
Abstract
Difficulties in demonstrating sex steroid receptors in the human brain by immunohistochemistry (IHC) may depend on postmortem delay and a long fixation time. The effect of different postmortem times was therefore studied in rat brain kept in the skull at room temperature for 0, 6, or 24 hr after death. After a long fixation for 20 days, hypothalami were embedded in paraffin and sections were immunohistochemically stained for androgen receptor (AR), estrogen receptor-α (ER), or progesterone receptor (PR). Retrieving the antigenic sites by microwave pretreatment was essential to obtain successful IHC in all groups studied. In general, immunoreactivity was restricted to the cell nuclei. However, the intensity of the staining appeared to be strongly dependent on the different receptor antigens and postmortem time. Both AR and ER but not PR immunoreactivity were decreased after immersion-fixation compared to the perfused sections at time point zero. In brains fixed by immersion, all three receptors decreased gradually with increasing postmortem time, and ER became hardly detectable after 24 hr postmortem. The results of these experiments show that, with the protocol used, postmortem variables and lengthy fixation do not, in principle, prevent sex steroid receptor IHC in human material. The outcome of the immunostaining, however, might be strongly dependent on the epitopes and/or antibody used.
Keywords
T
The effect of postmortem delay on steroid receptors is strongly dependent on the technique used to visualize them. In spite of the labile receptor binding properties of corticosteroids during a postmortem period of 3–4.5 hr (Sapolsky and Meaney 1988), postmortem stability of the mRNAs for glucocorticoid and mineralocorticoid receptors was found for 12–24 hr after death using in situ hybridization histochemistry (Wetzel et al. 1994). However, as far as the immunohistochemical visualization of sex steroid receptors was concerned no parametric data were available. To clarify the effect of postmortem delay, we have therefore systematically tested the stability of various sex steroid receptors using immuno-histochemistry in the rat hypothalamus and adjacent structures obtained within 0–24 hr after death and fixed for 20 days, as is commonly done in human studies.
Material and Methods
Animals and Tissue Preparation
Male Wistar rats were purchased from Harlan CPB (Zeist, The Netherlands). All aspects of animal care and treatment were carried out according to the local guidelines of the experimental animal care committee (No. 95.19/00). Rats weighing 200 ± 20 g (n = 20) were sacrificed by decapitation and the brains were either removed immediately (n = 5) or left in the skull for 6 (n=5) or 24 h (n=5) at room temperature (RT). For comparison, five intact animals were deeply anesthetized with sodium pentobarbital (0.1 ml/100 g body weight) and perfused through the ascending aorta with 50 ml of physiological saline, followed by 300 ml of fixative containing 4% paraformaldehyde in 0.1 M PBS, pH 7.4. All groups were then treated as described below. After a postmortem delay of 0, 6, or 24 hr the brains were removed from the skull and postfixed for 20 days in 4% paraformaldehyde solution in 0.1 M PBS at 4C. The forebrains were processed for routine paraffin (Histowax) embedding. Coronal sections (8 μm) were cut, mounted on 3-amynopropyltriethoxisilane-coated slides (Menzel; Darmstadt, Germany) to prevent detachment due to the high temperature during antigen retrieval treatment. Antigen retrieval was performed by microwave oven heating according to Shi et al. (1993). After deparaffinization and rehydration, the slides were placed in a jar filled with 0.01 M citrate buffer (pH 6.0) and heated in a microwave oven, three times for 5 min at 700 W. The sections were allowed to cool down to RT. Meanwhile, adjacent sections were left in citrate buffer at RT to verify the effect of the microwave treatment.
