
Abstract
The 16-kD subunit of the vacuolar H+ -ATPase (V-ATPase), or ductin, is essential for the activity of this proton pump and has roles in intercellular communication and control of cell growth and differentiation. The V-ATPase is important for acidification-dependent degradation of tissue matrices through which some cell types move, and for pH regulation across some epithelial cell layers. Placentation involves intricate signaling, cell proliferation, and controlled invasion. We examined the distribution of three subunits of the V-ATPase in bovine trophoblast and endometrium at the time of implantation to determine the relationship of ductin expression to that of two other subunits, A (approximately 73 kD) and B (approximately 58 kD). Epithelial expression of all three subunits was observed, and in nonpregnant animals this expression was apical. As pregnancy proceeded, expression of all subunits became pericellular in luminal but not glandular epithelium, suggesting a redistribution of V-ATPase activity. The trophoblast expressed all three subunits during initial contact with the epithelium. In the stroma, ductin expression was reduced after implantation, and we discuss the possibility that ductin plays a role in the shifting communication between stromal and epithelial cells induced by embryo attachment.
T
The acidification of cytoplasmic vesicles by V-ATPase provides an environment within which receptor-ligand complexes are targeted for degradation or recycling to the cell surface (Brown et al. 1983; Johnson et al. 1993; Clague et al. 1994; Reaves and Banting 1994). In this way, the enzyme is an important component of intercellular communication pathways. The ATPase activity is responsible for acidification of the extracellular environment because unlike other ATPases, it is able to extrude hydrogen ions without the need for a counter-ion. This extracellular acidification is critical for processes such as bone resorption by osteoclasts (Blair et al. 1989; Tuukkanen and Lakkakorpi 1990) and scavenging by macrophages (Heming et al. 1995). Furthermore, the importance of V-ATPase for maintenance of cytoplasmic pH is highlighted by observations that inactivation of the enzyme leads to apoptosis as a result of acidification (Gottlieb et al. 1995; Kinoshita et al. 1996; Long et al. 1998).
In addition to its role in the V-ATPase pump, ductin is found as a hexameric gap junction protein in diverse tissue types (Finbow et al. 1993, 1994). In a survey of human pancreatic tumor samples, 42 of 46 samples from invasive cells showed reactivity to ductin, whereas all of the 13 samples of noninvasive tumors had little detectable ductin (Ohta et al. 1996).
The molecular and cellular events that occur during initiation of placental development in mammals lead to dramatic physiological changes of the uterine endometrium. The exact molecules that initially serve to tether the embryo to the epithelium are probably different among species, and include glycoproteins and adhesion molecules. After attachment of the embryo to the uterine epithelium, limited invasion occurs and there is extensive remodeling of the epithelium and underlying stroma, including stromal proliferation and vascularization. A reduction in electrostatic charge has been observed in many species before attachment. The abundance of the V-ATPase, or ductin, in invasive osteoclasts, macrophage, and metastatic cells, and the involvement of the enzyme in acidification of vesicles or the outer membrane environment, suggests that it could play a significant role in implantation.
In the cow, embryo attachment occurs 17 to 18 days after fertilization, by which time the trophoblast has elongated to several centimeters (King and Thatcher 1993). Trophoblast-derived binucleate cells fuse with uterine epithelial cells, creating trinucleate cells that represent the full extent of invasion into maternal tissue (Ireland et al. 1979; King et al. 1982; Hoffman and Wooding 1993). We report here on an analysis of the distribution of subunits A, B, and c (ductin) of V-ATPase during and after bovine embryo attachment.
Materials and Methods
Animals
Heifers of various breeds were synchronized with prostaglandin F2α (Lutalyse; Pharmacia-UpJohn, Orangeville, Ontario, Canada) and bred approximately 12 hr after the onset of estrus (Day 0). Reproductive tracts were collected after exsanguination and the ovaries and uterus examined to confirm stage of cycle and/or pregnancy. Three tracts each at Days 18, 21, and 24 of pregnancy and at Day 18 of the estrous cycle were cut into blocks and snap-frozen in liquid nitrogen. Blocks were stored at −80C until cryosectioning. Eight-μm sections were acetone-fixed on aminopropyltriethoxysilane (ICN; Costa Mesa, CA)-coated slides.
