
Abstract
We studied the distribution of the four human apomucins MUC1, MUC2, MUC4, and MUC5AC in hyperplastic polyps, serrated adenomas, and traditional adenomas of the colorectum using immunohistochemical techniques, with the aim of comparing and contrasting their patterns of expression. A series of 12 hyperplastic polyps, 27 serrated adenomas, and 20 traditional adenomas was studied. No significant change in apomucin expression was observed in traditional adenomas compared with normal colorectal epithelium, except for MUC5AC, which was present in 12 of the adenomas (60%) and only 20% of the normal samples. In both hyperplastic polyps and serrated adenomas, MUC2 and MUC5AC mucin expression was consistently and markedly increased. In 50% of the hyper-plastic polyps, MUC4 was reduced but in the remaining cases was similar to normal. Loss of MUC4 expression was observed in all serrated adenomas. MUC1 was not increased in the hyperplastic polyps but increased expression was seen in 17 of the serrated adenomas (63%). Similar altered distribution patterns of MUC2, MUC4, and MUC5AC were seen in hyperplastic polyps and serrated adenomas, whereas traditional adenomas showed little change from normal patterns of expression. Although hyperplastic polyps are commonly defined as benign lesions without neoplastic potential, the similar phenotypes of hyper-plastic and serrated adenomas and the existence of mixed polyps suggest that these lesions may represent a histogenetic continuum. (J Histochem Cytochem 47:1039–1047, 1999)
Keywords
T
The MUC1 gene located on chromosome 1q21-24 is fully sequenced and encodes a transmembrane glycoprotein (Swallow et al. 1987a,b). The gel-forming mucins MUC2 and MUC5AC are found in a cluster on chromosome 11p15.5 (Nguyen et al. 1990; Toribara et al. 1993; Pigny et al. 1996). The MUC2 gene codes for a typical secretory mucin that predominates in colorectal goblet cells (Tytgat et al. 1994). The MUC5AC gene is mainly expressed in gastric and tracheobronchial mucosae (Guyonnet Duperat et al. 1995). MUC4 located on chromosome 3q29 codes for a transmembrane mucin, which is mainly expressed in normal tracheobronchial and intestinal mucosa by both goblet and columnar cells (Nollet et al. 1998). Alterations in the synthesis and expression of MUC1, MUC2, MUC3, and MUC4, and the subsequent glycosylation of these protein core molecules, have been described in colorectal polyps, cancer, and inflammatory bowel disease (Lan et al. 1987; Ho et al. 1993; Buisine et al. 1996; Tytgat et al. 1996).
Traditionally, colorectal polyps have been classified into two common types: hyperplastic polyps and adenomas. In 1990, Longacre and Fenoglio-Preiser introduced the concept of mixed epithelial polyps, which can be divided into two subtypes. The first type consists of both adenomatous and hyperplastic glands that can be clearly identified as hyperplastic polyp and adenoma (HP/AD). The second type combines architectural and cytological features of hyperplastic and adenomatous epithelium. These authors proposed the term “serrated adenoma” for the latter, a distinctive subtype of colorectal epithelial polyp. Serrated adenomas show prominent glandular serration similar to hyperplastic polyps and therefore are often misdiagnosed as hyperplastic polyps. However, cytological features such as incomplete mucinous differentiation, nuclear enlargement, pseudostratification, increased mitotic activity, and increased architectural complexity are indicative of the neoplastic nature of serrated adenomas. Intramucosal carcinoma occurs in 10% of serrated adenomas (Longacre and Fenoglio-Preiser 1990).
It has been suggested that hyperplastic polyps may develop into serrated adenomas as a separate histogenetic pathway of carcinogenesis (Jass 1997). Several investigations, including both molecular and histochemical studies, support a histogenetic link between hyperplastic polyps and serrated adenomas (Veress et al. 1991; Jass and Smith 1992; Ajioka et al. 1998; Uchida et al. 1998; Iino et al. 1999).
In the present study we used monoclonal antibodies (MAbs) for the mucins MUC1, MUC2, MUC4, and MUC5AC to determine the different patterns of mucin distribution and to clarify the histogenetic relationships in hyperplastic polyps, serrated adenomas and traditional adenomas.
