
Abstract
The human HtrA family of proteases consists of three members: HtrA1, HtrA2, and HtrA3. In bacteria, the chief role of HtrA is recognition and degradation of misfolded proteins in the periplasm, combining a dual activity of chaperone and protease. In humans, the three HtrA homologues appear to be involved in diverse functions such as cell growth, apoptosis, allergic reactions, fertilization, control of blood pressure, and blood clotting. Previous studies using RNA blot hybridization have shown that the expression of HtrA1 is ubiquitous in normal human tissues. Here we show by immunohistochemistry (IHC) that HtrA1 is widely expressed, although different tissue distributions and/or levels of expression were detected in the different tissues examined. In particular, high to medium HtrA1 expression was detected in mature layers of epidermis, in secretory breast epithelium, in liver, and in kidney tubules of cortex, in concordance with its secretory properties. Furthermore, we show a higher protein expression level in the epithelium of proliferative endometrium, in contrast to epithelium of secretory endometrium, which is almost completely negative for this protein. This suggests a possible role for HtrA1 in the modulation of tissue activity in this organ. The various expression levels in human tissues indicate several possible roles for HtrA1 in different cell types.
The bacterial serine protease HtrA, also known as DegP, is a heat shock-induced, envelope-associated serine protease (Pallen and Wren 1997). Its chief role is recognition and degradation of misfolded proteins in the periplasm, combining a dual activity of chaper-one and protease (Spiess et al. 1999; Krojer et al. 2002). Homologues of HtrA have been described in a wide range of bacteria and in eukaryotes. To date, three human homologues of HtrA have been identified and have been named, respectively, human HtrA1 (PRSS11 or L56) (Zumbrunn and Trueb 1996; Hu et al. 1998), hHtrA2 (Omi) (Faccio et al. 2000; Gray et al. 2000), and hHtrA3 (PRSP) (Nie et al. 2003a,b). The cDNA sequence of the fourth member of the family, named HtrA4, has been recently cloned (GenBank accession no. AK075205.1). These proteases appear to be involved in diverse functions such as cell growth, apoptosis, and inflammation (Clausen et al. 2002).
HtrA1 is the first identified member of the human HtrA protein family. It is a secreted protein, shown to be upregulated during osteoarthritis or to be significantly downregulated in transformed fibroblasts, in ovarian cancer, and in melanoma (Zumbrunn and Trueb 1996; Hu et al. 1998; Baldi et al. 2002,2003; Shirdar et al. 2002). Moreover, its overexpression inhibits cell growth and proliferation in vitro and in vivo, thus suggesting a possible role as a tumor suppressor (Baldi et al. 2002). Finally, HtrA1 is upregulated in human skeletal muscle in Duchenne muscular distrophy (Bakay et al. 2002).
HtrA2, initially identified as a stress-activated protease, is considered to be a member of the apoptotic machinery that binds and therefore regulates the function of the inhibitors of apoptotic proteins (IAPs) (van Loo et al. 2002; Verhagen et al. 2002). This action results in the elimination of the caspase-inhibitory activity of IAPs and thus in the promotion of cell death. Interestingly, to acquire the ability to bind IAPs, HtrA2 is first imported into mitochondria and is processed to a truncated form that lacks the amino-terminal domain.
In contrast to HtrA1 and HtrA2, the precise function of HtrA3 is still largely unknown. Given its higher homology with HtrA1, it is speculated that it could have similar functions. This hypothesis is strengthened by the observation that mRNAs coding for the two proteases display a similar RNA expression pattern in human tissues (Nie et al. 2003a,b).
Here we describe a study of the expression pattern of HtrA1 in adult human tissues conducted by imunohistochemistry (IHC).