Immunofluorescence
Frozen sections were thawed and rehydrated in PBS. Sections were blocked in PBS containing 10% preimmune rabbit serum for 30 min, then incubated with polyclonal rabbit anti-mouse ductin (raised for us by Alpha Diagnostics; San Antonio, TX) against an N-terminal eight amino-acid peptide, NPEYSSFF), rabbit anti-human subunit A (Lee et al. 1996), or chicken anti-human subunit B (Lee et al. 1996). Antibodies were diluted in PBS containing 2% preimmune rabbit serum and incubated for 1 hr at room temperature (RT). The sections were washed for 3 min in PBS, then incubated for 30 min with Alexa 488-conjugated goat anti-rabbit (Molecular Probes; Eugene, OR) or FITC-conjugated goat anti-chicken (Sigma; St Louis, MO) secondary antibodies. Sections were then given three 10-min washes in PBS, mounted with mounting medium (Sigma), and viewed with a Bio-Rad MRC-600 laser scanning confocal microscope (LSCM) equipped with a krypton-argon mixed-gas laser. Adjacent sections used as negative controls were incubated with preimmune rabbit serum or with chicken IgG instead of primary antibody. No fluorescence was detected in these negative controls. Sections through horse liver and kidney, tissues in which the V-ATPase is highly abundant, were used as positive controls for staining.
Western Blot Analysis
The crossreactivity of the antibodies used in immunofluorescence studies was assessed by Western blot analysis. Whole-cell lysates were prepared from bovine kidney. Samples of kidney were frozen in liquid nitrogen and ground with a mortar and pestle. The powder was lysed in RIPA buffer and cell debris was removed by centrifugation. An equal amount of SDS-PAGE loading buffer was added and the samples were boiled for 10 min. The total cell protein was electrophoresed on a 10% polyacrylamide gel, transferred to PVDF membrane in CAPS (3-[cyclohexylamino]-1-propane-sulfonic acid) buffer and incubated at RT with either anti-subunit B or anti-subunit A antibody. After two 10-min washes in PBS, Western blot detection of subunit B was achieved using rabbit anti-chicken IgG unconjugated antibody (Pierce Biochemicals; Rockford, IL), followed by incubation with alkaline phosphatase-conjugated sheep anti-rabbit IgG (Roche Molecular Biochemicals; Indianapolis, IN) and color development with the alkaline phosphatase substrates NBT and BCIP. Subunit A protein was similarly detected, using sheep anti-rabbit IgG alkaline phosphatase-conjugated secondary antibody.
Results
Ductin Expression in Nonpregnant and Pregnant Animals
The distribution of V-ATPase subunits in the endometrium of nonpregnant animals at Day 18 of the estrous cycle was similar but not identical to that of pregnant animals at Day 18 of gestation. In cycling cows (i.e., nonpregnant), both glandular and luminal epithelium expressed ductin preferentially on the apical surface (Figure 1A). In pregnant animals at Day 18, ductin expression on luminal epithelium began to shift to both apical and basal surfaces (Figure 1B), so that by Day 21 similar levels of ductin were seen on both surfaces, a shift that was absent in glandular epithelium (Figure 1D). After attachment, by Day 21, ductin expression in the stroma was downregulated (compare Figures 1B and 1D). The extent of this reduction is difficult to ascertain because immunohistochemistry is not a very quantitative assay, but it was a recurrent observation. This downregulation was transient, as expression is restored by Day 24 (Figure 1E).