Immunohistochemistry
After rehydration, the sections were first rinsed extensively with PBS and incubated overnight in one of the primary antibodies diluted in Tris-buffered saline (TBS) containing 0.05 M Tris, 0.5 M NaCl, 0.5% Triton X-100, and 0.1% sodium azide, pH 7.6, at 4C. The primary antibodies included PG 21 (1:500) (gift of Geoffrey Greene, University of Chicago), which is a rabbit polyclonal androgen receptor (AR) antibody directed at the first 21 amino acids at the N-terminus of the rat/human AR (Prins et al. 1991; Zhou et al. 1994), 1D5 (1:50) (DAKO; Glostrup, Denmark), which is a mouse monoclonal estrogen receptor-alpha (ERa) antibody raised against amino acids 118–140 in the A/B domain of the human ER (Al Saati et al. 1993), or a polyclonal progesterone receptor (PR) antibody (1:500) (DAKO), which was made against amino acids 533–547 in the DNA-binding domain of the rat/human PR (Traish and Wotiz 1990). After several washes in TBS, sections were incubated with biotinylated secondary antisera (biotinylated anti-rabbit IgG for AR, PR and biotinylated anti-mouse IgG for ER, 1:250 in TBS; Vector Laboratories, Burlingame, CA) for 1.5 hr at RT. This was followed by a 1-hr incubation at RT with avidin-biotin-peroxidase (1:400; ABC Elite Kit, Vector Laboratories). Tissue-bound peroxidase was visualized with the 3,3′-diaminobenzidine tetrahydrochloride (DAB) chromogen reaction intensified with nickel-ammonium sulfate (7.5 mg DAB, 300 mg nickel-ammonium sulfate, 5 μl 30% H2O2 in 15 ml 0.05 M Tris-HCl buffer (TRIS), pH 7.6) for 6–15 min at RT. Sections were then rinsed in distilled water, dehydrated in a graded series of ethanol, cleared in xylene, and cover-slipped with Entellan (Merck; Darmstadt, Germany). Alternate sections were stained with thionin for orientation.
Quantitative Analysis
To determine the postmortem effects in the staining intensity of AR/ER/PR-containing neurons, immunoreactivity was evaluated by optical density measurements. Brain regions rich in a given receptor subtype were studied, including the lateral septum (AR), arcuate nucleus (ER, PR), and medial preoptic nucleus in which all three receptors (AR, ER, PR) were present. Two sections per cell group per animal were analyzed for a total of 100 sections using an IBAS image analysis system (Kontron Instruments; Munich, Germany). Sections were viewed under brightfield illumination with a Zeiss Axioplan microscope with a SONY CCD XC-77CE black-and-white camera. Gain and black level of the camera were fixed. A place outside the tissue was chosen for 100% transmission (gray level 255).
