
Abstract
To investigate the expression of MUC6 mucin in gastric carcinomas, we generated a novel monoclonal antibody (MAb CLH5) using an MUC6 synthetic peptide. MAb CLH5 reacted exclusively with the MUC6 peptide and with native and deglycosylated mucin extracts from gastric tissues. MAb CLH5 immunoreactivity was observed in normal gastric mucosa restricted to pyloric glands of the antrum and mucopeptic cells of the neck zone of the body region. In a series of 104 gastric carcinomas, 31 (29.8%) were immunoreactive for MUC6. The expression of MUC6 was not associated with histomorphological type or with clinicopathological features of the carcinomas. Analysis of the co-expression of MUC6 with other secreted mucins (MUC5AC and MUC2) in 20 gastric carcinomas revealed that different mucin core proteins are co-expressed in 55% of the cases. MUC6 was co-expressed and co-localized with MUC5AC in 45% and with MUC2 in 5% of the cases. Expression of MUC2 alone was observed in 25% of the cases. All carcinomas expressing MUC2 mucin in more than 50% of the cells were of the mucinous type according to the WHO classification. The co-expression of mucins was independent of the histomorphological type and stage of the tumors. In conclusion, we observed, using a novel well-characterized MAb, that MUC6 is a good marker of mucopeptic cell differentiation and is expressed in 30% of gastric carcinomas, independent of the clinicopathological features of the cases. Furthermore, we found that co-expression and co-localization of mucins in gastric carcinomas is independent of histomorphology and staging. Finally, we observed that intestinal mucin MUC2 is expressed as the most prominent mucin of the mucins tested in mucinous-type gastric carcinomas.
M
The expression of mucins exhibits organ-, tissue-, and cell-specific patterns (Zotter et al. 1988; Devine et al. 1993; Gambús et al. 1993; Ho et al. 1993, 1995; Lesuffleur et al. 1994). Normal gastric mucosa expresses MUC1, MUC4, MUC5AC, and MUC6 (Audie et al. 1993; De Bolós et al. 1995; Ho et al. 1995; Byrd et al. 1997; Reis et al. 1997), with MUC1, MUC5AC, and MUC6 being the most prominent (Ho et al. 1995). MUC1 is widely expressed in mucous cells of the surface epithelium of the antrum, and is focally expressed in oxyntic glands of the body and pyloric glands of the antrum (Burchell et al. 1987; Girling et al. 1989; Ho et al. 1993, 1995; Walker et al. 1995; Reis et al. 1998a). MUC5AC is highly expressed in foveolar cells of the body and the antrum (De Bolós et al. 1995; Ho et al. 1995; Byrd et al. 1997; Reis et al. 1997), whereas MUC6 is expressed in mucopeptic cells of the neck region of the body and in pyloric glands of the antrum (De Bolós et al. 1995; Ho et al. 1995; Byrd et al. 1997; Bartman et al. 1998; Reis et al. 1999). The characteristic cell- and tissue-specific distribution of mucin expression observed in the gastric mucosa may be related to the specific function of each mucin (De Bolós et al. 1995; Ho et al. 1995; Byrd et al. 1997).
Quantitative and qualitative alterations of mucin expression have been frequently reported in gastric carcinomas (Bara et al. 1993; Gambús et al. 1993; Ho et al. 1993, 1995; Carrato et al. 1994; Reis et al. 1997, 1998a; Sakamoto et al. 1997; Baldus et al. 1998; Utsunomiya et al. 1998). The mucin expression pattern of gastric carcinomas is heterogeneous. It includes mucins normally expressed in the gastric mucosa (MUC1, MUC5AC and MUC6) and de novo expression of the intestinal mucin MUC2 (Ho et al. 1993, 1995; Carrato et al. 1994; Ho et al. 1995; Sakamoto et al. 1997; Reis et al. 1998a; Baldus et al. 1998; Utsunomiya et al. 1998). Furthermore, decreased expression of “gastric” mucins and de novo expression of MUC2 have also been described in intestinal metaplasia (Ho et al. 1995; Filipe et al. 1996; Reis et al. 1998b, 1999), a precancerous lesion of gastric carcinomas (Correa 1988, 1992; Filipe et al. 1994). These observations suggest that the repertoire of mucins synthesized by gastric carcinoma cells is tightly associated with their differentiation program. The pattern of mucin expression may therefore provide new insights about the differentiation pathways of gastric carcinomas.
Here we report the development of monoclonal antibodies (MAbs) CLH4 and CLH5 against a synthetic peptide based on the MUC6 tandem repeat amino acid sequence. The biochemical and immunohistochemical properties of MAb CLH5 are described in detail. MAb CLH5 was used to study the expression profile of MUC6 in 104 gastric carcinomas to determine if there is any association between the expression of MUC6 and the clinicopathological characteristics of the carcinomas. Furthermore, we aimed to analyze if the expression of MUC6 is related to the expression of another “gastric” mucin, MUC5AC, and/or to that of the “intestinal” mucin MUC2.
Materials and Methods
Synthetic Peptides
The synthetic peptides used in this study (Table 1) were HPLC-purified and the structure confirmed by amino acid analyses and mass spectrometry. Peptides MUC1, MUC2, and MUC5AC were synthesized by Carlbiotech (Copenhagen, Denmark). Peptide MUC7 was synthesized by Neosystem (Strasbourg, France). Peptide MUC3 was a kind gift of Dr. G. Hansson. Peptide MUC6 was a kind gift of Dr. C de Bolós (IMIM, Barcelona). Peptide MUC6 sequence corresponds to an antigenic region within the tandem repeat of the MUC6 mucin and was selected using the PEP polypeptide analysis system version 5.4 (IntelliGenetics; Mountain View, CA). Peptide MUC6 and control peptide MUC5AC were coupled to maleimide-activated keyhole limpet hemocyanin (KLH; Pierce, Rockford, IL) by mixing 1 mg of peptide with 1 mg of KLH, followed by incubation for 1 hr at room temperature (RT). The coupled peptides were dialyzed overnight at 4C against PBS.
Production of Monoclonal Antibodies
Female 8-12-week old BALB/c mice were immunized IP three times with the coupled MUC6 peptide mixed with Freund's adjuvant, with an interval of 14 days between immunizations. Blood samples from the immunized mice were collected 7 days after the third immunization and the sera tested by ELISA, immunohistology, and Western blotting. Three days after an IV boost, spleen cells from one mouse were fused with NS-1 cells and hybridomas secreting anti-MUC6 antibodies were cloned by limiting dilution. The screening strategy involved (a) direct ELISA using synthetic peptides, (b) inhibitory ELISA with the MUC6 peptide and irrelevant peptides, (c) immunohistology using normal gastric mucosa, and (d) Western blotting with native and deglycosylated mucin extracts from gastric mucosa. Hybridomas producing MAbs specifically reactive with MUC6 peptides and showing reactivity with gastric mucin samples in Western blots were selected. Two hybridomas, CLH4 and CLH5, were cloned. Immunoglobulin isotypes were determined using subclass-specific goat anti-mouse peroxidase-conjugated antibody (Southern Biotechnology Associates; Birmingham, AL).
