
Abstract
Localization of the MCC (mutated in colorectal cancer) gene product, a cell cycle-regulating protein mutated in several colorectal tumors, in various mouse tissues was examined by immunohistochemistry and immunoelectron microscopy. MCC was localized on microvilli and in the apical cytoplasm in renal proximal tubule epithelial cells and pancreatic acinar cells. In hepatocytes, MCC was exclusively detected on microvilli. MCC was highly expressed in the cerebral cortex and the molecular layer of the cerebellar cortex and was partially associated with membrane organelles in neuronal elements. Adrenal chromaffin cells showed little expression of MCC. MCC was localized to the cell margins of ependymal cells, thyroid follicular cells, and anterior pituitary cells. In parotid acinar cells, only the apical surface was immunopositive. MCC was not expressed in skeletal and cardiac muscle. MCC was present at lateral cell borders in the duodenum and colon epithelium. In addition, the apical cytoplasm of colon epithelial cells exhibited intense immunoreactivity. The amount of MCC increased during differentiation of NGF-treated PC12 cells. In conclusion, MCC was expressed in differentiated cells and was associated with the plasma membrane and membrane organelles. In addition to the negative regulation of the cell cycle, MCC may be involved in cell differentiation.
Keywords
T
The product encoded by the MCC gene is predicted to be composed of 829 amino acids and has little homology to other known proteins (Kinzler et al. 1991a). The MCC protein uniquely contains several regions that have a high probability of forming a coiled-coil structure. We previously found that MCC is a membrane-associated cytoplasmic phosphoprotein with a molecular weight of 100 kD, and its overexpression blocks cell cycle progression from the G0/G1 to the S-phase (Matsumine et al. 1996b; Senda et al. 1997). This suggests that MCC plays a role in the signaling pathway that negatively regulates cell cycle progression. MCC was shown to be highly expressed in the cerebellar cortex, apical villi of the small intestine, and surface epithelium of the colon, all of which are composed of differentiated cells (Matsumine et al. 1996b; Senda et al. 1997). These results raise the possibility that MCC is involved in cell differentiation. In this regard, it is important to determine the expression and subcellular localization of MCC in other murine tissues.
In this study we examined the expression and localization of MCC in various tissues of mice by fluorescence immunohistochemistry and immunogold electron microscopy. In addition to asking about the relationship between MCC and cell differentiation, we analyzed MCC expression during nerve growth factor-mediated differentiation of PC12 cells.
Materials and Methods
Antibody
Anti-MCC antibody was prepared by immunizing rabbits with synthetic peptides representing amino acid residues 817-829 (Matsumine et al. 1996b). Specific antibody was purified by affinity chromatography using a column to which the synthetic peptide had been linked. The specificity of the antibody was checked by immunoprecipitation (Matsumine et al. 1996b).
Immunofluorescence Microscopy
Normal 8-week-old male ddY mice (n = 10) were used for this study according to the guidelines of Nagoya University School of Medicine on the use and care of animals. Seven mice were perfused with 3% paraformaldehyde (Merck; Darmstadt, Germany)/0.1 M phosphate buffer (pH 7.4) through the left cardiac ventricle for 5 min. Kidney, liver, pancreas, anterior pituitary, thyroid gland, parotid gland, psoas muscle, heart, cerebrum, cerebellum, adrenal gland, duodenum, and distal colon were resected and immersed in the same fixative for 2 hr. The fixed tissues were then washed thoroughly in graded concentrations (10, 15, and 20%) of sucrose/PBS, embedded in OCT compound (Miles; Elkhart, IN), and frozen quickly with liquid nitrogen. Six-μm-thick frozen sections were cut with a cryostat (CM-41; Sakura Seiki, Tokyo, Japan), and incubated with the anti-MCC antibody diluted 1:100 with PBS at 4C overnight. After washing in PBS, the sections were incubated with fluorescein isothiocyanate (FITC)-conjugated goat anti-rabbit IgG antibody (Seikagaku Kogyo; Tokyo, Japan) diluted 1:100 with PBS at room temperature (RT) for 1 hr. The sections were washed in PBS, mounted in 90% glycerol containing 1 mg/ml p-phenylenediamine as an antifade, and examined with a fluorescence microscope (Fluophoto: Nikon, Tokyo, Japan). Control sections were incubated with the anti-MCC antibody preabsorbed with the recombinant MCC protein used as antigen. The immunohistochemical staining was repeated several times and the same results were obtained from all the tests.