The same illumination and analog settings were used during acquisition and analysis of all images. The gray values per pixel were transformed into optical density (OD) per pixel and average OD per cell was measured. For each cell group from each animal, OD readings from two sections (20 cells) were summed. To analyze background staining, five OD readings per section were taken from a cell-sized area of neuropil in the vicinity of labeled neurons.
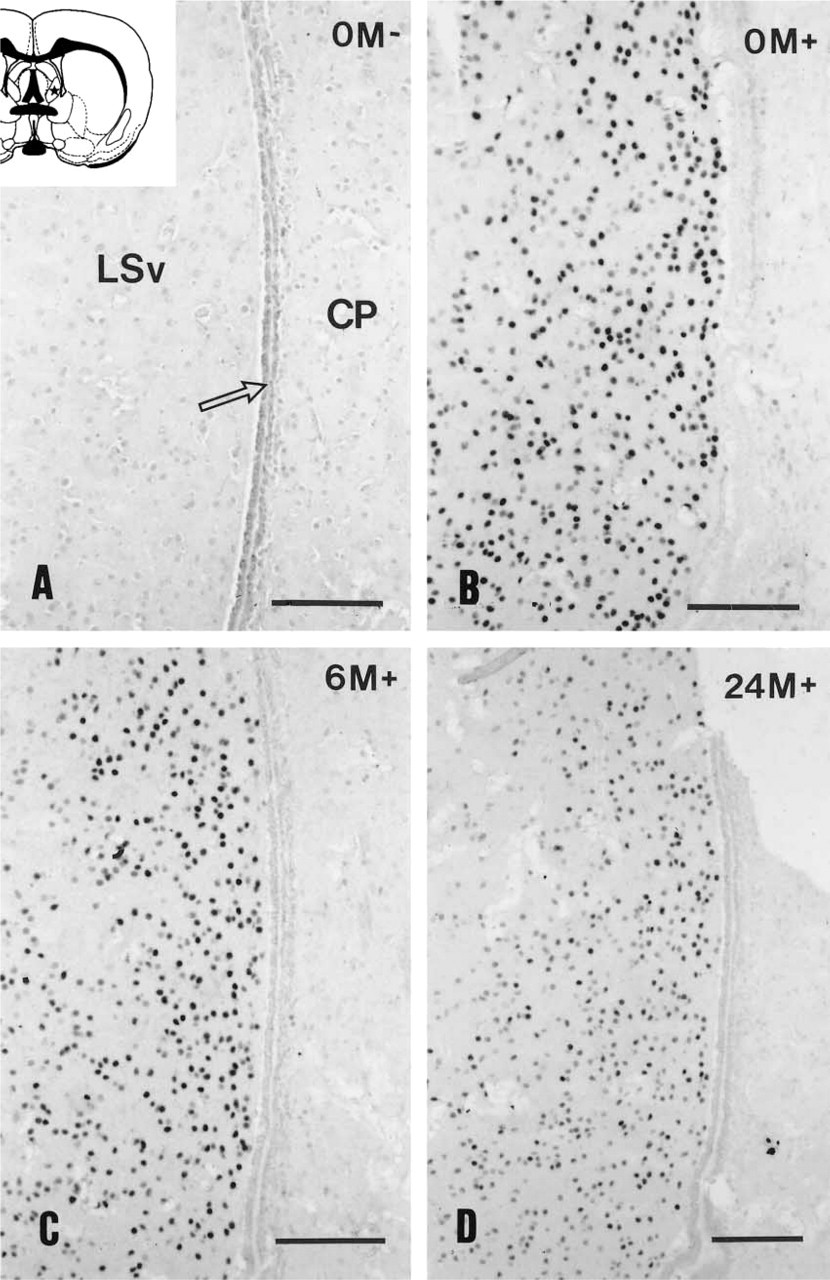
Androgen receptor immunohistochemical staining of the ventral part of the lateral septum (LSv) in rat brain that was immediately fixed by immersion (
The differences between the intensity of the immunohistochemical staining for each time point was analyzed by ANOVA tests. Differences were considered statistically significant at p<0.05.
Results
Paraffin-embedded sections of rat forebrain showed specific nuclear labeling for all three sex steroid receptors studied, in spite of the long fixation period of 20 days. However, for visualization of the immunoreactivity microwave irradiation was essential. An example is given in Figure 1, in which the lateral septum (AR-containing region) and the neighboring caudate putamen complex (AR-negative region) are shown. In general, sections without microwave pretreatment showed nonspecific reaction in the ependymal layer of the ventricular system, high background, and absence of specific nuclear staining all over the section (Figure 1A), in contrast to the microwave-pretreated sections, in which specific nuclear staining became visible only in the expected area and the ependymal staining disappeared (Figure 1B).
Androgen Receptor Immunoreactivity
Cells containing AR immunoreactivity were widely distributed throughout the intact rat forebrain. The highest number of cells was found in the lateral septum (Figure 1), the bed nucleus of the stria terminalis, amygdala, medial preoptic area, and the hypothalamic ventromedial, arcuate, and premamillary nuclei. Immunopositivity was restricted to the cell nuclei as was shown previously in immunohistochemical staining in the gonad-intact male (Wood and Newman 1993). The subcellular localization was not affected by postmortem time. One-way ANOVA revealed significant effects in the medial preoptic nucleus (F= 10.229; p<0.001) and the lateral septum (F=7.148; p<0.003). However, in the brains fixed immediately by perfusion (not shown) or immersion (Figure 1B) there was no visible difference in the distribution pattern or the intensity of the immunoreactivity (Table 1). After 6- and 24-hr postmortem delay the intensity of the immuno-staining was reduced noticeably (Table 1). However, neither the background nor the distribution pattern changed significantly (Figures 1C and 1D; Table 1). No significant differences in the two areas measured were observed at various time points (Table 1).