Materials and Methods
Formalin-fixed, paraffin-embedded human tissue specimens of endoscopically resected hyperplastic polyps (n = 12), serrated adenomas (n = 27), tubular adenomas (n = 18), and tubulovillous adenomas (n = 2) were obtained from the files of the Pathology Department, Niigata University Medical School, Japan. Histologically normal mucosa from the margins of the specimens served as control tissue. The size ranges for hyperplastic polyps, serrated adenomas, and traditional adenomas were 4.5–10 mm (mean 5.5 mm), 3–30 mm (mean 10.5 mm), and 3–25 mm (mean 8.0 mm), respectively. All of the colorectal polyps were sessile except for 9/27 serrated adenomas and 5/20 traditional adenomas, which were polypoid. Low-grade dysplasia was observed in all adenomas (Jass and Sobin 1989).
Preparation of Slides
Paraffin sections (3–4 μm) were affixed to Superfrost Plus adhesive slides (Menzel-Gläser; Braunschweig, Germany) and air-dried overnight at 37C. Sections were dewaxed in xylol and rehydrated through descending graded alcohols to Tris-buffered saline (0.05 M Tris, 0.15 M NaCl; TBS), pH 7.2–7.4. Sections were incubated in 1% periodic acid in distilled H2O for 30 min (MUC1), or in 0.1% porcine trypsin (ICN Biomedicals Australasia; Sydney Australia) with 0.1% CaCl2 in PBS for 30 min (MUC2), or transferred to 0.01 M citric acid buffer, pH 6, and boiled twice for 5 min each and then transferred to TBS (MUC4, MUC5AC) (Shi et al. 1991).
The sections were then incubated in 1.0% H2O2, 0.1% NaN3 in TBS for 10 min to block endogenous peroxidase activity, then washed in three changes of TBS for 5 min each. Nonspecific antibody binding was inhibited by incubating the sections in 4% skim milk powder in TBS for 15 min, followed by a brief wash in TBS. The sections were then placed in a humidified chamber and incubated with 10% normal (nonimmune) goat serum (Zymed; San Francisco, CA) for 20 min. Excess normal serum was decanted from the sections and the primary antibody applied overnight at room temperature, except for BC2 (MUC1), which was applied for 60 min. MAbs used were BC2 (1.5 μg˙ml-1) against MUC1 (Xing et al. 1989), 4F1 (4 μg˙ml-1) against MUC2 (Devine et al. 1993), M4.275 (5 μg˙ml-1) against MUC4 (Xing et al. 1997), and M1 (2 μg˙ml-1) against MUC5AC (Neomarkers; Fremont, CA) (Bara et al. 1998).
