↑. Nonproliferating PR-positive epithelial cells constitute the majority of the ductal cells. Bar = 16 μm.
Stacking bars indicate the relative proportion of cells positive (▪) or negative () for PR and ERα expression in all basal (A) and luminal cells (B) of adult mice in pro-estrus and estrus.

Cell counts of proliferating cells either (▪) positive or () negative for PR and ERα expression in the basal (A) and luminal (B) mammary epithelial cells of adult mouse mammary ducts during pro-estrus and estrus.
Discussion
Progesterone can stimulate mammary epithelial proliferation, and its specific receptor, PR, is expressed by a proportion of mammary epithelial cells (Shyamala 1997; Fendrick et al. 1998). In this study, the relationship between PR expression and proliferation in mammary epithelial cells was characterized histologically. The numbers of PR-positive cells labeled with [3 H]-Tdr, their distribution in the mammary gland, and their morphological type were assessed during normal pubertal development and adult estrous cycle. Furthermore, expression of ERα and ERβ in mammary epithelial cells was compared with these data.
In the mammary glands of pubertal mice, the majority of cells within the TEBs expressed PR, a finding consistent with the staining patterns presented in a previous immunohistochemical study (Silberstein et al. 1996). The majority of proliferating cap and body cells in the mitotically active TEBs were also PR-positive. Expression of PR by cells in the TEBs has led others to speculate a role for progesterone in directing proliferation during puberty (Silberstein et al. 1996). However, the finding that pubertal development was largely unaffected in mice with a disrupted PR (Lydon et al. 1995) has suggested that the PR pathway does not play a critical role during pubertal development. In light of these reports, the function of the PR pathway in the large number of PR-positive [3 H]-Tdr labeled cells in the TEBs of pubertal mice remains to be determined.
Proximal to the TEBs, the basal and luminal cells of the ducts are continuous with and regarded as being developmentally related to the cap and body cell layers of the TEBs, respectively (Williams and Daniel 1983). Consistent with this, the majority of luminal cells also expressed PR, as did the majority of luminal cells labeled with [3 H]-Tdr. In contrast, whereas less than 50% of the basal cells expressed PR, the finding that 80% of [3 H]-Tdr-labeled basal cells were PR-positive indicated that the PR pathway may play a role in directing their proliferation in pubertal ducts. Proliferation of basal cells during puberty may be specifically related to the formation of the side branches that arise from the elongating ducts. The hypothesis that PR is directing proliferation leading to ductal side branching is supported by the finding of severely impaired side branching in mice with a disrupted PR (Lydon et al. 1995).
Progesterone has been proposed as the principal mammary mitogen in adult mice (Shyamala 1997). In support of this, we observed that during estrus, when many mammary epithelial cells are recruited into S-phase, the majority of proliferating basal and luminal cells were PR-positive. Similar to pubertal mice, whereas the majority of basal cells were PR-negative, the majority of those labeled with [3 H]-Tdr expressed PR. During estrus, proliferation of mammary epithelial cells results in the formation of alveolar buds from the walls of mature ducts, and it has been proposed that these are derived from clusters of PR-positive epithelial cells (Atwood and Vonderhaar, personal communication). In agreement with a previous study (Silberstein et al. 1996), we did not observe clustering of PR-positive epithelial cells. However, PR expression did appear to be specifically associated with the proliferating basal cells, supporting the proposal of a PR-mediated pathway in the formation of alveolar buds.
Estrogen has an established role in stimulating mammary proliferation. However, in the present and in a previous study we have demonstrated that the majority of proliferating mammary epithelial cells do not express ERα (Zeps et al. 1998). In particular, the cap cells of the TEBs and the basal cells of the ducts in both pubertal and adult mice were uniformly ERα-negative. Although estrogen induces PR expression in adult mice (Shyamala 1997), our study indicates that all PR expression in basal cells of the adult mammary ducts occurred in the absence of detectable ERα expression. Furthermore, a comparison of the proportion of ERα-positive and PR-positive luminal cells reveals that approximately 30% do not co-express these two receptors in adult mammary ducts. Recently, studies in ovariectomized mice have demonstrated constitutive expression of PR in a proportion of mouse mammary epithelial cells (Shyamala et al. 1997). In other studies, it has been suggested that estrogen-induced responses, such as PR expression, are mediated indirectly by paracrine mechanisms involving epidermal growth factor (EGF), insulin-like growth factors (IGFI and II), and transforming growth factors (TGF-α and -β (Imagawa et al. 1990; Katzenellenbogen and Norman 1990; Haslam and Counterman 1991; Fendrick et al. 1998).
The identification of ER(expression in the mammary glands of pubertal and adult mice in the present study was anticipated by the finding of ERβ mRNA in the mammary gland (Tremblay et al. 1998; and our unpublished data). Our results indicate low-level but widespread expression of ERβ in epithelial and stromal cells. Although the reported lack of developmental abnormalities in the mammary glands of mice with a disrupted ERβ (BERKO mice) suggests that ERβ is not critical to normal mammary gland development or function (Krege et al. 1998), it remains feasible that the ERβ pathway is activated by and can mediate estrogen effects in the normal mouse mammary gland.