Synthetic Peptides
Enzyme-linked Immunosorbent Assay
Maxisorp plates (Nunc; Roskilde, Denmark) were coated with peptides diluted in PBS, pH 7.2, overnight at 4C. Plates were blocked with 5% BSA diluted in PBS, followed by incubation for 1 hr with hybridoma supernatants or sera. Plates were then incubated with peroxidase-conjugated rabbit anti-mouse antibody (DAKO; Glostrup, Denmark) for 1 hr and developed using 0.4 mg/ml of 1,2-phenylendiamin (OPD) tablets (DAKO) in 0.1 citric acid-phosphate buffer (pH 5.0) and 1 μl of 30% H2O2/ml of development solution. The inhibitory ELISA was carried out as above, except that the primary antibody was incubated with inhibitor for 1 hr before it was added to the coated and blocked ELISA plate.
Gastric Mucin Extracts, SDS-PAGE, and Western Blotting
Gastric mucins were obtained from surgical specimens of patients with gastric carcinoma undergoing surgical resection in Hospital S. João (Porto, Portugal). Samples were collected after informed consent. Mucin samples from antral gastric mucosa adjacent to carcinomas were purified, characterized, and deglycosylated as previously described (Reis et al. 1997). Mucins were partially deglycosylated by trifluoromethanesulfonic acid (TFMSA) (Sojar and Bahl 1987). TFMSA-treated mucin was monitored with the anti-Tn antibody HB-Tn1 (DAKO). Electrophoresis was carried out using pre-casted 4-15% gradient polyacrylamide gels, which were transferred to nitrocellulose in the Pharmacia Phastsystem according to the manufacturer's instructions (Pharmacia; Uppsala, Sweden). Western blotting was performed as described elsewhere (Reis et al. 1997).
In Vitro Glycosylation of Peptides
Peptide MUC6 and control peptide MUC2 were glycosylated in vitro using a recombinant enzyme UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferase T2 (GalNAc-T2). The production and characterization of the recombinant enzyme have been described elsewhere (White et al. 1995; Wandall et al. 1997). Glycosylation of 50 μg of peptide was performed using a reaction mixture consisting of 25 mM Tris-HCl (pH 7.2), 0.25% Triton X-100, 5 mM MnCl2, 5 mM 2-β-mercaptoethanol, 10 mM UDP-[14C]-GalNAc (300 cpm/nmol), and 5 mU/ml enzyme in a final volume of 100 μl. To obtain different glycoforms of the peptide, the glycosylation mixture was incubated at 37C for different periods of time to a maximum of 72 hr. Glycosylation was monitored by reversephase HPLC and matrix-assisted laser desorption ionization mass spectrometry (MALDI-MS).
Determination of Glycopeptide Structure
The different glycoforms of the peptide MUC6 were purified on a microbore HPLC system (ABI 130A; Perkin-Elmer, Norwalk, CT) using a Brownlee C18 narrow-bore column (30 mm-2.1 mm, 7-μm particle size), with a gradient of 0-80% acetonitrile in 0.1% trifluoroacetic acid (TFA). HPLC fractions were collected manually and analyzed by MALDI-MS. MALDI mass spectra of nonglycosylated and glycosylated peptides were acquired on a Voyager-Elite MALDI time-offlight mass spectrometer (PerSeptive Biosystems; Framingham, MA) equipped with delayed extraction. The MALDI matrix was 2,5-dihydroxybenzoic acid 10 g/liter (Aldrich; Milwaukee, WI) dissolved in 0.1% TFA in water/acetonitrile 2/1 (v/v). Reverse-phase HPLC purified samples were prepared for MALDI analysis by placing 1 μl of the fractions on a probe tip followed by 1 μl of matrix solution. All mass spectra were obtained in the linear mode and calibrated using external calibration. Data processing was carried out using GRAM/386 software.
Immunohistochemistry
Normal tissue samples from several organs were selected from the files of the Department of Pathology, Hospital S. João (Porto, Portugal), using surgical specimens removed from patients with nonneoplastic diseases not affecting the tissues under study (Table 2). Gastric carcinoma specimens (n = 104) were obtained from patients undergoing surgery. All tissue sections were collected after informed consent. Sections were fixed in formalin buffered with PBS and embedded in paraffin. Hematoxylin and eosin-stained sections were used to classify gastric carcinomas according to the classifications of Carneiro et al. (1995), Laurén (1965), Ming (1977), and the World Health Organization (WHO) (Watanabe et al. 1990). Mucinous carcinomas were defined as tumors with abundant intracellular and extracellular mucus occupying more than 50% of the sections evaluated (Watanabe et al. 1990). Clinicopathological characteristics of the cases (depth of penetration of the gastric wall, lymphatic invasion, nodal metastasis, venous invasion, pTNM staging) were also recorded in every case. Intestinal metaplasia was classified according to Filipe (1986,1992) as complete type, which is characterized by the presence of absorptive cells, Paneth cells, and goblet cells secreting sialomucins, and the incomplete type, which is characterized by columnar cells and goblet cells with both cell types secreting sialo- and/ or sulfomucins.
Immunoreactivity of antibody CLH5 with normal tissues

Sections from normal tissues and gastric carcinomas were immunostained by the avidin-biotin complex method (Hsu et al. 1981). Paraffin sections were dewaxed and rehydrated. Sections designed for neuraminidase treatment were washed twice in Tris-buffered saline (TBS), pH 7.6, and incubated with neuraminidase from Clostridium perfringens Type VI (Sigma; St Louis, MO) diluted in 0.1 M Na-acetate buffer, pH 5.5, to a final concentration of 0.1 U/ml. The incubation was carried out for 2 hr at 37C and was followed by three washings in ice-cold water. Sections were treated with 0.5% H2O2 in methanol for 30 min, followed by 20 min of incubation with rabbit nonimmune serum. Sections were rinsed and incubated with primary antibody (Table 3) overnight at 4C. Sections were rinsed and incubated with biotin-labeled rabbit anti-mouse serum diluted 1:200 in TBS for 30 min, rinsed with TBS, and incubated with avidin-biotin-peroxidase complex for 1 hr. Sections were rinsed in TBS and developed with 0.05% 3,3′diaminobenzidinetetrahydrochloride (DAB) freshly prepared in 0.05 M TBS containing 0.1% H2O2. Sections were counterstained with hematoxylin, dehydrated, and mounted. Antibodies and nonimmune serum were diluted 1:5 in TBS containing 1% BSA. All series included positive controls; negative controls were performed by substitution of the primary MAbs with immunoglobulins of the same class and at the same concentration.