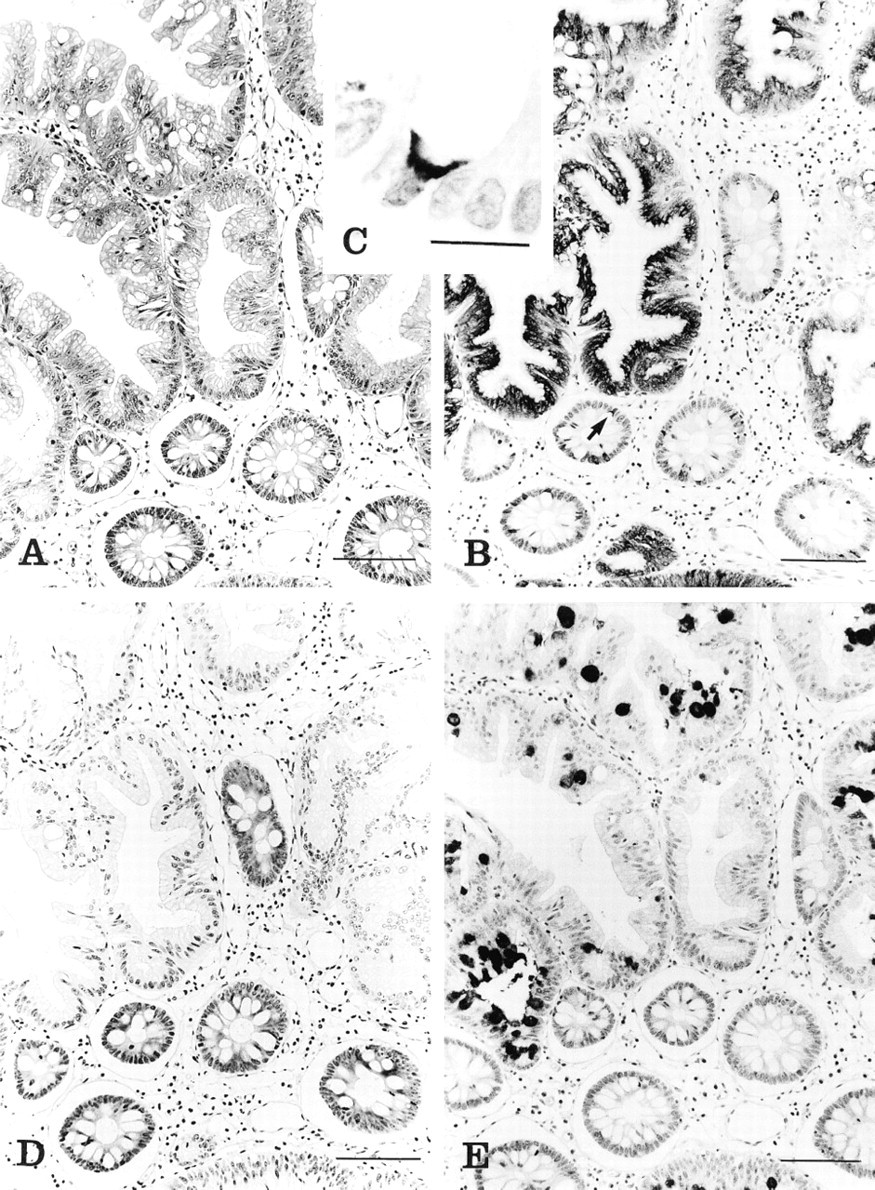
(A) Serrated adenoma and adjacent normal tissue (H&E). Bar = 100 μm. (B) Strong expression of MUC2 in the cytoplasm of both goblet and columnar cells in serrated adenoma. In the adjacent normal tissue, MUC2 is focally expressed in the perinuclear cytoplasm of the goblet cells (arrow). Immunostaining with 4F1. Bar = 100 μm. (C) High-power view of normal crypt, showing focal expression of MUC2 in the perinuclear cytoplasm of goblet cells. Immunostaining with 4F1. Bar = 15 μm. (D) Loss of MUC4 in the serrated adenoma but strong expression in the adjacent normal crypts. Immunostaining with M4.275. Bar = 100 μm. (E) MUC5AC expression is seen in the goblet cell thecae in serrated adenoma. Normal adjacent crypts show a minute trace of MUC5AC in the cytoplasm of goblet cells. Immunostaining with M1. Bar = 100 μm.
Sections were washed in three changes of TBS for 5 min each [the first buffer change contained 0.5% (v/v) Triton X-100] and then incubated with biotinylated goat anti-mouse immunoglobulins (Zymed) for 30 min. Sections were washed again in three changes of TBS for 5 min each [the first wash contained 0.1% (v/v) Triton X-100], incubated with streptavidin-horseradish peroxidase conjugate (Zymed) for 15 min, and washed in three changes of TBS for 5 min each. Color was developed in 3,3′-diaminobenzidine (Sigma Chemical; St Louis, MO) with H2O2 as substrate for 5 min. Then sections were washed in running tapwater, lightly counterstained in Mayer's hematoxylin, dehydrated through ascending graded alcohols, cleared in xylene, and mounted using DePeX (BDH Gurr; Poole, UK).
The sections were scored by two independent observers (A-E, B-H, JRJ). Apomucin staining was scored as negative (−), trace (+/-), weak (+), or strong (++). Scores were estimated by comparing intensity with the adjacent normal mucosa of the same specimen. Note was made of both cellular and subcellular localization with regard to goblet cells (theca vs perinuclear cytoplasm) and columnar cells (cytoplasm vs apical membrane).