Immunoelectron Microscopy
Lowicryl K4M ultrathin sections of kidney, liver, pancreas, cerebral cortex, and distal colon of three mice were prepared according to the method of Senda et al. (1991). The Lowicryl thin sections of these tissues were incubated with the anti-MCC antibody diluted 1:100 with PBS at RT for 2 hr. After washing in PBS, the sections were incubated with 10-nm colloidal gold-conjugated goat anti-rabbit IgG antibody (Amersham Japan; Tokyo, Japan) diluted 1:30 with PBS at RT for 1 hr. The sections were stained with uranyl acetate and lead citrate and examined with an electron microscope (H-7100: Hitachi, Tokyo, Japan). Control sections were incubated with the anti-MCC antibody preabsorbed with the antigen protein. The immunoelectron microscopic analysis was repeated several times and the same results were obtained from all the tests.
Cell Culture and Western Blot Analysis
The rat pheochromocytoma cell line PC12 is known to be differentiated into a neuronal cell line by treatment with nerve growth factor (NGF) and is therefore widely utilized as a cell model for neuronal differentiation. PC12 cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal calf serum. The cells plated in a 60-mm culture dish coated with Type 1 collagen were cultured in DMEM containing 10% fetal calf serum and 5% horse serum. For induction of differentiation, PC12 cells were treated with 50 ng/ml of mouse 2.5S-NGF (Wako; Tokyo, Japan). For Western blot analysis, the cell lysates were resolved on an 8% SDS-polyacrylamide gel and transferred to a polyvinylidene difluoride membrane filter (Immobilon-P: Millipore, Tokyo, Japan). The filters were incubated with the anti-MCC antibody and subsequently with alkaline phosphatase-conjugated mouse anti-rabbit IgG antibody (Promega: Tokyo, Japan). Alkaline phosphatase activity was developed according to the manufacturer's specifications.
Results