Estrogen Receptor Immunoreactivity
ER immunoreactivity was distributed as previously described (Cintra et al. 1986). Dense populations of ER-positive neurons were found in the medial preoptic nucleus (Figure 2), bed nucleus of the stria terminalis, and arcuate and ventromedial nuclei. The staining was confined to the cell nuclei independently of the postmortem delay. Nevertheless, the intensity of immunohistochemical staining was dependent on the type of fixation. At time point zero the intensity of immunoreactivity in the transcardially perfused material (Figure 2A) was superb compared to the non-perfused immersion-fixed tissue (Figure 2B). Although the distribution pattern was identical, immersion-fixation resulted in noticeably decreased immunoreactivity in all regions. The nuclear staining was more intense and homogeneous in the perfused animals (Figure 2A) compared to the immersion-fixed sections, in which a decrease in immunoreactivity was visible to various degrees in the positive nuclei (Figure 2B). Signal was still detectable after 6 hr at variable intensities (Figure 2C), but after a 24-hr delay ER immunoreactivity was hardly visible (Figure 2D). The diminution of the staining was confirmed by the OD measurements (Table 1), and ANOVA revealed significant effects on the medial preoptic nucleus and arcuate nucleus (F=8.135 and 8.722; p<0.002). No significant difference was measured for the two nuclei analyzed at various time points. Prolonged chromogen reaction did not result in enhancement of the specific staining but instead caused nonspecific reaction in the cytoplasm of neurons in the supraoptic and paraventricular nuclei, all cortical layers, and glial cells in the optic tract (not shown).