By Day 24 there was extensive remodeling of the luminal epithelium and trophectoderm as embryo development and attachment accelerated. At this stage, expression of ductin in the epithelium was essentially pericellular (Figure 1F), whereas the distribution in glandular epithelium continued to remain primarily apical (Figure 1E). In the developing trophoblast, ductin expression at Day 18 was evident throughout trophoblast cells proximal to the uterine epithelium (Figures 1B and 1C) and continued to persist at Day 24, primarily at the interface with luminal epithelium (Figures 1E and 1F).
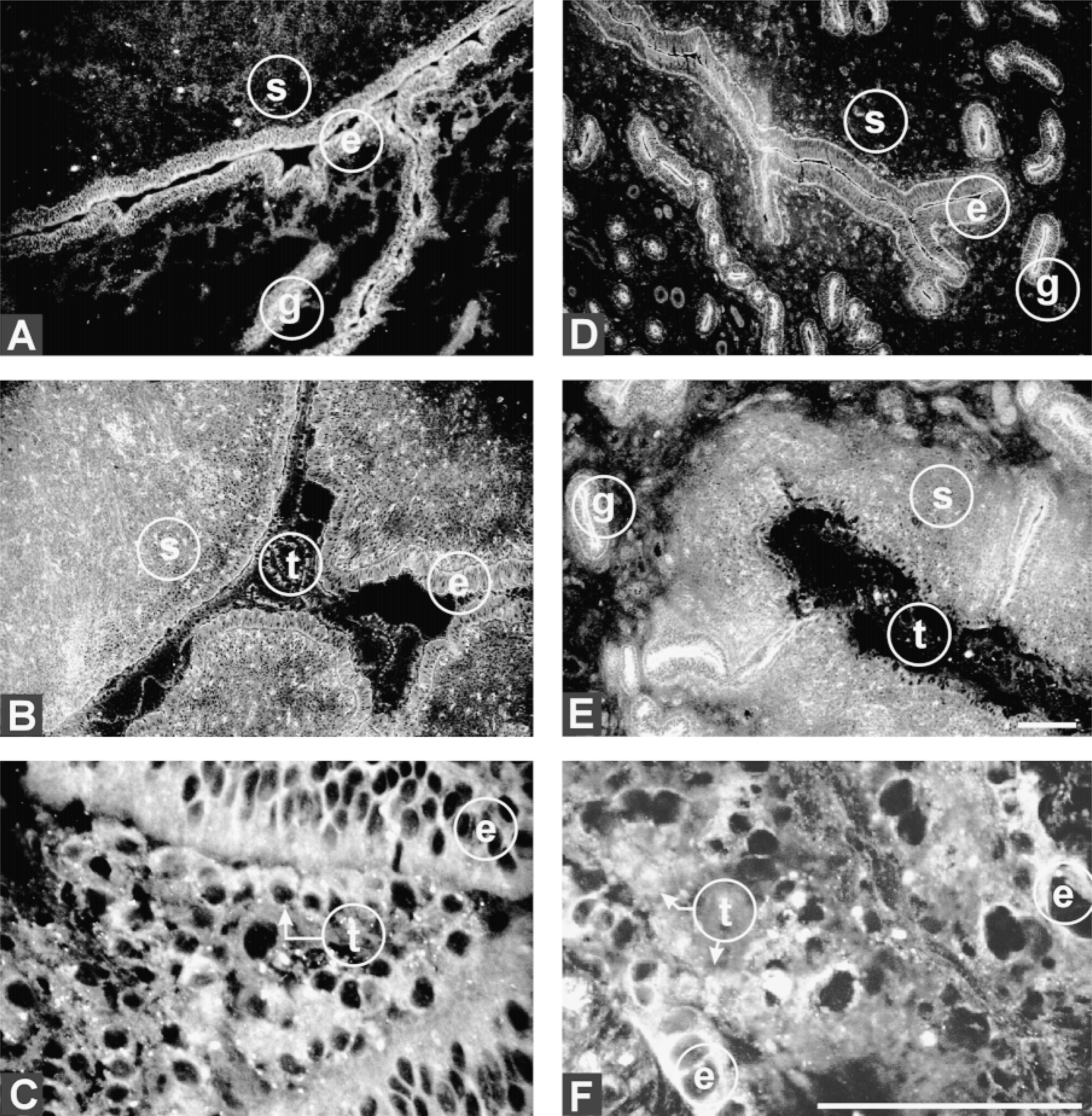
Ductin expression in the uterus of nonpregnant and pregnant bovine and in trophoblast of the developing embryo. (
In all analyses, adjacent sections incubated with control preimmune serum or chicken IgG served as negative controls and permitted background levels of fluoresence to be subtracted from the images. No fluorescence was detectable at the levels used to take the positive images.