Results
Normal Mucosa
Normal mucosa was present in 59 cases, and of these 29 (49%) showed positive staining for MUC1 (BC2) in the cytoplasm of goblet and columnar cells of the lower third of crypts, as well as along the apical membrane, whereas the remainder reacted very weakly or not at all. MUC2 (4F1) was expressed in the perinuclear cytoplasm of the goblet cells, albeit focally (Figure 1B, arrow). The cytoplasm of both columnar and goblet cells was stained for MUC4 (M4.275), staining being more intense in the lower two thirds of crypts (Figure 1C). Columnar cells were consistently negative for MUC5AC (M1), whereas in 12 of the 59 cases (20%) focal staining of the cytoplasm and the mucous droplets of goblet cells was observed (Figure 1D).
Serrated Adenomas
In the serrated adenomas, 17 of the 27 cases showed an increase in MUC1 (BC2) staining, the remaining 10 showing only very focal or negative reactivity. MUC2 (4F1) staining was strongly increased in the cytoplasm of both columnar and goblet cells (Figure 1B). Complete or near-complete loss of MUC4 (M4.275) was seen in all serrated adenomas (Figure 1C). MUC5AC (M1) staining was similar to that of MUC2, being present within the thecae of the entire mucus-secreting population, but in some cases was restricted to goblet cells (Figure 1D).
Hyperplastic Polyps
In hyperplastic polyps, MUC1 (BC2) expression was decreased compared with the adjacent normal mucosa, with only four of the 12 polyps showing positive and mainly focal staining in goblet and columnar cells. MUC2 (4F1) staining was generally increased, with MUC2 present in the cytoplasm of columnar cells as well as goblet cells (Figure 2B). MUC4 (M4.275) was negative in six cases and reduced in four, with only two cases expressing MUC4 (M4.275) at the level seen in normal mucosa (Figure 2C). MUC5AC (M1) presented a staining pattern similar to that of MUC2, with an increase in reactivity in the goblet cell theca (Figure 2D).
Traditional Tubular and Tubulovillous Adenomas
The 20 traditional adenomas included 15 flat or sessile adenomas and five polypoid adenomas. Two of the polypoid adenomas were tubulovillous and the remainder were tubular. In 12 adenomas the apical membrane and apical cytoplasm were stained for MUC1 (BC2), a weak reaction was observed in five cases, and the remaining three were negative. Of the 20 adenomas, 19 showed a normal pattern of MUC2 (4F1) expression in goblet cells (Figure 3B). One tubular adenoma showed increased MUC2 (4F1) expression in the perinuclear cytoplasm, and no change in either the intensity, intracellular localization, or crypt distribution of MUC4 (M4.275) expression was seen (Figure 3C). MUC5AC (M1) expression was increased in 12 adenomas, with the remaining eight showing only focal positivity. MUC5AC staining was mainly in the theca of goblet cells (Figure 3D).
Patterns of apomucin immunoreactivity in each category of polyp were assessed with regard to polyp size, and no discernible differences were observed with increasing lesion size. The immunohistochemical results for normal colon and colorectal polyps are summarized in Table 1.
Discussion
The majority of epithelial polyps of the colorectum can be classified as either hyperplastic polyps or adenomas. Traditionally, these have been regarded as fundamentally different lesions, with only the adenomas being neoplastic and having potential for progression to malignancy (Fenoglio and Lane 1974; Williams et al. 1980). Recent observations have challenged this view. Mixed polyps have been described that combine hyperplastic and adenomatous features (Urbanski et al. 1984). Mixed polyps may represent a combination of typical hyperplastic polyp and traditional adenoma (HP/AD), as described by Cooper et al. (1979) and Estrada and Spjut (1980), or may blend the serrated architecture of hyperplastic polyps with the cytological and architectural atypia of an adenoma. The latter has been described as serrated adenoma (Longacre and Fenoglio-Preiser 1990). Hyper-plastic polyps, serrated adenomas, and mixed polyps may occur together as a form of polyposis syndrome, and this has been recorded within a familial setting (Jeevaratnam et al. 1996). Interestingly, identical clonal changes have been identified in the hyperplastic and serrated adenomatous components of mixed polyps, suggesting that the changes are not collisions but arise as a histogenetic continuum (Iino et al. 1999).