In the kidney, MCC was expressed in the proximal urinary tubules in the cortex and the outer portion of the medulla (Figures 1A and 1B). Glomeruli were immunonegative for MCC (Figure 1C). Observation at higher magnification showed that the apical surface of the proximal tubule epithelial cells was strongly immunostained (Figure 1D). Immunoreactivity was not seen in control sections incubated with the antigen-preabsorbed antibody (Figure 1E). By immunoelectron microscopy, gold particles were distributed on microvilli and in the apical cytoplasm of the epithelial cells of the proximal urinary tubules (Figure 1F). Few gold particles were seen in control sections incubated with the antigen-preabsorbed antibody (Figure 1G).
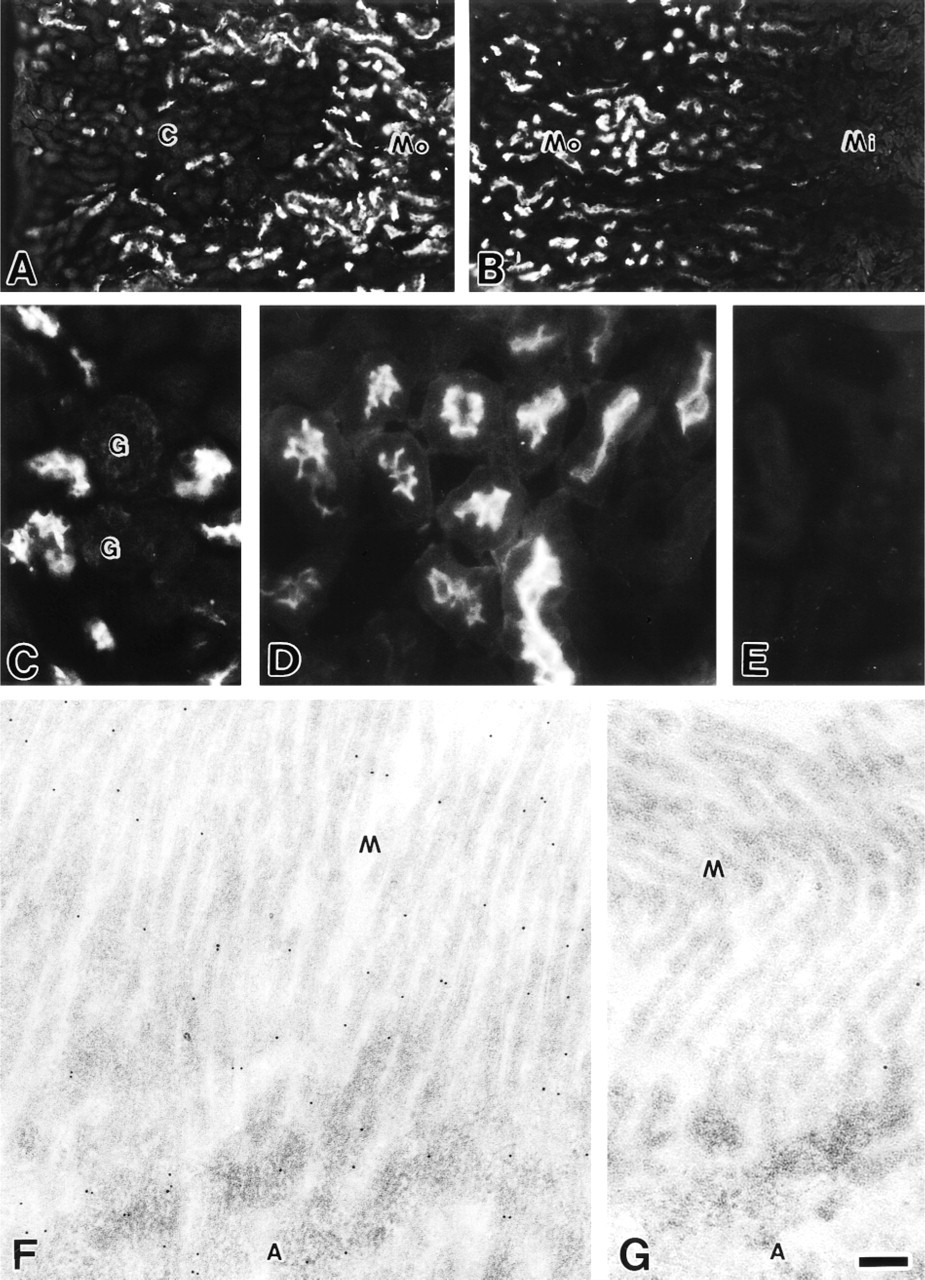
Immunofluorescence (
Strong immunoreactivity of MCC was observed along bile canaliculi between hepatocytes throughout the mouse liver (Figure 2A). Immunoelectron microscopy revealed that in hepatocytes MCC was localized exclusively on microvilli protruding into the bile canaliculus (Figure 2G). In the pancreas, immunofluorescence of MCC was shown at the lumina and intercellular canaliculi of the exocrine acini (Figure 2B). Endocrine cells in Langerhans islets were negative (not shown). By electron microscopic immunocytochemistry, gold particles were present on microvilli and in the apical cytoplasm of the pancreatic acinar cells (Figure 2H). MCC was restricted to cell margins of the ependymal cells (Figure 2C), anterior pituitary cells (Figure 2D), thyroid follicular epithelial cells (Figure 2E), and parotid acinar cells (Figure 2F). Intense immunofluorescence of MCC was detected at the lateral cell borders of the ependymal cells (Figure 2C). In anterior pituitary endocrine cells, the entire cell margin was immunopositive (Figure 2D). Whereas MCC immunoreactivity was more intense at the basal margin than at the apical margin in thyroid follicular epithelial cells (Figure 2E), only the apical margin facing the acinar lumen showed MCC reactivity in parotid acinar cells (Figure 2F). The psoas and cardiac muscles exhibited no MCC expression (not shown). The fluorescence and gold particles in these tissues and cells were completely absorbed in control sections (not shown).