Expression of V-ATPase Subunit A in Pregnant and Nonpregnant Animals
Two other subunits of the V-ATPase were similarly analyzed to determine whether their expression matched that of ductin. Subunit A, like ductin, was primarily apical in both glandular and luminal epithelium of Day 18 nonpregnant animals (Figure 2A). In pregnant animals, by Day 18 its distribution had become peri-cellular within the luminal epithelium (Figures 2B and 2C) but remained unchanged in glandular epithelium. At Day 21 there was enhancement of apical and basal distribution (Figure 2D). Throughout the remodeling epithelial cells at Day 24, expression became pericellular, similar to that seen with ductin (Figure 2F). The expression of subunit A remained constant in the uterine stroma throughout the stages of pregnancy examined (Figures 2B, 2D, and 2E). There was no change in expression in glandular epithelium from Day 18 to Day 24.
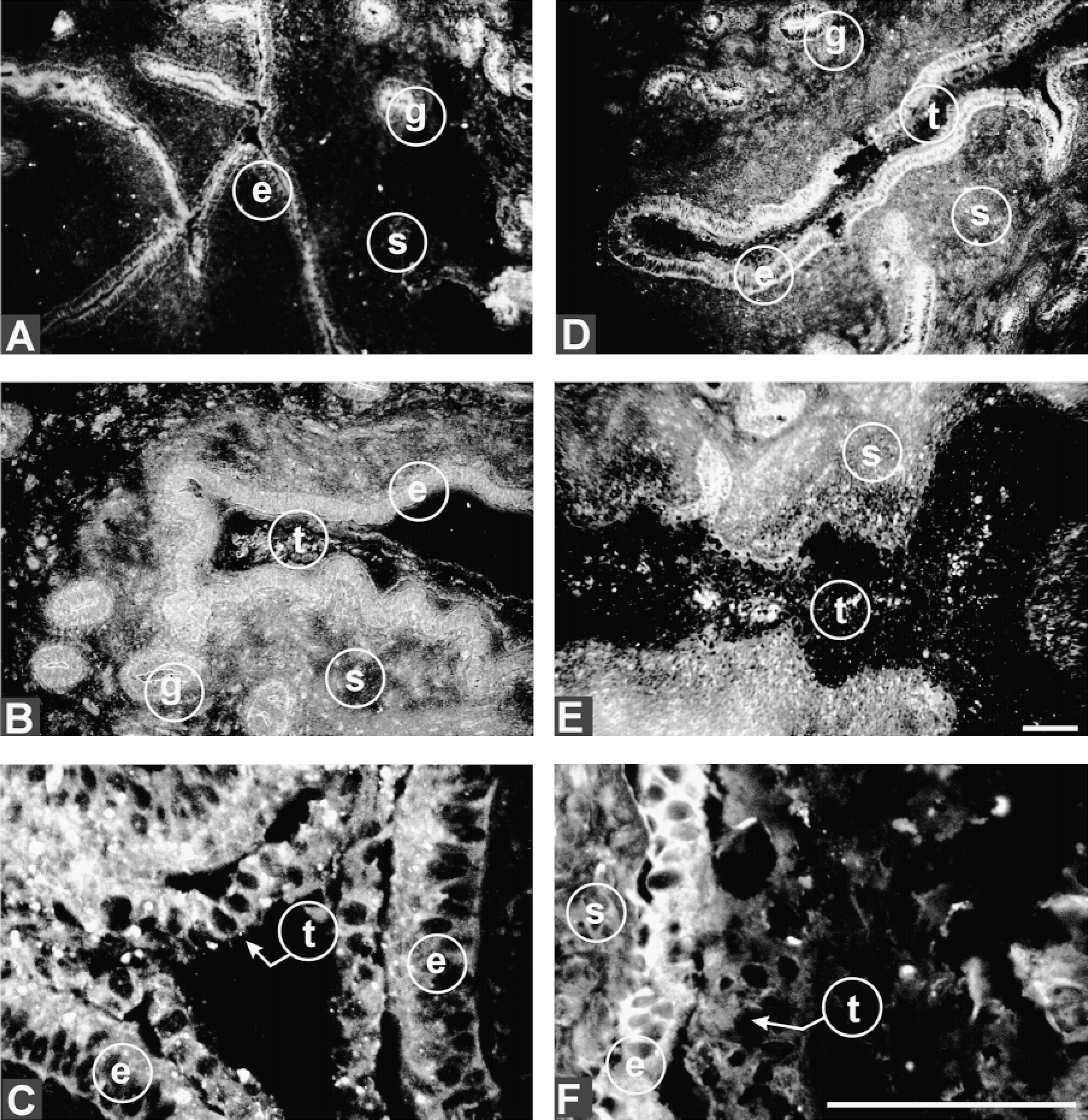
Distribution of subunit A of the V-ATPase in developing bovine trophoblast and maternal epithelium. Expression of the 70-kD V-ATPase (subunit A) in the luminal epithelium of Day 18 nonpregnant (A), Day 18 pregnant (B,C), Day 21 pregnant (D), and Day 24 pregnant (E,F) animals. t, trophoblast; e, maternal epithelium; s, stroma; g, gland. Arrows point to trophectoderm that stains more intensely for subunit A. Bars = 100 μm.
Within the trophoblast this subunit was visible throughout cells proximal to the uterine epithelium at Day 18 (Figure 2C) but by Day 24 had become diffuse, and was no longer prevalent at points of contact with endometrium (Figures 2E and 2F).
Expression of V-ATPase Subunit B in Pregnant and Nonpregnant Animals