Optical density readings after different postmortem times (in hr) a
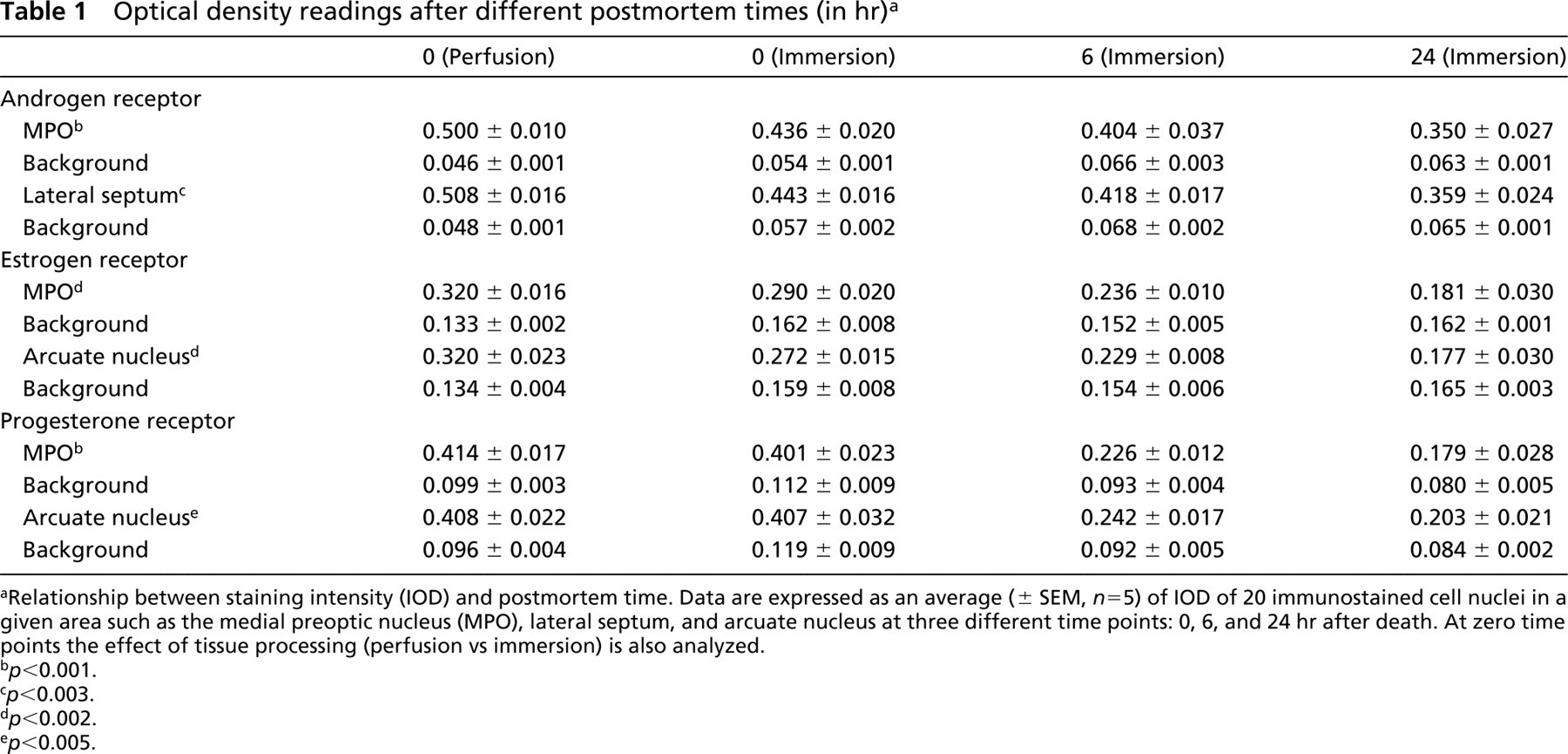
aRelationship between staining intensity (IOD) and postmortem time. Data are expressed as an average (± SEM, n=5) of IOD of 20 immunostained cell nuclei in a given area such as the medial preoptic nucleus (MPO), lateral septum, and arcuate nucleus at three different time points: 0, 6, and 24 hr after death. At zero time points the effect of tissue processing (perfusion vs immersion) is also analyzed.
b p<0.001.
c p<0.003.
d p<0.002.
e p<0.005.
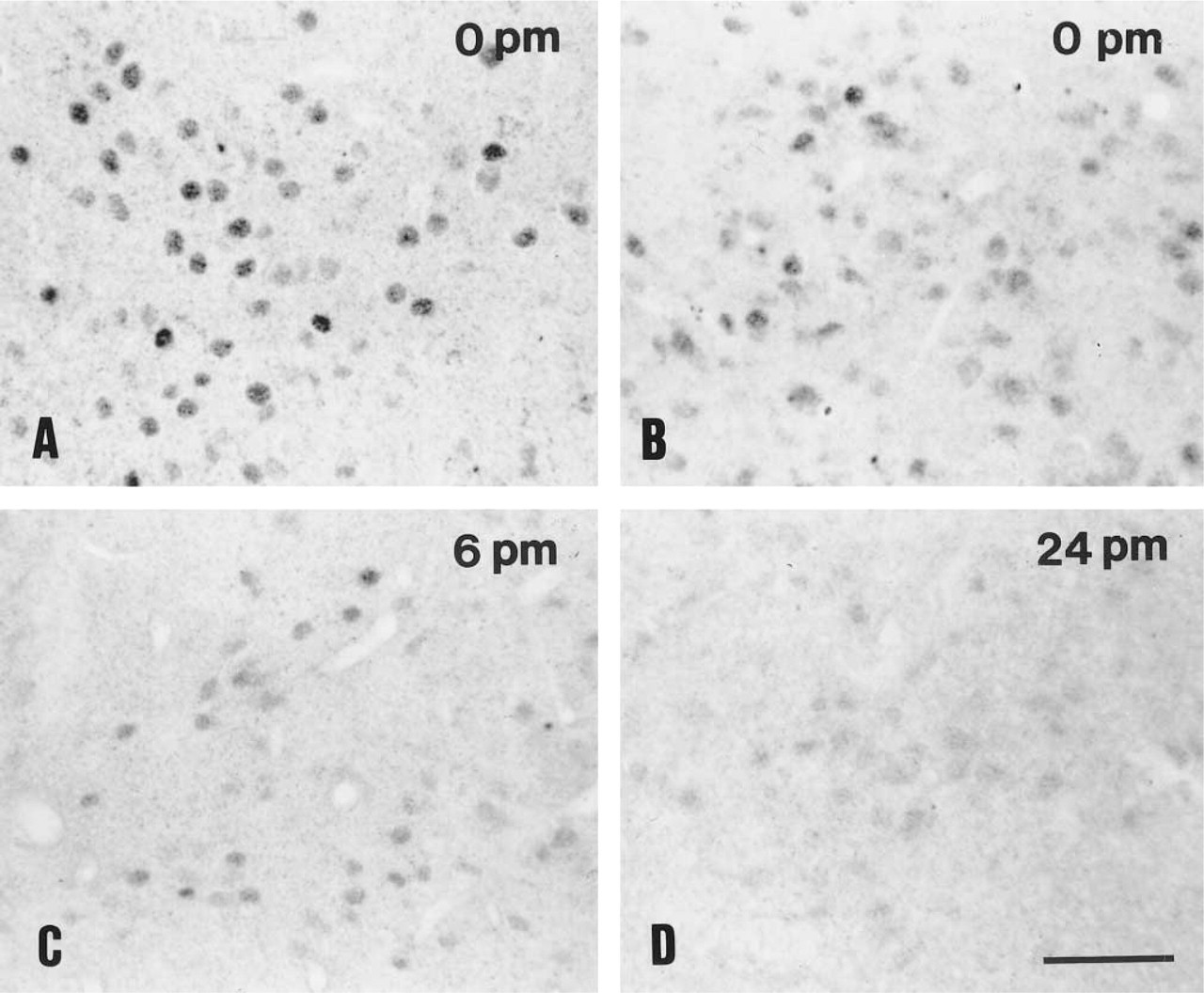
Representative photomicrographs of estrogen receptor immunopositivity in the intact male rat. Medial preoptic area from brain fixed by perfusion (
Progesterone Receptor Immunoreactivity
At time point zero, PR immunoreactivity was observed in cell nuclei of neurons in the preoptic area and mediobasal hypothalamus (Figure 3) as previously described (Warembourg et al. 1989; Bethea et al. 1992). No significant difference in the distribution pattern or the number of immunopositive nuclei was seen between the perfused and immersion-fixed groups at time point zero (Figures 3A and 3B). However, the intensity of the PR immunoreactivity was dramatically reduced by approximately 30%, in brains left for 6 hr at RT (Figure 3C). Very weak immunostaining was still detectable after 24 hr in virtually all regions in the cell nuclei (Figure 3D). In a small number of cells, however, we did find stronger immunostaining. In these cells the chromatin appeared to be looser and immunoreactivity also appeared in the cytoplasm (Figure 3D). One-way ANOVA showed significant effects of postmortem time in the medial preoptic nucleus (F=65,202; p<0.001) and arcuate nucleus (F=22,265; p<0.005). No significant difference was measured in the two regions analyzed at various postmortem time points.