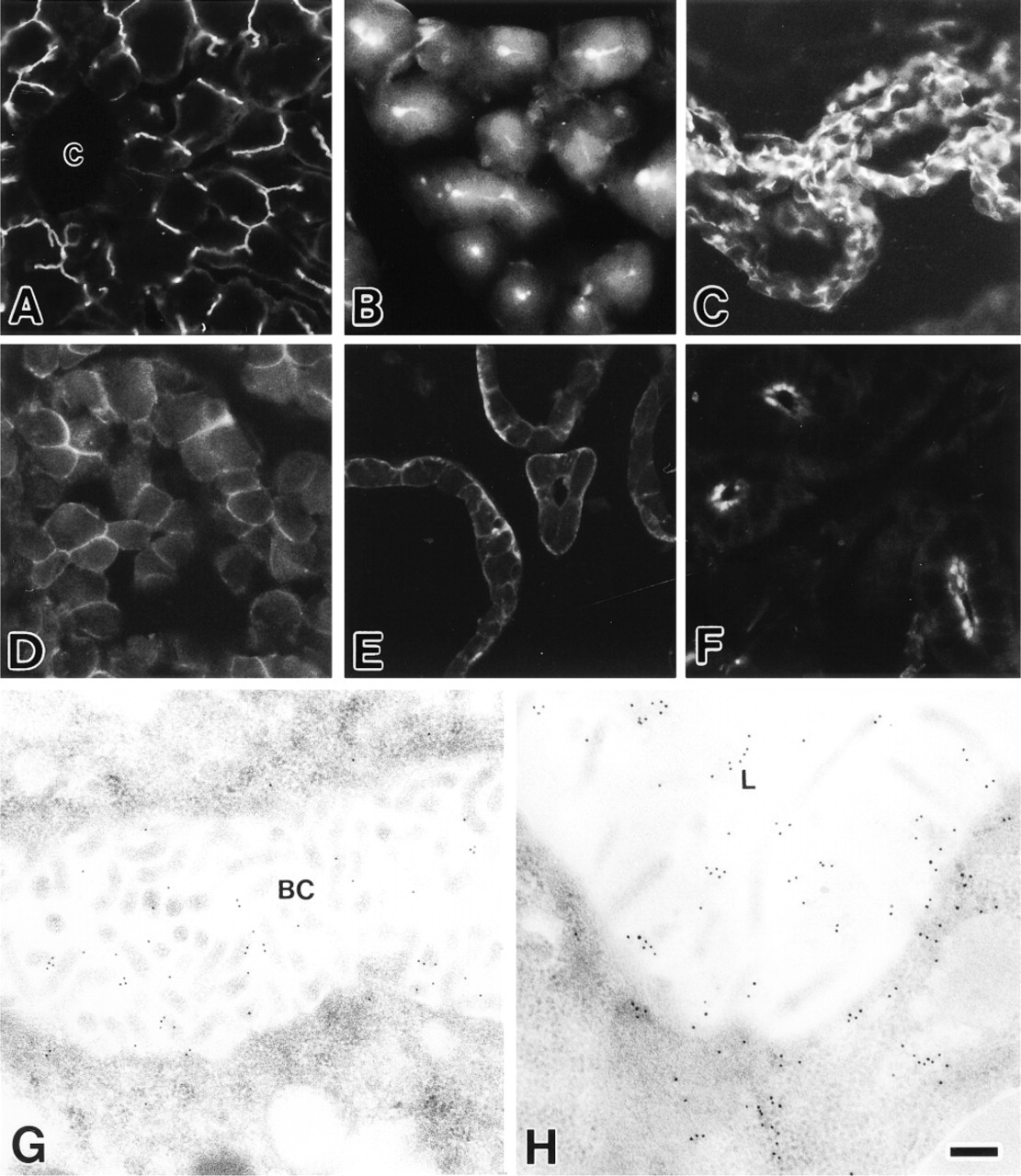
Immunofluorescence (
MCC was highly expressed in the cerebral cortex (Figure 3A) and the molecular layer of the cerebellar cortex (Figure 3B). At higher magnification, the cell bodies of both, except for nuclei, and nerve fibers of the cerebral and cerebellar cortex neurons were full of fine, granular MCC immunoreactivity (Figures 3C and 3D). By immunoelectron microscopy, gold particles were distributed widely in neuronal elements, such as cell bodies, nerve fibers, and nerve terminals (Figure 3E). Significant deposits of the gold particles were associated with membrane organelles, particularly mitochondria (Figure 3E). Expression of MCC in the adrenal medulla was much lower than in the cerebral and cerebellar cortices (Figure 3F).
MCC was highly expressed in the epithelia of the duodenum and colon and was localized at the cell margins of the epithelial cells (Figures 4A-4C). The lateral cell borders were immunostained strongly in both duodenum (Figure 4A) and colon (Figure 4B) epithelia. MCC immunoreactivity was more intense in the apical region of the intestinal villi than in the lower region (Figure 4A) and in the surface colon epithelium than in the crypts (Figure 4B). By immunoelectron microscopy, gold particles were preferentially distributed on microvilli and in the apical and lateral cytoplasm of colon epithelial cells (Figure 4C).