Co-expression of MUC6, MUC5AC, and MUC2 was analyzed in a group of 20 gastric carcinomas. The MAbs used are listed in Table 3.
Scoring of Immunostaining in Gastric Carcinomas and Statistical Analysis
A semiquantitative approach was used to score the immunostaining: +, few positive cells (<25%); ++ well-defined areas with positive cells (25-50%); +++, extensive areas with positive cells (50-75%); ++++, most cells stained (>75%). The chi-square test with the Yates correction was used for statistical analysis. Differences were considered to be significant at p<0.05. Fisher's test was applied whenever appropriate.
Specificity and reference of monoclonal antibodies
Code number MAb2011 from Chemicon International (Temecula, CA)
Results
Production and Characterization of the Anti-MUC6 MAb
Two hybridomas, CLH4 (IgG1) and CLH5 (IgG1), were selected and cloned. Both MAbs reacted specifically with the MUC6 peptide in ELISA and stained mucin extracts from gastric mucosa by Western blotting. MAb CLH5 was used for further immunohistochemical studies because it showed stronger staining with paraffin-embedded sections. Figure 1 shows an ELISA testing MAb CLH5 with several synthetic peptides (MUC1, MUC2, MUC3, MUC5AC, MUC6, and MUC7). MAb CLH5 reacted only with MUC6 peptide. Inhibitory ELISA also showed the specificity of MAb CLH5 for the MUC6 peptide (not shown).
Western blot analyses showed that MAb CLH5 reacted with mucin extracts from gastric mucosa in the high molecular weight range (Figure 2). Deglycosylation of mucins with TFMSA has been shown to cleave carbohydrate side chains of mucins, leaving some of the innermost carbohydrate residue, GalNAcα1-O-Ser/Thr (Tn antigen) (Gerken et al. 1992). MAb CLH5 showed a stronger reactivity with mucin extracts after TFMSA deglycosylation. MAb HB-Tn1 (anti-Tn antigen) was used as control to monitor deglycosylation of the samples (Figure 2).
To evaluate the interference of GalNAc glycosylation with the epitope recognized by CLH5 in vitro, GalNAc-O-glycosylation of the MUC6 peptide was performed. The exhaustively glycosylated MUC6 peptide incorporated up to 5 mol of GalNAc per mol of peptide (Figure 3). MAb CLH5 showed reactivity in an inhibitory ELISA with the different glycoforms of the MUC6 peptide, as shown in Figure 4.
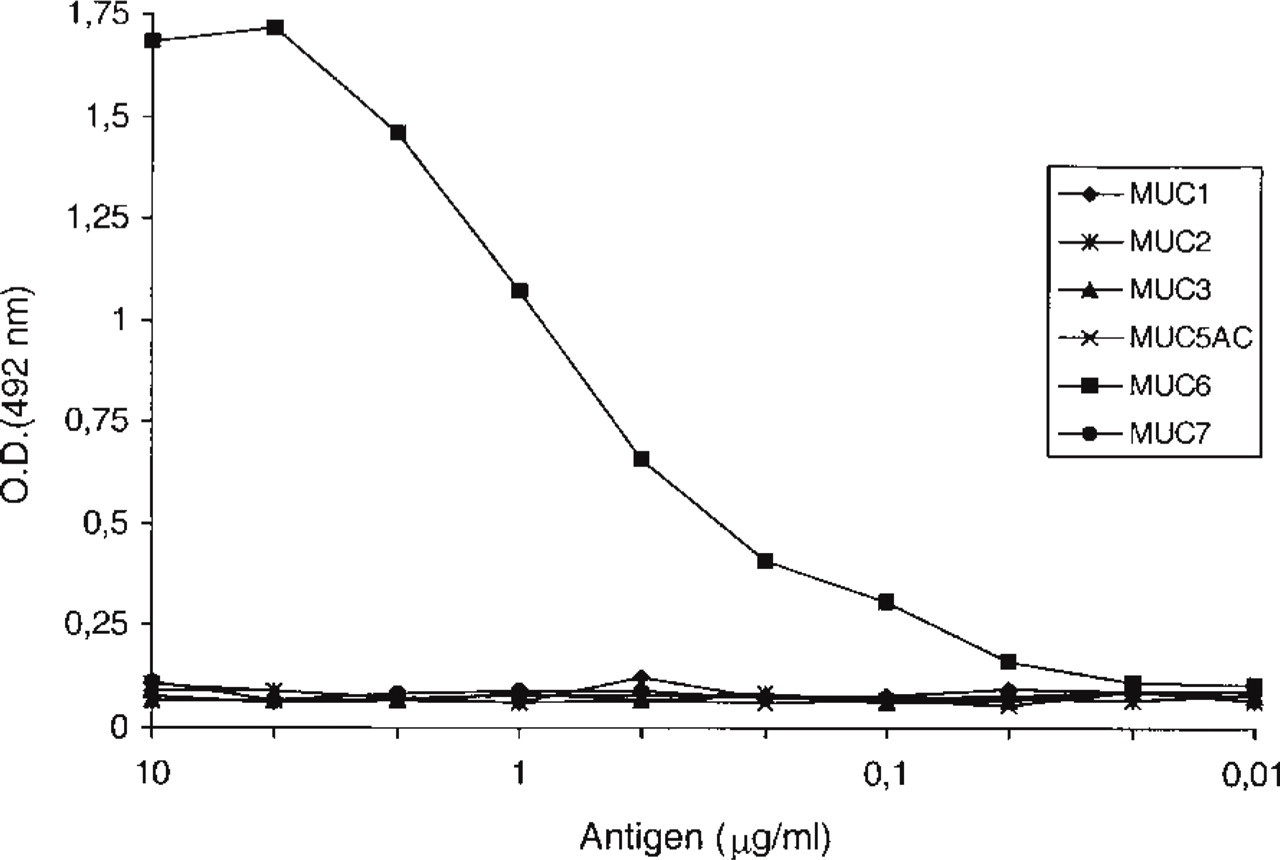
Immunoreactivity of MAb CLH5 with mucin-related peptides in ELISA. MAb CLH5 reacted specifically with MUC6 peptide.