Discussion
This is the first report evaluating postmortem effects on sex steroid receptor immunohistochemistry in paraffin-embedded rat brain. In recent years, several studies have indicated that immunohistochemical demonstration of sex steroid receptors in the brain of different species, including humans, is feasible. The descriptive studies in mammalian brain, with one exception (Okamura et al. 1992), have been carried out in perfused floating material or frozen specimen (Cintra et al. 1986; Warembourg et al. 1989; DonCarlos et al. 1991; Fox et al. 1991; Bethea et al. 1992; Lehman et al. 1993; Li et al. 1993; Iqbal et al. 1995; Michael et al. 1995; Sisk and DonCarlos 1995; Van Leeuwen et al. 1995; Thind and Goldsmith 1997; Shughrue and Merchenthaler 2001). In contrast, the majority of the human studies available, because of the aforementioned practical reasons, were done mostly on paraffin-embedded material (Puy et al. 1995; Fernández-Guasti et al. 2000; Ishunina et al. 2000; Taylor and Al-Azzawi 2000; Ishunina and Swaab 2001; Kruijver et al. 2001). Regardless of the species and tissue processing, the majority of these reports agreed that the duration of the (post)fixation time and the intensity of the immunoreactivity were inversely correlated because of epitope masking caused by the aldehyde-containing fixatives. It was also reported that the optimal staining on free-floating material could be obtained from brains that were shortly perfused and postfixed for only a few hours.