In Day 18 nonpregnant animals, subunit B was abundant on the apical surface and lateral aspects of luminal epithelial cells and less evident on the basal surface (Figure 3A). In Day 18 pregnant animals its expression remained primarily apical. Like subunit A and ductin, by Day 21 its expression was primarily apical and basal (Figure 3C), a pattern that continued through to Day 24 (Figure 3D). Expression in glandular epithelium was, like that of the other subunits, primarily apical throughout all stages (e.g., Figures 3C and 3D). Also like subunit A, the expression of subunit B remained constant in the uterine stroma throughout the stages of pregnancy examined (Figures 3B-3D). Within the trophoblast, subunit B expression at Day 24 was also similar to that of the other subunits, becoming diffuse throughout the embryo (Figure 3D).
Specificity of Antibodies to Subunits A and B
The anti-V-ATPase subunit antibodies used in these experiments were raised against nonbovine proteins. The specificities of the anti-subunit A and anti-subunit B sera for the bovine equivalents were verified by Western blot analysis of proteins isolated from bovine kidney, an organ known to be enriched for V-ATPase. The antibodies detected uniquely proteins of the appropriate molecular weights (Figure 4).
Although the anti-ductin antibody used here does not recognize denatured proteins on Western blots, for several reasons we have no doubts about its specificity. First, ELISA data provided by Alpha Diagnostics confirmed “excellent” affinity for the peptide antigen. Second, antibodies raised against the same rodent-derived peptide specifically detect human ductin (e.g., Ohta et al. 1996; and our own observations). The bovine and human sequences spanning this peptide are in fact identical and differ from rodent by only two conservative substitutions at positions 1 and 5 (Gillespie et al. 1991; Goldstein et al. 1991), so the crossreactivity we observed with bovine material was fully anticipated.