Materials and Methods
Normal Tissues
Normal tissues from autopsies were obtained from the Section of Anatomic Pathology of the Second University of Naples, Italy. Tissues were formalin-fixed and paraffin-embedded. Representative sections of each specimen were stained with hematoxylin-eosin and were examined by a pathologist to confirm the histological preservation of the microanatomic structure. For each tissue examined, at least three specimens from two different individuals were analyzed. A human tissue array (AA4 Normal organs; Superbiochips, Korea), containing 59 normal human adult tissues, was also used.
Immunohistochemistry
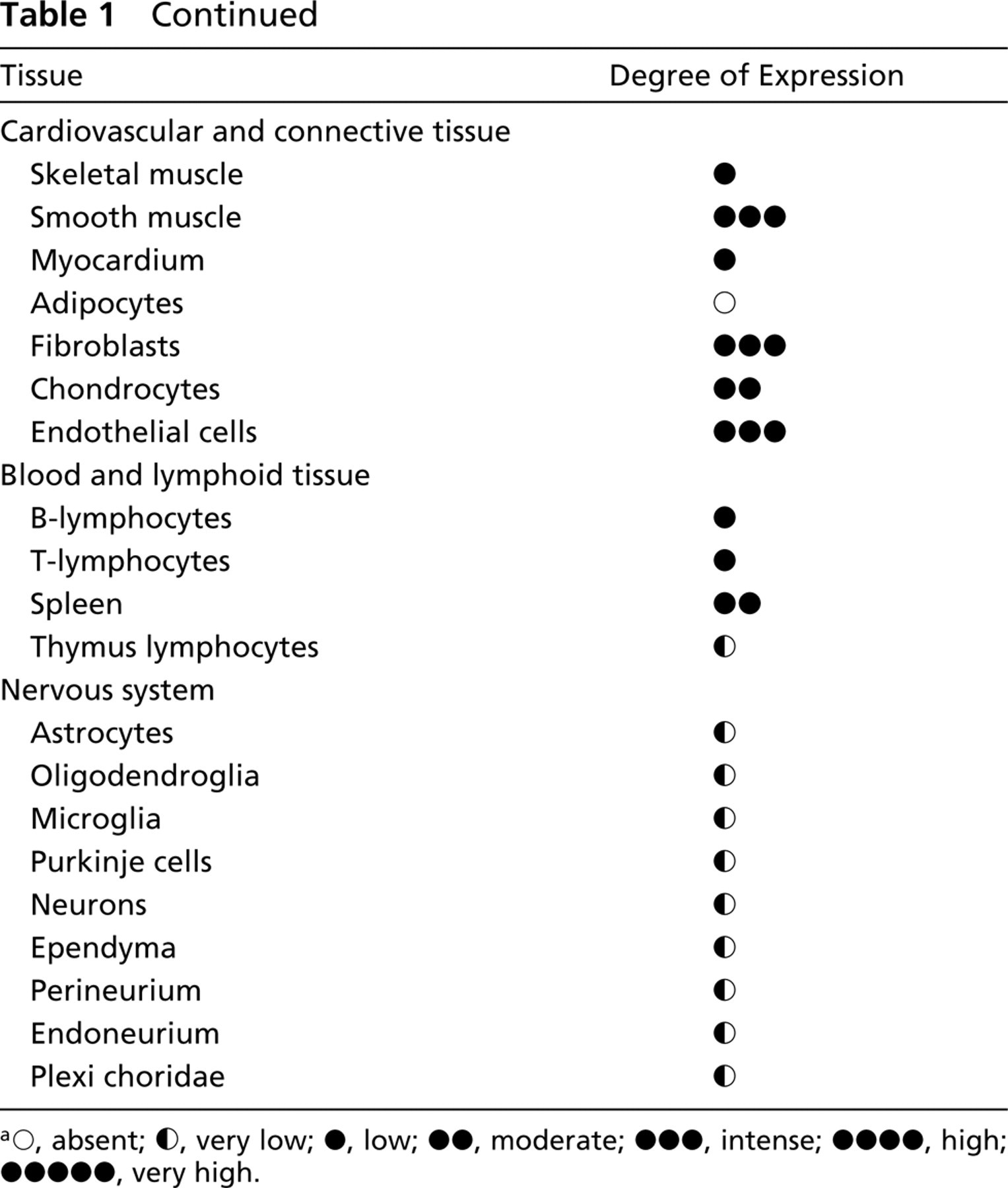
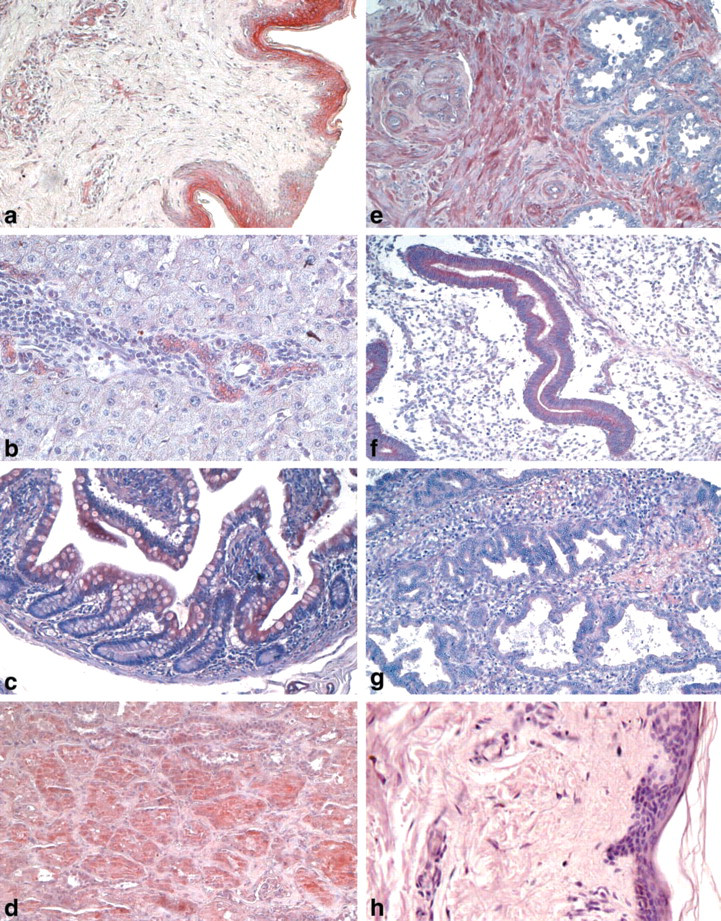
IHC was carried out essentially as described previously (Baldi et al. 2002). Briefly, sections were deparaffinized in xylene, rehydrated through a graded alcohol series, and washed in PBS. PBS was used for all subsequent washes and for antiserum dilution. Tissue sections were quenched sequentially in 3% H2O2 and blocked with PBS-6% nonfat dry milk (Biorad; Hercules, CA) for 1 hr at room temperature. Slides were then incubated at 4C overnight with the rabbit polyclonal immune serum raised against HtrA1 (Baldi et al. 2002) at a 1:10 dilution and then with diluted anti-rabbit biotinylated antibody (Vector Laboratories; Burlingame, CA) for 1 hr. All slides then were processed by the ABC method (Vector Laboratories) for 30 min at RT. Novared (Vector Laboratories) was used as the final chromogen and hematoxylin was used as the nuclear counterstain. Negative controls for each tissue section were prepared by substituting the primary antiserum with the respective preimmune serum (Figure 1 h). All samples were processed under the same conditions. The expression level of HtrA1-stained cells per field (×250) at light microscopy was calculated and compared in different specimens by two separate observers (A.B. and F.B.) in a double-blind fashion and described as absent (∘), very low (∘), low (•), moderate (••), intense (•••), or a very high (••••). Noticeable quantitative variations in HrtA1 expression in staining intensity were observed, with less variation in the total number of stained cells.