For systematic, retrospective, and quantitative work on the human brain, however, the most convenient way to collect brain material is to embed the tissue block in paraffin after appropriate fixation. Experimental animals are mostly transcardially perfused with the fixative, whereas human autopsy material is usually immersion-fixed. Working with human tissue usually involves additional unfavorable factors such as the variable postmortem delay, no perfusion, and therefore inadequate fixation because fixation of the outer parts of the tissue blocks forms a barrier. As a consequence, a longer fixation time is necessary for the larger human brain structures (Swaab 1997), which may critically affect the preservation of sex steroid receptors. In addition, antemortem events in the brain, such as hypoxia and chronic disease, or the use of medications (Erkut et al. 1998) may exert an important influence on a number of neurochemical parameters. Moreover, mRNA preservation, enzyme activities, and neurotransmitter and neuropeptide concentrations have all been shown to vary significantly with varying agonal state (Spokes 1979; Perry et al. 1982; Hardy et al. 1985).
The aim of our study was to see whether postmortem delay, independently of other factors, might affect sex steroid receptor immunohistochemistry. For that purpose we used intact adult male rats because we wanted to avoid cyclic changes in the hormonal milieu.
To detect the steroid receptor proteins in the paraffin-embedded rat brain it was necessary to pretreat the samples to open the tissue matrix and to minimize the crosslinkage caused by the aldehyde fixative (Shi et al. 2001). Previous qualitative studies on steroid receptor-positive carcinomas showed the superiority of microwave pretreatment over other “unmasking” techniques such as enzyme digestion (Shi et al. 1993; Szekeres et al. 1994). Therefore, we chose to microwave the sections in citrate buffer at pH 6. Without this pretreatment no immunostaining was observed, most probably due to “masking” of the antigenic sites. Although several studies showed that the composition of buffer can influence the quality of the immuno-staining (Lucassen et al. 1993; Shi et al. 1993,1995), we could not confirm these data for nuclear receptor antibodies in our laboratory. We also tested whether the pH of the antigen retrieval buffer might influence the quality of the immunohistochemical staining using pH 2, 4, 6, and 8 in citrate buffer. Our results suggested that the optimal staining for steroid receptors could be obtained at pH 6 (unpublished results).