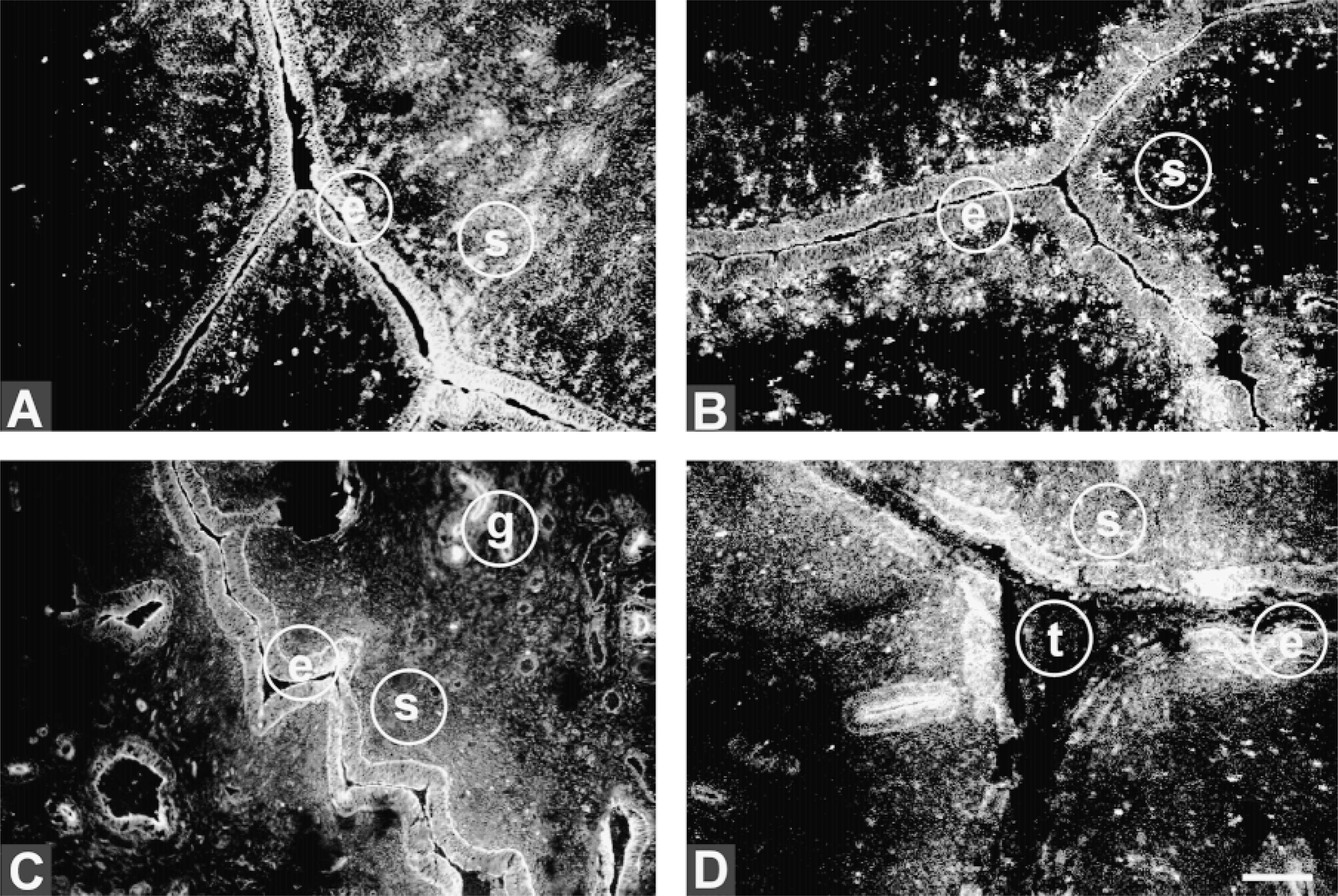
Distribution of subunit B of the V-ATPase in maternal epithelium. Sections through uteri of nonpregnant animals at Day 18 of the estrous cycle (A), Day 18 pregnant (B), Day 21 pregnant (C), and Day 24 pregnant (D). t, trophoblast; e, maternal epithelium; s, stroma; g, gland. Bar = 100 μm.