MUC6 Distribution in Normal Adult Tissues, Including Gastric Mucosa, Detected with the Novel MAb CLH5
Table 2 summarizes the reactivity of MAb CLH5 with normal gastrointestinal and non-gastrointestinal adult human tissues. In the stomach, MAb CLH5 stained mucopeptic cells, which are characterized by particular histochemical and ultrastructural features of the mucin granules (Fiocca et al. 1987; Hanby et al. 1999) and include the glands of the antrum and the mucous cells of the neck zone of the body region. The predominant staining patterns were diffuse cytoplasmic and perinuclear. Foveolar cells of both antrum and body regions as well as chief and parietal cells of oxyntic glands did not react with MAb CLH5. Neuraminidase treatment of the samples did not change the reactivity pattern. MAb CLH5 also stained deep glands of the gallbladder and Brunner's glands of the duodenum, as well as the cytoplasm of a few acinar cells of the pancreas (Figure 5).
Expression of MUC6 in Gastric Carcinomas
In contrast to normal gastric mucosa, only 31 of 104 gastric carcinomas (29.8%) displayed immunoreactivity for CLH5. The expression of MUC6 mucin in positive cases was heterogeneous, either clustered in focal areas or dispersed in isolated cells, and showed variable scoring (+, 8 cases; ++, 18 cases; +++, 4 cases; ++++, 1 case). Positive cells showed diffuse cytoplasmic staining. The comparison of MUC6 expression in the different histomorphological types of gastric carcinoma disclosed no significant differences. The expression of MUC6 was not associated with other clinicopathological characteristics of gastric carcinomas, such as depth of wall penetration, lymphatic invasion, nodal metastasis, venous invasion, and pathological TNM stage (Table 4).
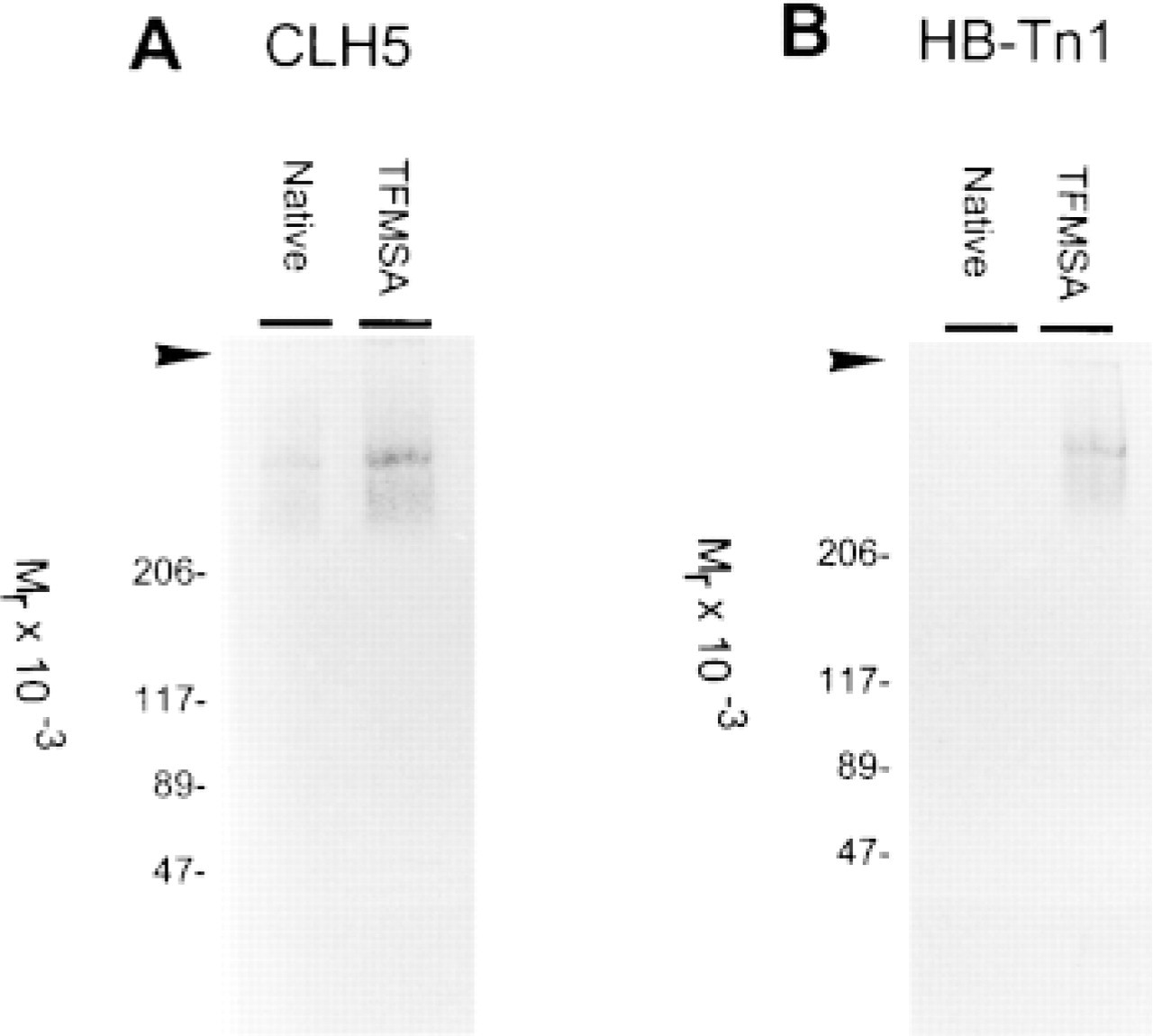
Western blots using (
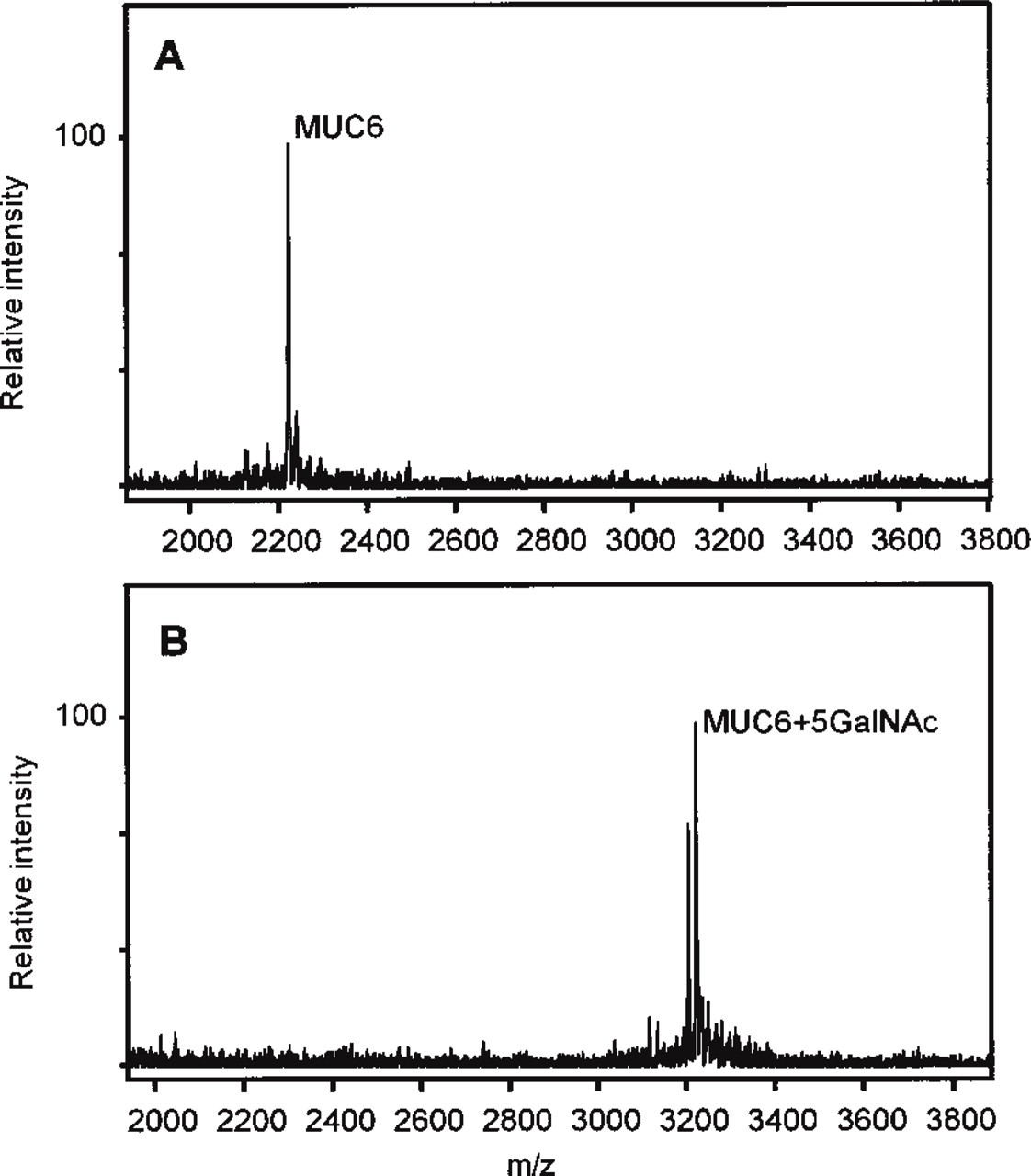
Mass spectrometry of unglycosylated and glycosylated MUC6 peptide. (
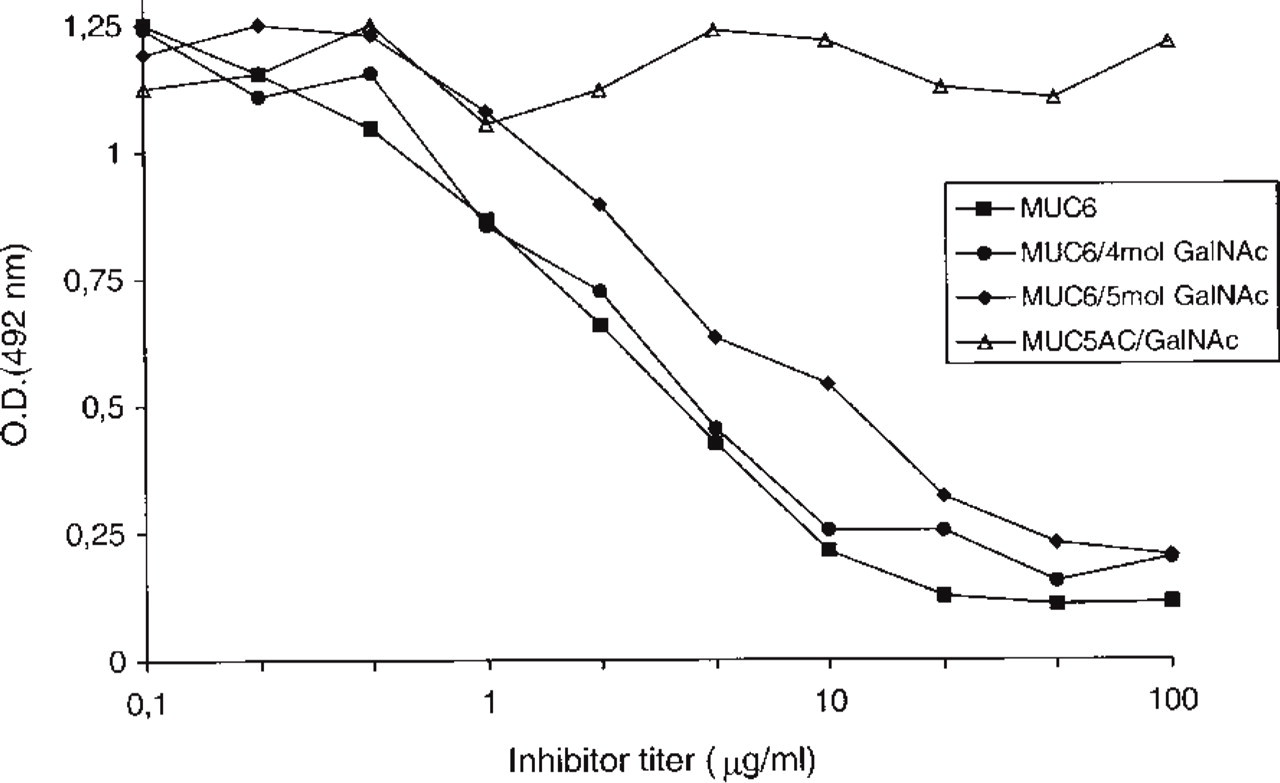
Immunoreactivity of MAb CLH5 in an inhibitory ELISA. The plate was coated with peptide MUC6. MAb CLH5 was preincubated with four different peptides and glycopeptides as described in Materials and Methods. Peptide MUC6 and glycopeptides MUC6 containing up to 5 mol GalNAc inhibited the binding of MAb CLH5 to the coated MUC6 peptide. Control peptide MUC5AC/GalNAc did not inhibit the binding of MAb CLH5 to the coated MUC6 peptide.