Results
We used IHC to determine the amount and localization of HtrA1 in a panel of different human tissues. HtrA1 was expressed widely, although a different tissue distribution and/or level of expression was detected in the different organs examined (Table 1). Noticeable quantitative variations in HrtA1 expression were observed in staining intensity, with less variation in the total number of stained cells.
Expression of HtrA1 in Epithelial Tissues
Epithelial cells from either simple or stratified epithelium showed variable positive staining for HtrA1. Stratified epithelia, such as those from esophagus, cervix, and vagina, expressed a low level of this protein with a major localization in the mature and outer layers, with respect to basal layers that showed an undetectable level of the protein. In contrast, in the skin there was a high expression level for HtrA1 in mature layers of epidermis, whereas a low expression level was found in hair follicles, sebaceous glands, and sweat glands (Figure 1a). A medium expression level was seen in secretory breast epithelium compared to a low expression level in breast duct epithelium. Low immunoreactivity was detected in the stratified columnar epithelia of trachea, bronchi, and adjacent glands. Pneumocytes showed an undetectable level of the protein.
In the gastrointestinal system, low positive cytoplasmic staining for HtrA1 was found in secretory ducts, compared with an undetectable level in acini of salivary glands. An intense level was shown in the duct cells of the liver, whereas a low level was seen in the hepatocytes (Figure 1b). Low-level HtrA1 expression was seen in the stomach and gallbladder, and a moderate level was seen in the colon epithelia (Figure 1c). The exocrine portion of the pancreas showed a moderate level of expression for HtrA1, whereas the endocrine portion of the gland was less intense.
In the urinary system, HtrA1 was expressed at a medium level in all kidney tubules of cortex (Figure 1d), whereas a low expression of the protein was shown in the tubules of medulla and in the glomeruli. Uroepithelium and prostate epithelia were negative for HtrA1, whereas prostate stroma showed moderate positivity for this protein.
HtrA1 was expressed at a medium level in thyroid follicles. Its expression was low in the adrenal glands, where staining was more intense in the glomerulosa than in reticularis and fascicolata regions of the cortical portion, in contrast to an undetectable level of HtrA1 showed in the chromaffin portion of the glands.
HtrA1 protein expression in normal adult human tissues

∘, absent; ∘, very low; •, low; ••, moderate; •••, intense; ••••, high; •••••, very high.
The male reproductive system displayed low to undetectable positive cytoplasmic staining for HtrA1 in the epithelia. Low levels of expression were found in epididymis, in contrast with an undetectable level in testis and in epithelium of the seminal vesicle (Figure 1e), where only the stroma showed intermediate positivity for the protein.
Very interesting was the pattern of expression of HtrA1 in the female reproductive system. HtrA1 was expressed at an intermediate level, particularly in the epithelium of proliferating endometrium (Figure 1f), in contrast to epithelium of secretory endometrium, which was almost completely negative for this protein (Figure 1g). In the exocervix, an intermediate level of HtrA1 was found in the basal layer in contrast with the outer and mature layers, which showed a lower level of the protein. In the ovary, only the stroma showed an intermediate level of expression of HtrA1, whereas both oogonia and follicles showed a low level of the protein. Finally, greater expression of HtrA1 was found in third trimester placenta compared with placenta at the first trimester of gestation.
(
Expression of HtrA1 in Cardiovascular and Connective Tissues
Low immunoreactivity for HtrA1 was observed in skeletal muscle cells and in myocardial cells. An undetectable level was found in adipocytes, whereas a medium level was shown in chondrocytes, endothelial cells, and fibroblasts.
Expression of HtrA1 in Central and Peripheral Nervous System
All neurons from different areas of the brain, such as frontal cortex and midbrain and cells of the granular level of the cerebellum, displayed low staining for HtrA1. Perineural and endoneural cells of peripheral nerves and ganglion cells, such as astrocytes, oligodendroglial, and microglial cells of brain tissue, showed low levels of the protein.