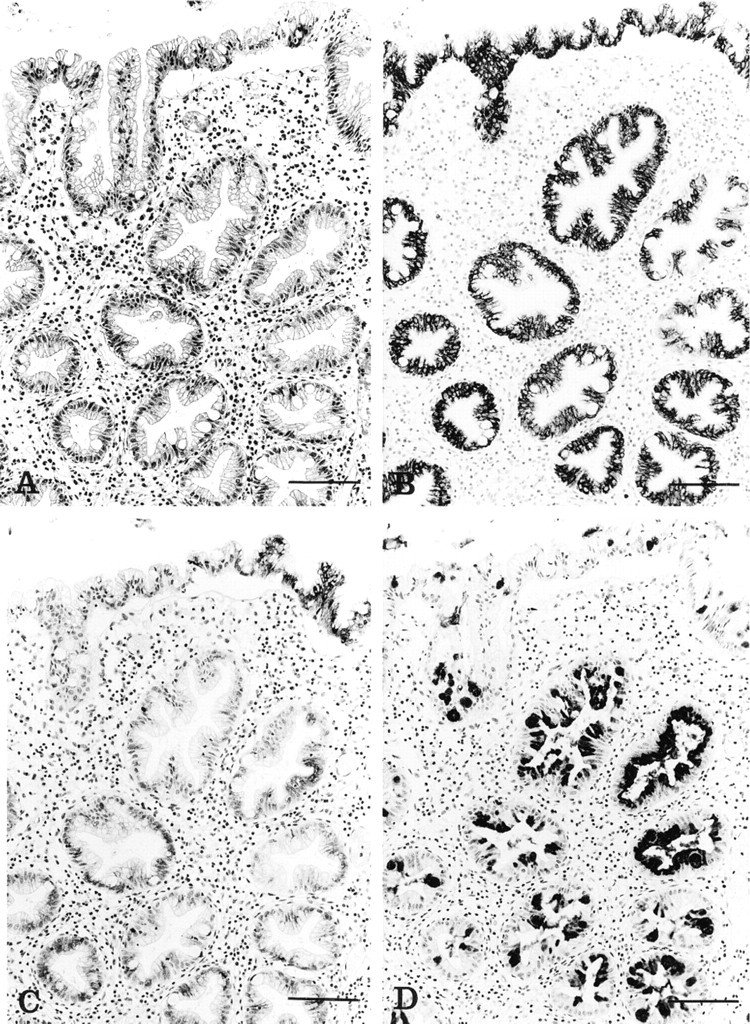
(
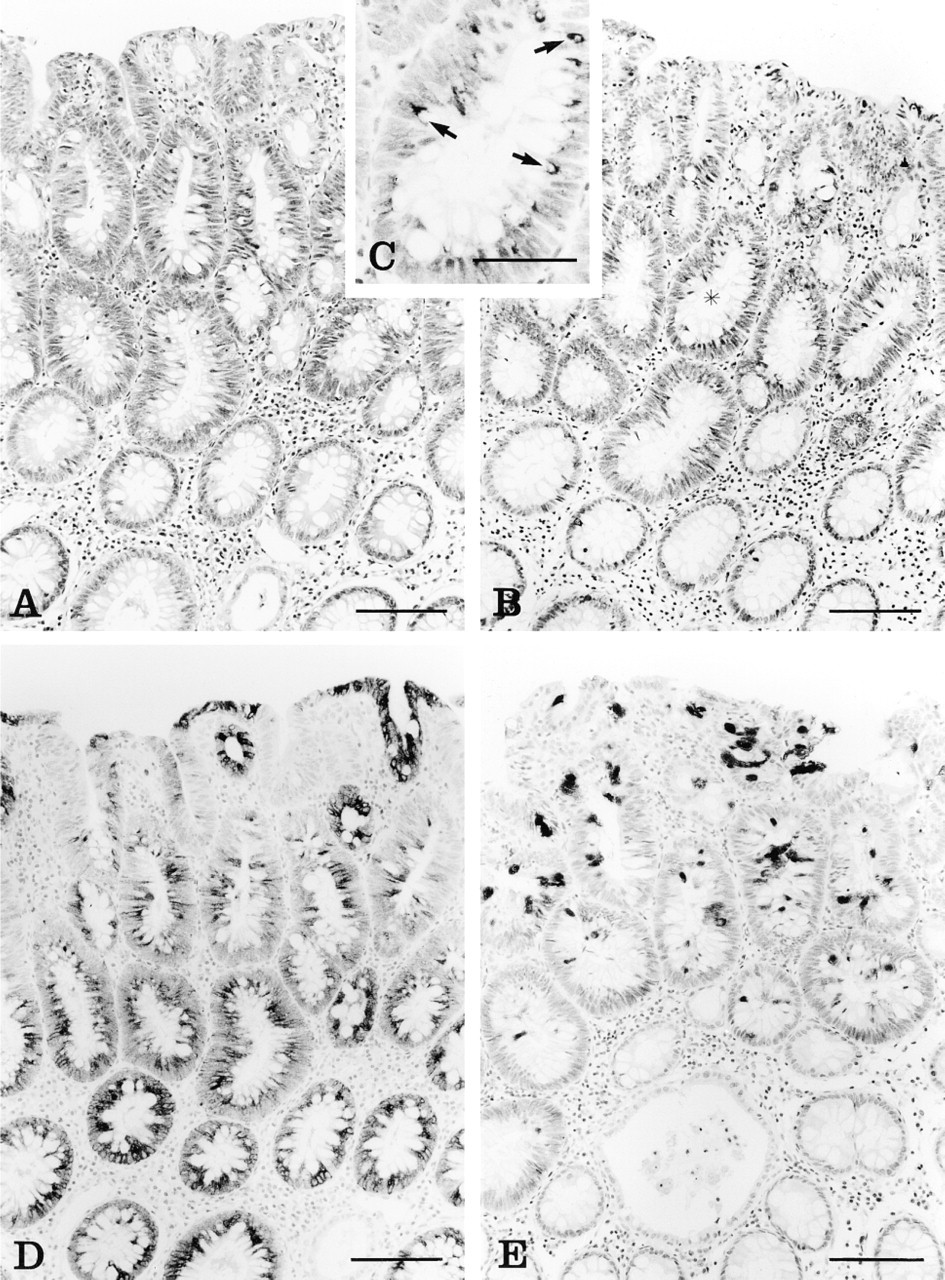
(
In normal human colon mucosa, MUC2 and MUC4 are abundantly expressed, whereas the expression or nonexpression of MUC1 has been a matter of some controversy (Ho et al. 1993; Tytgat et al. 1994; Nollet et al. 1998). In a separate study, expression of MUC1 (BC2 and MUSE11) is demonstrated at the ultrastructural level, with immunogold labeling in both columnar cells and goblet cells of the crypt base and expressed along the glycocalyx (Winterford et al. 1999). MUC2 immunoreactivity with MAb 3A2 is found exclusively in the perinuclear cytoplasm of goblet cells. MUC4 is expressed in the cytoplasm of columnar cells and goblet cells, but heavier labeling was observed in columnar cells at the ultrastructural level (Winterford et al. 1999). MUC1, MUC2, and MUC4 antibodies used in these studies react with peptide epitopes on the glycosylated domains and therefore react strongly with peptide precursors and more weakly or not at all with the mature glycosylated mucins.
MUC5AC is strongly expressed in gastric mucosa but not in normal colon mucosa (Audié et al. 1993; Buisine et al. 1998). Expression of MUC5AC has been described in fetal colon, normal colon adjacent to adenocarcinomas, and premalignant epithelium (Bara et al. 1998; Reid and Harris 1998). In the present study, patterns of MUC1, MUC2, MUC4, and MUC5AC mucin distribution in normal colorectal mucosa adjacent to the polyps were similar to those described in the literature.