Co-expression of Secreted Mucins (MUC6, MUC5AC, and MUC2) in Gastric Carcinomas
Analysis of the co-expression and co-localization of the gastric secreted mucins MUC6 and MUC5AC and the intestinal mucin MUC2 in a group of 20 gastric carcinomas is shown in Table 5. We observed that the majority of gastric carcinoma cases (n = 11; 55%) express more than one of the studied mucins. Co-expression and co-localization of MUC6 with MUC5AC and MUC2 were observed in nine cases and one case, respectively. Co-expression and co-localization of MUC5AC and MUC2 were observed in five cases. Simultaneous expression and co-localization of MUC6, MUC5AC and MUC2 were observed in one case (case 6). Topographic heterogeneity of the mucin expression profile
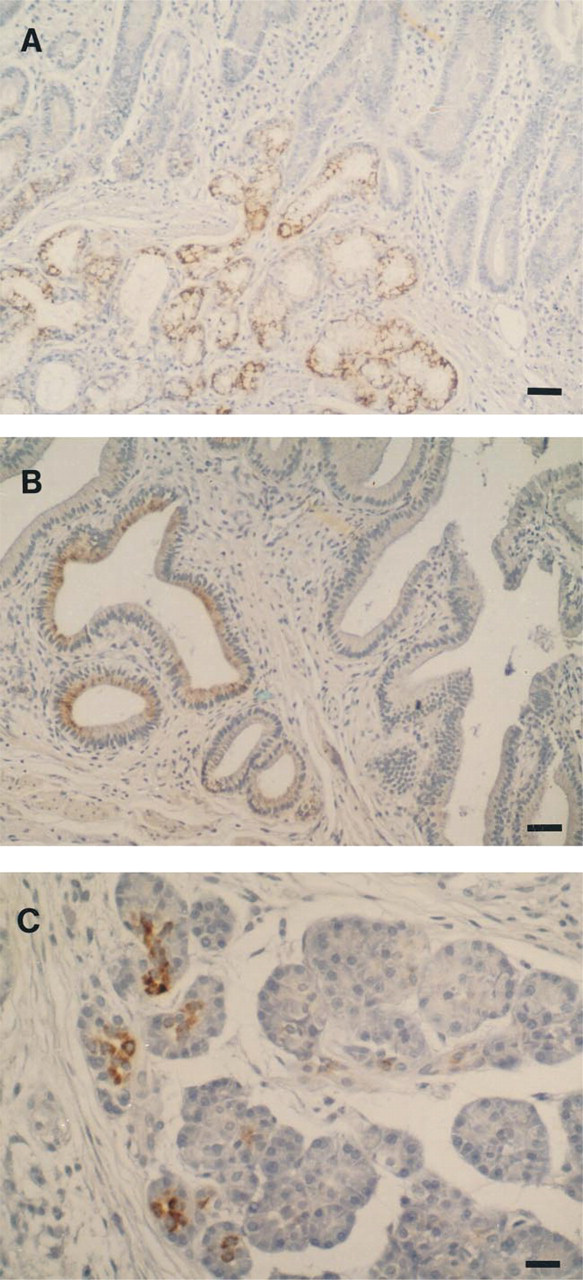
Immunohistochemistry with monoclonal antibody CLH5 in Brunner glands (
The patterns of mucin co-expression were not associated with the histological characteristics nor with the stage of the tumors (Table 5).
Relationships between clinicopathological features of gastric carcinomas and MUC6 expression (CLH5) immunoreactivity
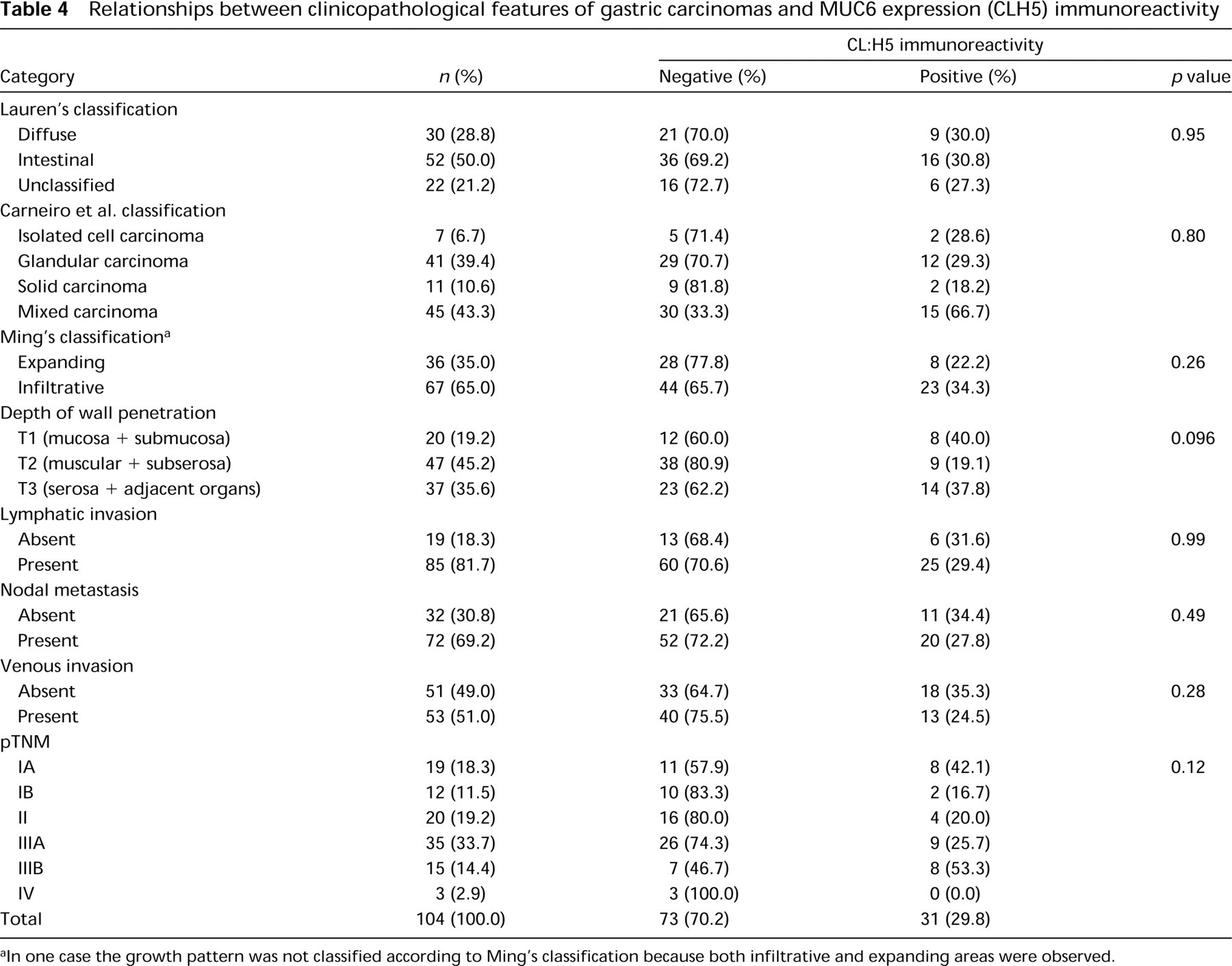
In one case the growth pattern was not classified according to Ming's classification because both infiltrative and expanding areas were observed.