Discussion
Our study reveals that all V-ATPase subunits examined undergo a loss of preferential expression on the apical surface of luminal epithelium in response to pregnancy, suggesting that in these cells there is a shift in polarization of V-ATPase activity. The apical distribution of the V-ATPase pump seen in the luminal and glandular epithelium of the nonpregnant animal suggests that hydrogen ions are being pumped into the uterine lumen and into the lumen of the glands. After embryo attachment, this loss of polarity may reflect the requirement to reduce acidification of the fetal environment and allow clearance of H+ ions by the maternal blood supply. This shift could be important for reduction of surface charge on the luminal epithelium. The net negative charge on the epithelium has been proposed to be an electrostatic barrier to implantation (Parr and Parr 1989).
The redistribution of the V-ATPase pump could also be indicative of transcytosis of receptor-ligand complexes or embryo-derived molecules from apical to basal surfaces, because V-ATPases are required for pH regulation in vesicles involved in this process (Nelson 1992). Placental lactogen, cholesterol, and iron are examples of molecules that are believed to be delivered across epithelial cells to the stroma. Maternal IgG is also selectively transported across the yolk sac of the fetal rat and the small intestine of the neonatal rat through binding of its receptor, the Fc receptor. In both cases, this transfer of IgG occurs in a pH-dependent manner, with binding to the Fc receptor occurring only at an acidic pH of 6.0 (Rodewald and Abra hamson 1982; Rodewald and Kraehenbuhl 1984). In the small intestine there is naturally an acidic environment, and formation of the IgG-receptor complex occurs on the cell surface. Internalized complexes are then transcytosed across the cell and IgG released into the neonate, thereby evading transport to degradative lysosomes. In contrast, the yolk sac environment of the fetal rat is not acidic, and Roberts et al. (1990) propose that the binding of IgG to its receptor occurs after nonspecific engulfment of proteins into acidic endosomes. In the cow, it is possible that the shift we observed in expression of V-ATPase subunits from the apical surface to apical and basal surfaces could reflect directed transport of carrier vesicles via transcytosis.
Trophoblast cells also expressed all three subunits of the V-ATPase pump, with staining most evident in proximity to the endometrium. Although we can only speculate, it is possible that V-ATPase activity at those contact points may reflect the need for some localized hydrolysis of matrix components to allow attachment, in a manner analogous to the role of V-ATPase in macrophages and osteoclasts. By Day 24, the trophoblast and endometrium were both becoming remodeled, and precise estimation of co-localization between the trophectoderm and epithelium became more difficult.
There is a growing body of literature to suggest that ductin, either as part of the V-ATPase complex or on its own as a gap junction, has additional roles in mediating cell interactions and cell growth. Evidence for roles in growth control include the observation that the PDGF-β receptor (Goldstein et al. 1992), the E5 oncoprotein of papillomaviruses (Goldstein et al. 1991), and β1 integrin (Skinner and Wildeman in press) can form complexes with ductin, and the observation that selective inhibition of ductin by bafilomycin can suppress proliferation and induce differentiation (Tamura and Ohkuma 1991; Manabe et al. 1993; Ohkuma et al. 1993). The pronounced expression of ductin on basal aspects suggests that it could also be acting as a gap junction, allowing epithelial cells to become receptive to signaling with the stroma and vasculature.
One aspect unique to ductin is its developmental regulation in the stroma. Expression is highest at Day 18 of pregnancy, drops before Day 21, and is restored by Day 24. Implantation begins at approximately Day 18 in the bovine, suggesting that ductin could play a role in cell-cell communication events important for implantation, but that this communication is then suppressed during a critical 5-6-day window when first embryo/maternal contacts are being strengthened. Little is known about the signals that pass through ductin gap junctions and whether those signals are different from those communicated via connexins, but some unique role for ductin in implantation appears likely. Interestingly, the downregulation of ductin expression in the stroma at Day 21 parallels the pattern seen with β1 integrin (unpublished observations), a cell matrix receptor that affects many signaling pathways.