Ischemia after decapitation may lead to serious deleterious effects on receptor staining at the protein level. Loss of immunoreactivity was observed in the estrogen and androgen receptor immunostaining compared to perfusion data. These results indicate that ischemic effects are dependent on receptor types and/or antigen epitopes. It is important to note, however, that in all groups immunohistochemically stained for ER, AR, or PR, the distribution pattern of immunopositive cells did not appear to differ between these two conditions at the zero time point. Although sex steroid receptor-containing nuclei were still visible 6 hr after death, the immunostaining was gradually and markedly decreased. ER became hardly detectable after a 24-hr postmortem delay. Weakly stained PR-immunoreactive cell populations were still visible 1 day after death, but the staining was less homogeneous due to strongly labeled single neurons. In comparison to ER and PR, however, after 1 postmortem day a less dramatic decrease in the AR immunoreactivity was observed. Taken together, ER and PR immunostaining was most prominently influenced by the postmortem time.
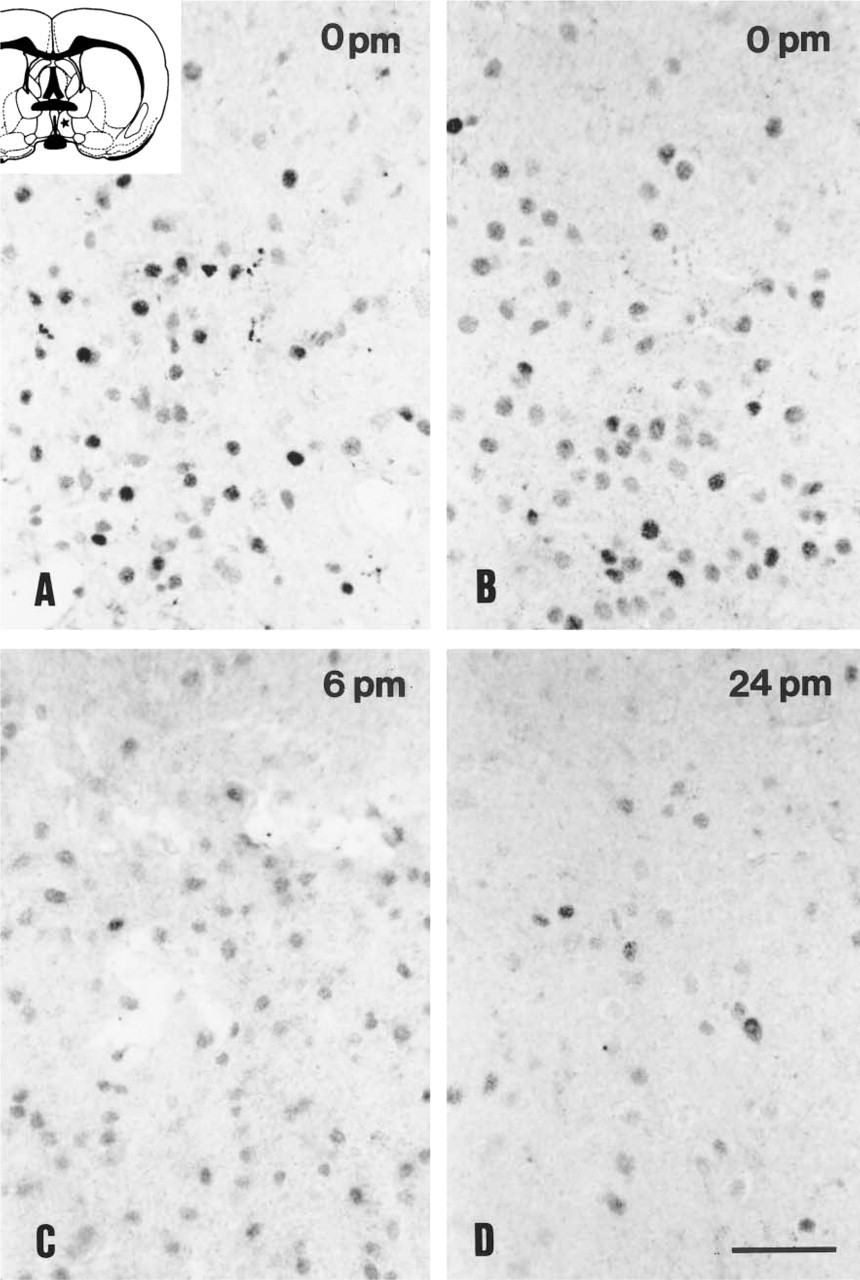
Progesterone receptor-containing neurons in the male rat hypothalamus fixed for 20 days, pretreated with microwave irradiation and immunostained with the Dako antibody. The medial preoptic area from brains fixed by perfusion (
Speculation that the length of fixation might result in cytoplasmic staining, as was reported earlier on free-floating material (Blaustein 1992), was not supported by our observations. The only change in the subcellular distribution occurred in a few PR-containing neurons after a longer postmortem time. In these cells, next to the nuclear staining perinuclear cytoplasmic staining was also observed, probably due to postmortem decomposition. Our present data showed, independently of the duration of postmortem time, that all three sex hormone receptors reside predominantly within the nuclear compartment. Furthermore, with either type of fixation (perfusion vs immersion), the staining pattern of receptor-containing tissues stayed exclusively nuclear in the paraffin-embedded rat brain. One should keep in mind that these data were obtained by using a given antibody for each receptor. Staining intensity can be different when antibodies against other epitopes of the same receptors are applied. Therefore, we suggest that before starting a quantitative study on human paraffin-embedded material the antibody's postmortem characteristics should be tested on a series of sections similar to those used in the present study.
Taken together, postmortem delay and long fixation time are believed to prevent successful immuno-histochemical staining of steroid receptors. The present study shows that the detrimental effect of prolonged fixation can be effectively reversed by microwave treatment. Although immunoreactivity of sex steroid receptors does not immediately deteriorate due to postmortem delay, the quality of the staining is very much dependent on antigens and antibodies.
Footnotes
Acknowledgements
Supported by NWO (#903–47004) to MF.
We thank Drs Gail S. Prins and Geoffrey L. Greene (University of Illinois, Chicago, IL) for the PG-21 antibody, Dr C. W. Pool and Mr J. J. van Heerikhuize for the IBAS program, Dr M. Diamant for the statistical analysis, Dr M. Hofman for critically reading the manuscript, and Mr G. Van der Meulen for the photography.