Discussion
In this article we describe the production and characterization of two MAbs to MUC6 mucin. These MAbs, CLH4 and CLH5, reacted exclusively with a peptide based on the tandem repeat of MUC6, as well as with native and deglycosylated mucin extracts from gastric tissues. MAb CLH5 reacted by immunohistology in agreement with the results of earlier immunohistochemical studies using polyclonal antibodies as well as with the results obtained by in situ hybridization (De Bolós et al. 1995; Ho et al. 1995; Byrd et al. 1997; Bartman et al. 1998).
As previously described (De Bolós et al. 1995; Ho et al. 1995; Byrd et al. 1997; Bartman et al. 1998), we observed MUC6 expression in the glands of the antrum and in mucopeptic cells of the neck zone of the body region of the stomach. Furthermore, we observed expression of MUC6 in the Brunner's glands in the duodenum, glands of the gallbladder, and in a few acinar cells of the pancreas. The immunocytolocalization pattern in normal tissues was mainly perinuclear and supranuclear, but some cells showed diffuse cytoplasmic staining, which may be due to variability of the type and/or rate of glycosylation in different cells. This staining pattern suggests that MAb CLH5 recognition of its epitope may be affected in some glycoforms of MUC6 mucin. These observations are further supported by the discrepancy between the staining pattern we observed in pancreatic tissues, with staining confined to a few acinar cells, and a previous report by Reid et al. (1997) showing that expression of MUC6 evaluated by mRNA in situ hybridization and immunocytochemistry is abundant in pancreatic ducts and centroacinar cells. To study the effect of GalNAc O-glycosylation on the antibody's epitope the MUC6 peptide was O-glycosylated using recombinant GalNAc-transferase-T2. Five mol of GalNAc per mol of peptide were incorporated, indicating that five of the potential nine sites in the MUC6 peptide were GalNAc-glycosylated. The reactivity of the MAb CLH5 was not inhibited by the incorporation of 5 mol of GalNAc in the peptide, suggesting that the epitope may be located in a region that is not glycosylated or is only partially glycosylated by in vitro O-glycosylation. In any case, the in vitro data do not exclude that in vivo glycosylation may affect CLH5 immunoreactivity.
Co-expression of secreted mucins MUC5AC, MUC6, and MUC2 in 20 gastric carcinomas a
When areas with different co-expression patterns were found they are depicted in the second column. Large boxes represent areas in which predominant expression of one mucin was found. Smaller boxes represent areas in which other mucins were found within the areas defined by larger boxes. When more than one box is presented without intersection, this means that areas expressing other mucins did not overlap. Dif, diffuse; Int., intestinal; Unclas., unclassified; SRC, signet ring carcinoma; Tub., tubular; Mucin., mucinous.
In normal gastric mucosa, CLH5 immunoreactivity was observed exclusively in the glands of the antrum and in the mucopeptic cells of the neck zone of the body region of the stomach. This observation indicates that MUC6 appears to be a good marker of mucopeptic differentiation. In this study we evaluated the expression of MUC6 in gastric carcinomas and observed a loss of MUC6 expression in the majority of the cases. Only 30% of gastric carcinomas showed expression of MUC6, and most of them expressed MUC6 in less than 50% of the cells. In agreement with our results, Ho et al. (1995) have shown a decreased expression of MUC6 in gastric carcinomas by immunohistochemistry, using a polyclonal antibody reacting with MUC6. Ho et al. (1995) immunodetected MUC6 in 57% of a series of 33 cases, a percentage that is higher than that observed in our series. Recent data from López-Ferrer et al., using a polyclonal antibody directed to the MUC6 tandem repeat (De Bolós et al. 1995) also show a higher percentage of immunoreactive gastric carcinomas (López-Ferrer et al., personal communication). We postulate that differences in specificities and different levels of tolerance to glycosylation among the antibodies may account for these discrepancies.
The present study showed that expression of MUC6 is not associated with the histological type of the gastric carcinomas. This finding is in agreement with the presence of heterogeneous cell populations in gastric carcinomas of different histotypes (Fiocca et al. 1987; Carneiro et al. 1992; Tatematsu et al. 1993; Solcia et al. 1996). If we assume that MUC6 expression is associated with mucopeptic differentiation, our results indicate that mucopeptic differentiation is not a prominent feature in gastric carcinomas and, moreover, that when present it is not associated with any histological type of gastric carcinoma. These findings agree with previous histochemical, immunohistochemical, and ultrastructural observations showing that mucopeptic cells occur only in a few gastric carcinomas, regardless of their histotype (Fiocca et al. 1987).
In normal tissues, the type of mucin expressed is closely associated with the cell phenotype: MUC5AC with foveolar cells, MUC6 with mucopeptic cells, and MUC2 with goblet intestinal-type cells. In a previous study we found that, in gastric carcinomas, expression of MUC5AC was significantly associated with diffuse-type carcinomas (Reis et al. 1997) and suggested that this is in agreement with the concept that this type of tumor usually keeps a “gastric” differentiation (Fiocca et al. 1987; Carneiro et al. 1992; Reis et al. 1997). In fact, MUC5AC is specifically expressed by foveolar and neck cells of the normal gastric mucosa (Reis et al. 1997) that are considered as gastric-specific mucus-producing cells (Fiocca et al. 1987). The lower percentage of diffuse gastric carcinomas expressing MUC5AC in the subseries evaluated for mucin co-expression (three of six cases; see Table 5) in the present study, compared to our previous observations (Reis et al. 1997), may reflect the small number of cases analyzed or an actual decrease of MUC5AC-expressing diffuse carcinomas when they acquire a mucinous phenotype. In the present study, expression of MUC6 was found to correlate to mucopeptic differentiation in normal gastric tissues and the expression of MUC6 in carcinomas supports that this may also apply to carcinoma cells (see above).
We observed the co-expression of different mucin core proteins in 55% of our series of gastric carcinomas. The presence of more than one cell type in a single tumor has been frequently observed in gastric carcinomas, with the different cell types occurring either in separate areas or mixed in a single area (Fiocca et al. 1987). Our results on the relatively frequent mucin co-expression in gastric carcinomas may reflect the heterogeneity of cell types in the tumors, with each cell type expressing one mucin. Alternatively, one can admit that genetic or epigenetic alterations may lead to the altered expression of several mucins by the carcinoma cells. The two aforementioned hypotheses are not mutually exclusive and should be clarified in future studies using immunoelectronic microscopy with double labeling techniques.