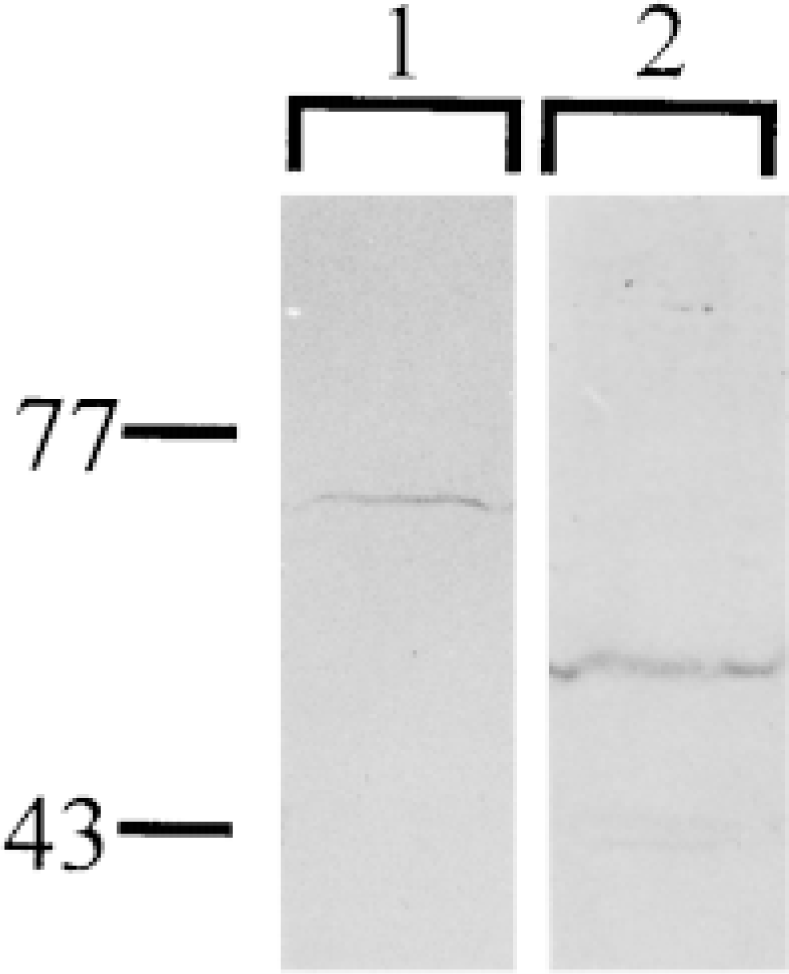
Western blot analysis of subunits A and B in bovine kidney. Lane 1, expression of subunit A; Lane 2, expression of subunit B in whole-cell lysates generated from bovine kidney.
The pregnancy-related developmental regulation of V-ATPase subunits suggests that hydrogen ion movement and resulting pH regulation play important roles in early stages of placentation. The complex roles of the 16-kD subunit of this enzyme, ductin, raise many interesting questions about the significance of its unique pregnancy-related changes in expression to ruminant implantation.
Footnotes
Acknowledgements
We thank Ms Penny Gavin, Sarah Kimmins, and Dr HaiChoo Smith for technical support in collecting uterine samples and preparing cryosections, and Dr Melissa Farquhar for technical support and advice with confocal microscopy. We also thank the staff at Brookside Abbattoir and HUB Meat Packers, Ltd., for their cooperation during tract collection. We also acknowledge the generous donation of polyclonal antibodies from Stephen L. Gluck, L. Shannon Holliday, and Shoji Ohkuma, and the donation of Lutalyse from Pharmacia-UpJohn. Chris Haworth and co-workers in the Department of Animal Science at the University of Guelph provided bovine kidney. Keith Betteridge provided valuable inputs to the discussion.