To investigate the regulation of MCC expression during cell differentiation, we performed Western blotting analysis with equal amounts of cell proteins extracted from PC12 cells at various time points after the treatment with NGF. PC12 cells treated with NGF showed phenotypic changes such as extension of cell processes, becoming neuron-like (not shown). The amount of MCC gradually increased after the treatment with NGF (Figure 5). The changes in the amount of MCC paralleled the time course of PC12 phenotypic changes (not shown).
Discussion
According to the predicted amino acid sequence of the MCC gene product, MCC consists of 829 amino acids, and does not have any obvious membrane-spanning region or nuclear localization signals (Kinzler et al. 1991a). In our previous study, cell fractionation experiments showed that most of the MCC proteins present in the cytosol, i.e., the 100,000 × g supernatant fraction, but a proportion of MCC was also detected in the crude membrane fraction (Matsumine et al. 1996b). Immunoelectron microscopic analysis showed that MCC is associated with the plasma membrane and membrane organelles in both native and MCC-overexpressing cells (Senda et al. 1997). In this study, we examined the expression and subcellular localization of MCC in various mouse tissues by fluorescence immunohistochemistry and immunogold electron microscopy, and demonstrated that MCC is mainly localized at the cell margins in most of the cell types examined. In cerebral and cerebellar cortex neurons in which MCC was highly expressed, MCC tended to be associated with membrane organelles. These results suggest that MCC is not a transmembrane protein but a cytosolic protein, and that a proportion of MCC localized in the peripheral cytoplasm may be linked to the plasma membrane. MCC in the neurons appears to be associated rather with membrane organelles.