In our series, the prominent expression of MUC2 appears to be a hallmark of the mucinous type of gastric carcinoma. MUC2 immunodetection was confirmed using two different MAbs specific for MUC2. The “ectopic” expression and overexpression of MUC2 have been described as common features of mucinous carcinomas of the colon, pancreas, breast, and ovary (Hanski et al. 1997a). Our data indicate that mucinous gastric carcinomas, similar to mucinous carcinomas in other organs, preferentially express the MUC2 mucin. Taking into account that in normal tissues as well as in intestinal metaplasia of the stomach MUC2 expression is closely associated with the goblet cell phenotype (Devine et al. 1993; Gambús et al. 1993; Ho et al. 1993; Reis et al. 1999), our results suggest that in mucinous carcinomas the goblet cell phenotype may be dominant. This assumption is in agreement with histochemical and ultrastructural studies showing that goblet cells are prominent in mucinous carcinomas (Fiocca et al. 1987). Recently, Hanski et al. (1997b) has shown that the methylation in the promoter region of the MUC2 gene is determinant for the expression of MUC2 in colon carcinoma cell lines. This mechanism of control of MUC2 gene expression in mucinous gastric carcinomas should be addressed in future studies.
At variance with Ho et al. (1995), we did not observe any trend towards the expression of several types of mucins in carcinomas with more advanced tumor stages, which may stem from the smaller number of cases analyzed in the present series (20 cases) and from the fact that only mucins MUC2, MUC5AC, and MUC6 were evaluated. Our results, including MUC2 overproduction in mucinous carcinomas regardless of staging, alternatively suggest that the type of mucin and its carbohydrate chains may be of greater importance for the putative biological role played by mucins in the progression of gastric carcinomas than the occurrence of multiple mucin expression (Sakamoto et al. 1989; David et al. 1992; Carneiro et al. 1994; Amado et al. 1998).
In conclusion, using the novel well-characterized CLH5 monoclonal antibody, we observed that MUC6 is a good marker of mucopeptic cell differentiation and is expressed in 30% of gastric carcinomas, independent of the clinicopathological features of the cases. Furthermore, we found that co-expression and co-localization of mucins in gastric carcinomas are independent of histomorphology and staging. Finally, we observed that the intestinal mucin MUC2 is expressed as the most prominent mucin in mucinous type gastric carcinomas.
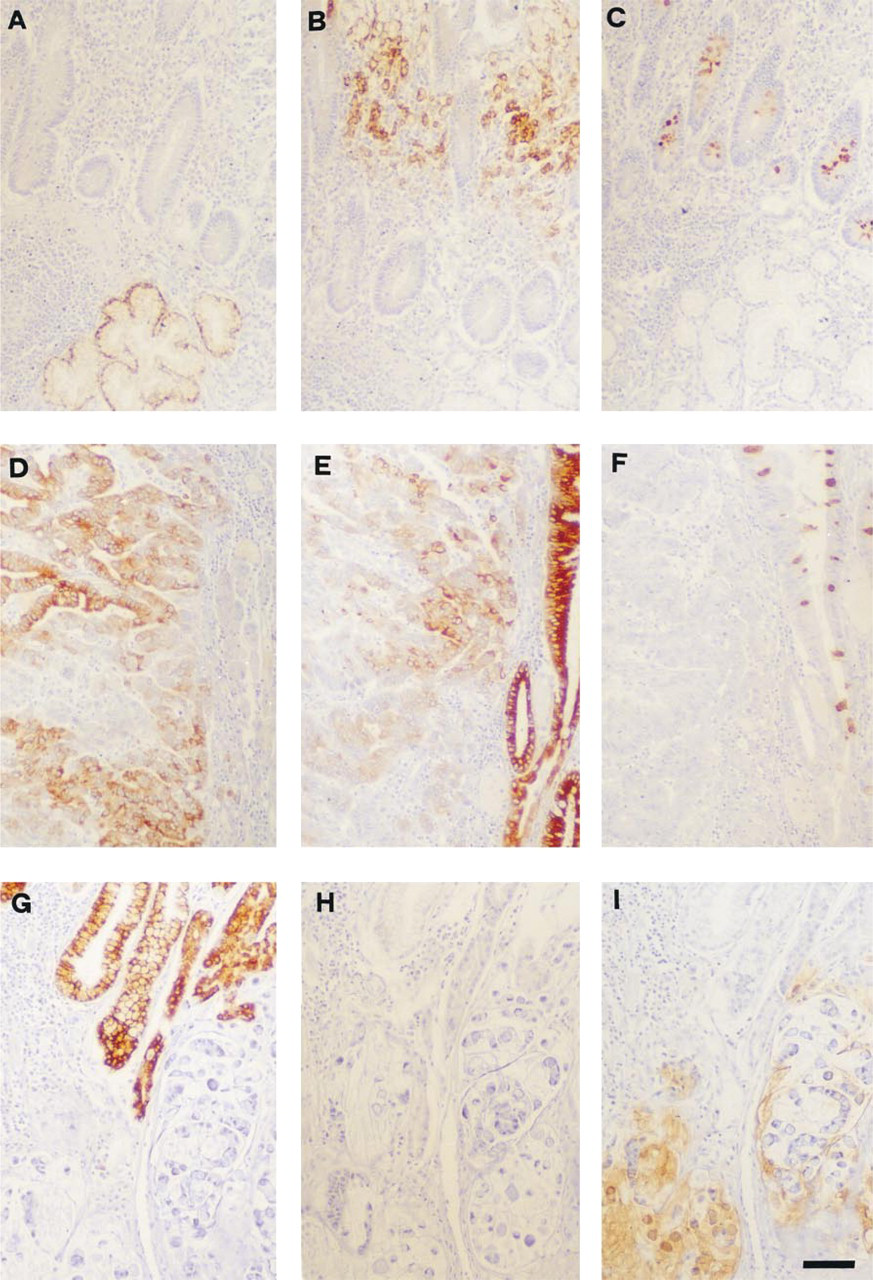
Immunohistochemistry with monoclonal antibodies: CLH5, anti-MUC6 (
Footnotes
Acknowledgments
Supported by Fundação para a Ciência e a Tecnologia (PRAXIS XXI, Project SAU/14111/1998), EU Concerted Action BMH4-CT98-3222, and by The Danish Cancer Society.
We thank Ms Lone Malte and Ms Dina Leitão for expert technical assistance.