Expression of HtrA1 in Hematopoietic Cells
An undetectable/low level of HtrA1 was observed in blood cells and in several lymphoid tissues.
Discussion
In contrast to HtrA2, the precise function of HtrA1 is still largely unknown. It is reported to be a secreted protein (Gray et al. 2000), and it has been suggested to be involved in the degradation of extracellular matrix proteins important for both arthritis and tumor progression and invasion (Clausen et al. 2002). The HtrA1 gene is expressed in many human tissues (Nie et al. 2003a,b). The recent development of a polyclonal antibody against hHtrA1 has contributed to defining the presence of this protein in some cell lines (Baldi et al. 2002). To the best of our knowledge, this is the first report describing the IHC localization and distribution of HtrA1 in human tissues.
An exhaustive knowledge of the differential cell- and tissue-specific pattern of expression of HtrA1 in normal human tissues is an essential requirement for a critical evaluation of the exact role played by this protein in cell homeostasis, especially in view of the fact that this serine protease is downmodulated in several human tumors, such as ovarian tumors (Shirdar et al. 2002) and melanoma (Baldi et al. 2002,2003). When we examine the IHC pattern of expression of HtrA1 in several human tissues, we can conclude that HtrA1 has a widespread pattern of expression that is consistent with previous data on the RNA expression profile for this protein in human tissues (Zumbrunn and Trueb 1996; Nie et al. 2003a,b). However, we found a different tissue distribution and/or level of expression in several organs. In particular, high to medium HtrA1 expression was detected in mature layers of epidermis, in the secretory breast, and in liver epithelium. This particular localization is consistent with a secreted protein. Furthermore, the peculiar distribution of HtrA1 in hepatic duct cells suggests its involvement in their proliferation. In fact, hepatic duct cells are particularly sensitive to growth-factor induction (Nagy et al. 1996).
We have shown a very particular expression of this serine protease in the endometrium. High protein expression is detected in the glandular epithelium of proliferating endometrium, in contrast to secretory endometrium that appeared almost completely negative for this protein. This peculiar pattern of expression of HtrA1 might account for distinct activities of this protein in this tissue. Intriguingly, it has been proposed that possible substrates of interest of HtrA1 include other proteases, extracellular matrix proteins, growth factors, and proteins that modulate growth factors (Hu et al. 1998). It is well known that basement membranes, which are elements of the extracellular matrix, play an important role in proliferation and differentiation of human endometrium and that their degradation influences estrogen transport from blood to stroma (Bulletti et al. 1991). Therefore, the increase in endometrial HtrA1 expression during the proliferating phase of endometrium suggests a possible role for this protein in preparation of the endometrium lining for implantation. In addition, it has been demonstrated that levels several serine protease inhibitors, such as placental protein 5 (Butzow et al. 1986) or bikunin (Hettinger et al. 2001), are higher in the secretory phase than in the proliferating phase. This suggests that HtrA1 is involved in the delicate balance between serine proteases and their inhibitors in the control of endometrial physiology. Consistent with a possible role of HtrA1 in placentation, the expression of this protease was dramatically elevated in the third-trimester placenta. This behavior is similar to that shown for HtrA3 (Nie et al. 2003a,b). We are investigating the expression of HtrA1 in a large number of first- and third-trimester placentas, with the aim of investigating the role of this protease in placentation.
In conclusion, our data concerning the IHC distribution of HtrA1 in normal human tissues demonstrate that the expression of this protease is modulated not only in pathological conditions but also in tissues with different physiological activities. Further studies aimed at investigating the transcription regulation of HtrA1 gene and at identifying functional targets of its protease activity will help to clarify the exact role of this protein in both physiological and pathological conditions.
Footnotes
Acknowledgements
Supported in part by the AIRC and Ministero della Salute grants to M.G.P., by Futura-ONLUS and by Second University grants to A.B. We thank the International Society for the Study of Comparative Oncology for its continuous support.
We also thank Mr Giuseppe Falcone for his contribution to the image elaboration.