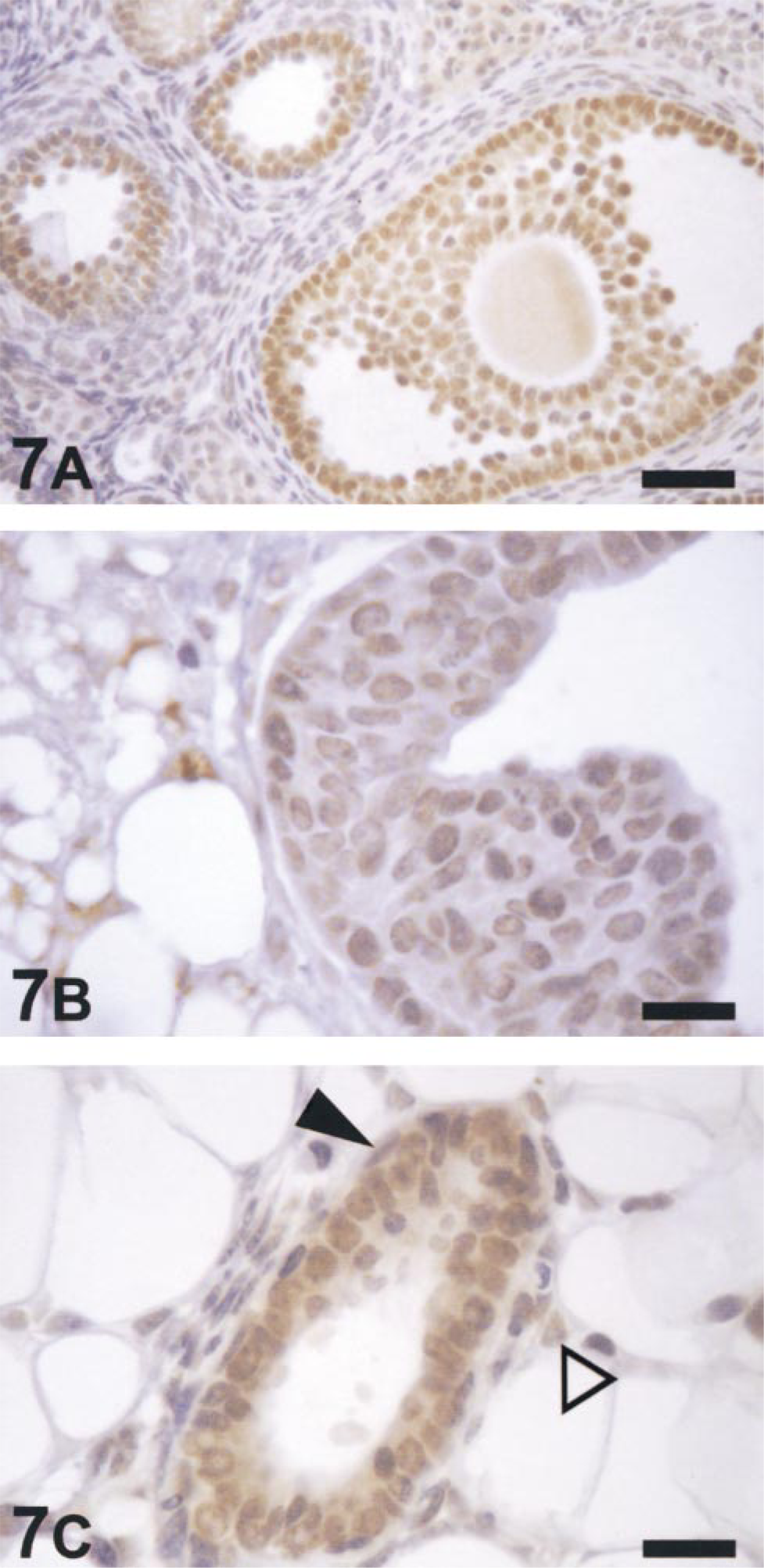
Light micrographs of immunohistochemical staining for ERβ in an ovary from a 10-week-old mouse during pro-estrus (A), a TEB in a mammary gland from a 5-week-old mouse (B) and a duct in a mammary gland from a 12-week-old mouse in pro-estrus (C). Positive staining was strong in the granulosa cells of the ovary (A) but weak in the cap and body cell populations of the TEB (B) and in the basal and luminal cells of the mammary duct (C). Cells immunoreactive for ERβ were predominantly luminal in the mammary duct with a few basal (▴) and stromal (Δ) cells also staining positive. Bars: A = 40 μm; B,C = 16 μm.
In summary, this study has demonstrated that epithelial cells of the pubertal and adult mammary glands in mice exhibit a characteristic expression of the steroid hormone receptors ERα, ERβ, and PR. Expression and co-expression of these receptors can be related to proliferation of specific cell types in the gland and are likely to direct formation of tertiary structures in the mammary gland, such as side branching in the pubertal gland and alveolar bud formation in the adult gland. Further mapping of co-expression of steroid hormone receptors in individual cells and proliferation or migration of these cells during puberty or in the adult mouse will delineate the functional roles of the steroid hormones in development and maintenance of the mammary gland and in the preparation of the mammary ducts for pregnancy and lactation.
Footnotes
Acknowledgements
Supported by an Arnold Yeldham and Mary Raine Medical Research Foundation grant and by the Urological Research Centre, University of Western Australia. NZ was supported by a Raine/SGIO scholarship.
We thank Ms Sharon Redmond for technical advice and Mr Janni Mirosevich for help with figure layout.