The expression of apomucins in adenomas is associated with polyp size, villosity, and degree of dysplasia. Investigations of mucin expression in colorectal tubular adenomas revealed an increase of MUC1 in foci with high-grade atypia and extension throughout the crypt length with increasing atypia (Ajioka et al. 1997). This might be explained in part by reduced glycosylation. MUC2 was reduced in areas of high-grade atypia in tubular adenomas as a reflection of decreased mucin production (Ho et al. 1996; Ajioka et al. 1997). MUC2 and MUC5AC are strongly expressed in villous adenomas, which secrete copious amounts of mucus. MUC5AC was detected in mildly dysplastic villous adenomas but, in contrast to MUC2, was not expressed in mucinous adenocarcinomas (Blank et al. 1994; Buisine et al. 1996). MUC4 is strongly expressed in normal colon but is absent in many colon tumors (Xing et al. 1997). In the present study, MUC1 was not significantly increased in tubular adenomas compared to normal, but the adenomas all showed low-grade dysplasia. MUC2 was not increased in the tubular adenomas compared to normal, except in one sample. MUC4 was expressed in tubular adenomas in a pattern similar to that seen in normal mucosa. An increase of MUC5AC apomucin could be detected in half the cases. In summary, there was little change from normal in this group of mainly tubular and low-grade adenomas, except for an increase in MUC5AC expression.
Hyperplastic polyps and serrated adenomas showed essentially similar changes, but these were very different from the pattern observed in tubular adenomas. There was increased expression of MUC2 and MUC5AC and reduced expression of MUC4, the latter being more consistently seen in serrated adenomas than in hyperplastic polyps. The main difference between these polyps was with regard to MUC1, which showed increased expression in 63% of serrated adenomas but reduced expression in 67% of hyperplastic polyps. MUC1 expression is associated with neoplastic progression and tumor aggressiveness of colorectal adenocarcinoma (Hiraga et al. 1998). The MUC1 upregulation in serrated adenomas may therefore reflect not only their stage of neoplastic progression but a possible relationship to an aggressive subset of colorectal cancer. The finding of MUC2 in all cells of hyperplastic polyps and serrated adenomas suggests not merely upregulation of expression but a switch in lineage differentiation from columnar cell to goblet cell. In other words, the “columnar” cells in these lesions have adopted a goblet cell phenotype. The anomalously expressed gastric mucin MUC5AC was observed mainly in cells with classical goblet cell morphology in hyperplastic polyps as well as in serrated adenomas.

N, normal; AD, colorectal adenoma; HP, hyperplastic polyp; SA, serrated adenoma.
Intensity: negative (−); trace(+/-); weak (+); strong (++).
The loss of MUC4 is interesting because this apomucin is expressed by goblet as well as columnar cells in normal colorectal mucosa, albeit at higher levels in columnar cells (Winterford et al. 1999). The peptide sequence of MUC4 suggests that, although it is expressed by goblet cells, it is of a structural and functional subclass of mucin-like molecules that differ from the classical secreted MUC2 and MUC5AC mucins. MUC4 appears to be a transmembrane mucin and contains two extracellular EGF-like domains that may interact with members of the c-erb family of growth factor receptors (Rossi et al. 1996; Moniaux et al. 1999). The mucinous change in hyperplastic polyps and serrated adenomas appears to favor mucin of a predominantly secretory nature.
The shared phenotypes, both structural and functional, observed in hyperplastic polyps and serrated adenomas suggest that the lesions are histogenetically related. This is further supported by the finding of low levels of DNA microsatellite instability (MSI-L) in these lesions (Iino et al. 1999). About 10% of colorectal adenocarcinomas are MSI-L (Jass et al. 1998), and it will be interesting to establish if some of these cancers express the mucin phenotypes that characterize serrated polyps: MUC2+/MUC4−/MUC5AC+.
Footnotes
Acknowledgments
We wish to thank Ms Estelle Schoch and Mr Nigel Misso for invaluable technical support, and Ms Lynne Reid and Mr Clay Winterford for contributing their photographic skills.