It is of interest to know how MCC binds to the membranes. Because the MCC protein detected in the insoluble crude membrane fraction was not solubilized with detergents (Matsumine et al. 1996b), MCC is assumed to be complexed in an insoluble aggregate. This is consistent with the fact that MCC contains heptad repeats throughout almost the entire length of the molecule (Kinzler et al. 1991a). Therefore, MCC may bind to the membranes via cytoskeletal components. The plasma membrane is known to be under-coated with fuzzy filamentous components called the “membrane skeleton,” and the cytoskeletal system, including actin microfilaments and intermediate filaments, terminates at and binds to the membrane skeleton (Bennett 1985; Marchesi 1985). Another finding obtained in the present study, that a large amount of MCC is localized in the apical terminal web region of the colon epithelial cells, further supports this possibility. The terminal web is known to be composed of abundant cytoskeletal elements, such as rootlets of microfilament cores extending from microvilli, cross-linker systems between them, and underlying intermediate filaments (Mooseker 1985). The mechanism of the MCC binding to cytoskeletal components is an intriguing question to be determined in the future.
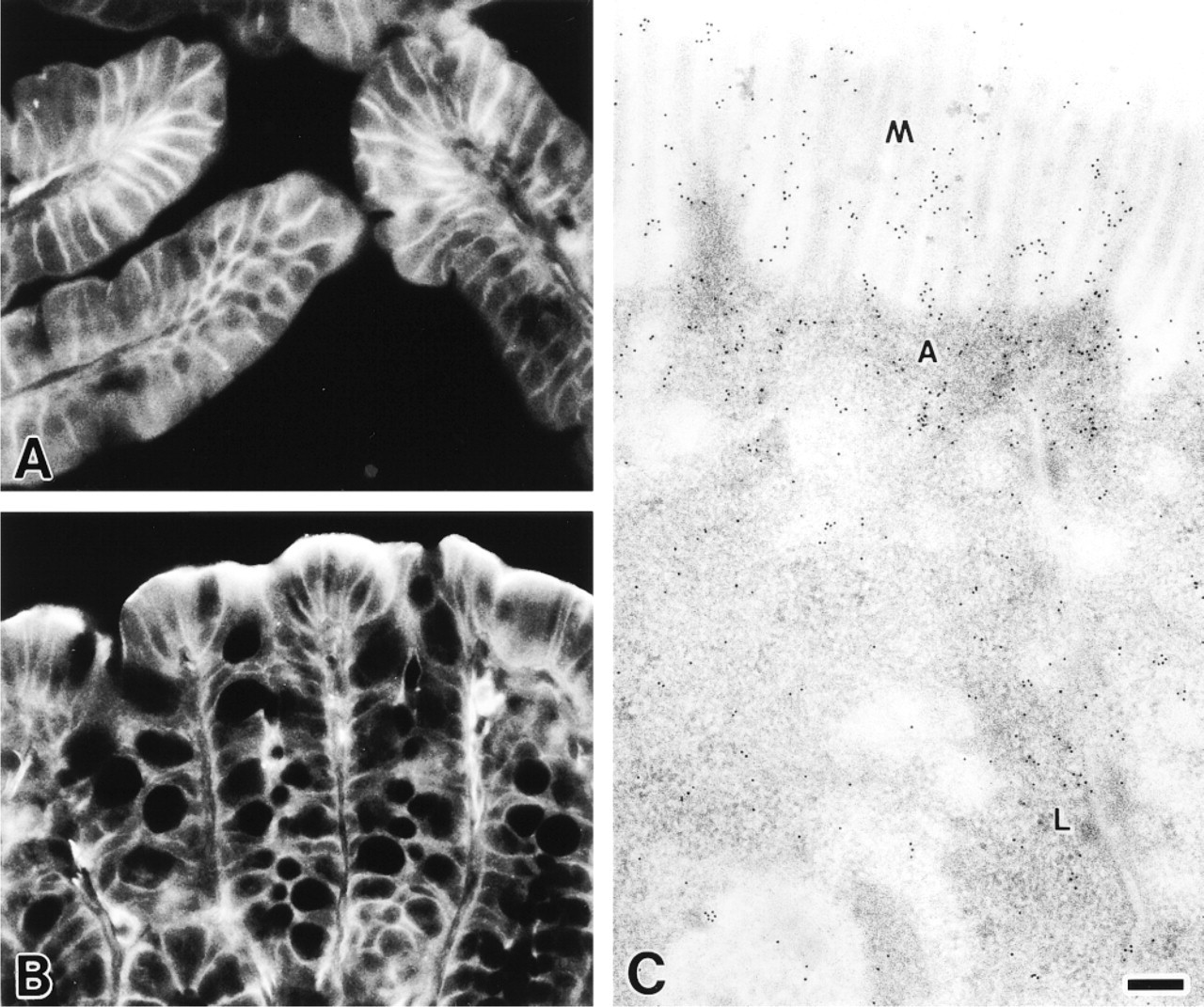
Immunofluorescence (
We have found that overexpression of MCC blocks cell cycle progression from the G1- to the S-phase and that its phosphorylation state changes markedly in a cell cycle-dependent manner (Matsumine et al. 1996b). These results suggest that MCC may negatively regulate cell proliferation and that the function of MCC is regulated by phosphorylation. In this study MCC was found to be highly expressed in differentiated cells. In particular, high levels of MCC expression were shown in cerebral and cerebellar cortex neurons, highly differentiated cells that have lost mitotic ability. Furthermore, MCC expression increased in PC12 cells after treatment with NGF, which induced differentiation of the PC12 cells into neuron-like cells with cell processes. This is consistent with the finding that MCC expression in adrenal medulla is much lower than in the cerebral and cerebellar cortices. Therefore, we speculate that MCC may be involved in cell differentiation by blocking cell cycle progression.
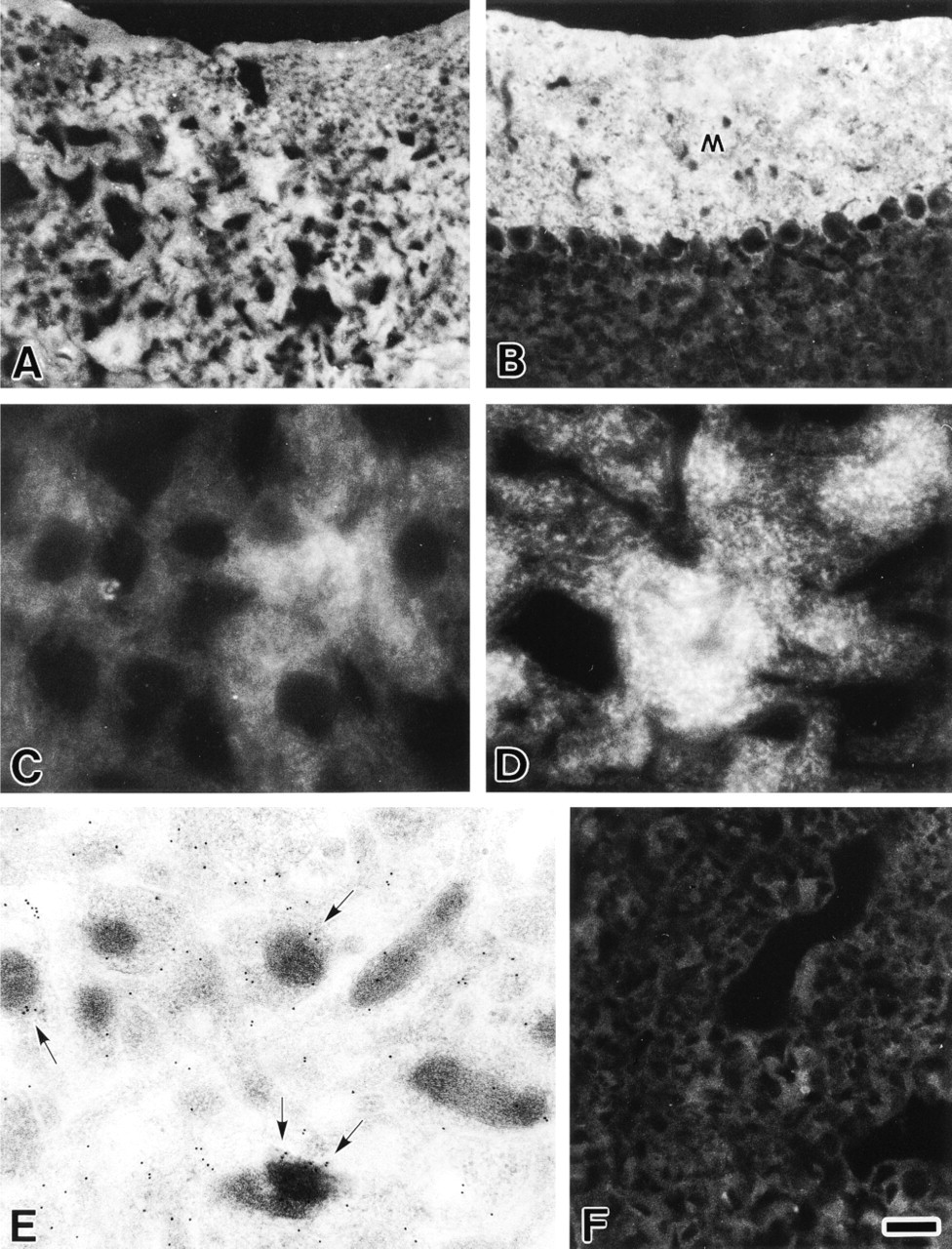
Immunofluorescence (
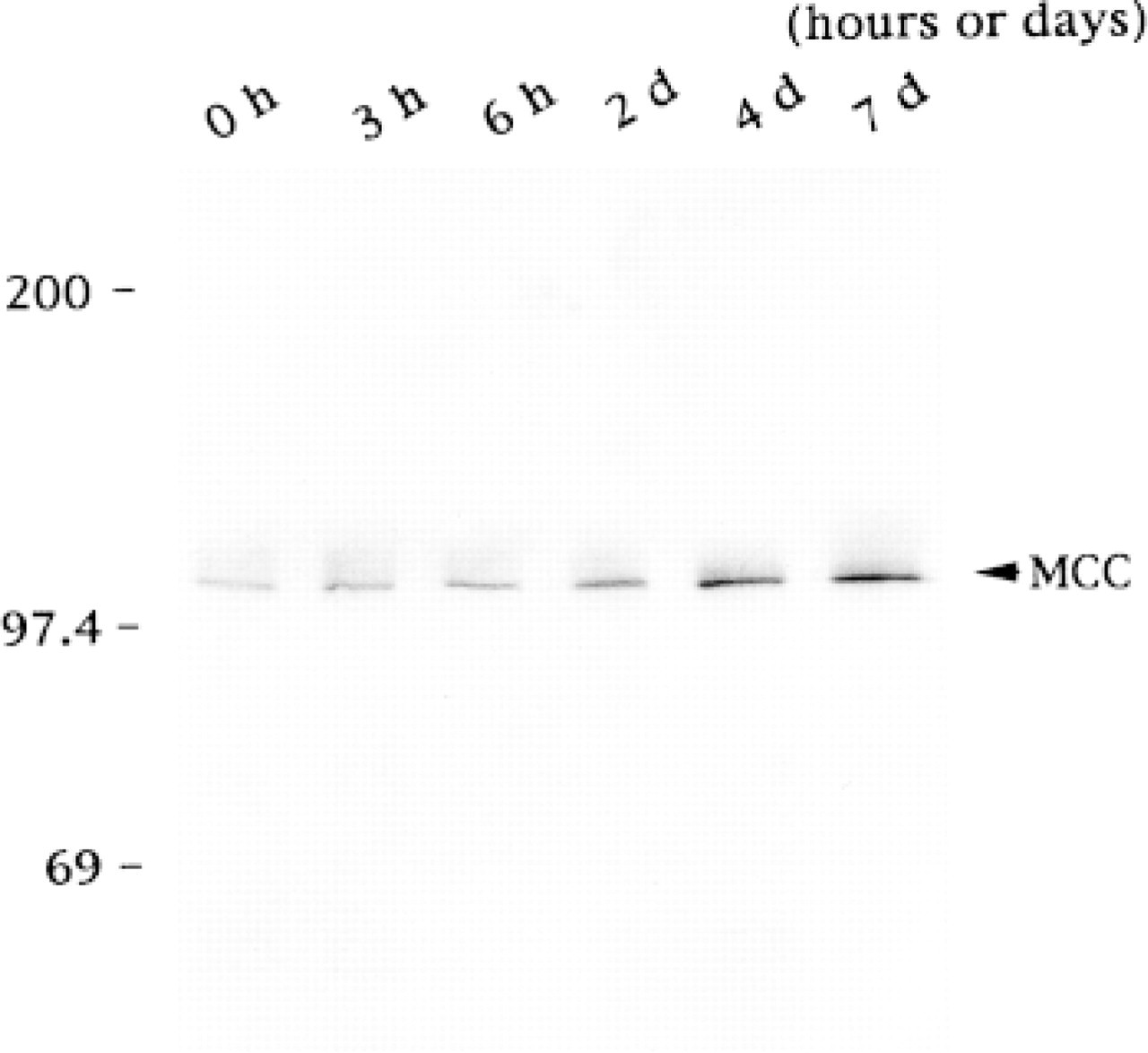
Expression of MCC during differentiation of PC12 cells treated with NGF. PC12 cells cultured in DMEM containing 10% fetal calf serum and 5% horse serum were treated with 50 ng/ml of mouse NGF. For Western blot analysis, the cell lysates were resolved on an 8% SDS-polyacrylamide gel and transferred to a polyvinylidene difluoride membrane filter. The filters were incubated with the anti-MCC antibody and then with alkaline phosphatase-conjugated mouse anti-rabbit IgG antibody. Alkaline phosphatase activity was developed according to the manufacturer's specifications. The PC12 cells treated with NGF showed the phenotypic change such as extension of cell processes (not shown). Numbers at left indicate molecular weight. The amount of MCC gradually increases after the treatment with NGF.
Because MCC is localized in the cytoplasm but not in the nucleus, signals elicited by MCC may be transduced into the nucleus through a certain signaling pathway leading to the cell cycle regulatory system. MCC contains a region possessing an amino acid similarity to the G-protein-coupled m3 muscarinic acetylcholine receptor (Kinzler et al. 1991a). In our previous study, MCC with mutations in this region did not exhibit any cell cycle-blocking activity (Matsumine et al. 1996b), suggesting that this region of MCC may play an important role in regulating cell cycle progression. However, little is known about the MCC-related signaling pathway, and no MCC binding protein has been identified. In contrast, the signaling pathway of APC, which also regulates cell cycle progression (Baeg et al. 1995), has been investigated intensively in recent years and several APC binding proteins have been identified: β-catenin (Rubinfeld et al. 1993; Su et al. 1993; Shibata et al. 1994), EB1 (Su et al. 1995) and hDLG (Matsumine et al. 1996a). APC is reported to be present not only in the cytoplasm but also in the nucleus in cultured cells (Wong et al. 1996; Neufeld and White 1997). Identification of MCC binding proteins will give new insight into the signaling pathway through which MCC regulates cell cycle progression.
Footnotes
Acknowledgements
Supported by grants from the Ministry of Education, Science, Sports and Culture, Japan and by the Aichi Cancer Research Foundation.