
Abstract
MUC1, a transmembrane member of the mucin family, is believed to have anti-adhesive properties because of its highly sialylated, extended, and rigid rod-like conformation. The ERM proteins (ezrin, radixin, and moesin) function as membrane–cytoskeletal linkers. MUC1 and ezrin are enriched in microvilli in MCF-7az breast carcinoma cells. Similar localization was also found in peripheral membrane areas and in filopodium-like protrusions. Whereas ezrin was consistently detected in the cell–cell contact region, MUC1 was less frequently found there. MUC1 was distinctly expressed in long filopodial protrusions and was highly concentrated at their tips, which also contained ezrin, whereas F-actin was found along the stalk. This localization of MUC1 suggests a role for MUC1 in transient cell structures of migrating cells and transient cell adhesion. No direct association has yet been found between MUC1 and ezrin. However, both MUC1 and ezrin had a similar overall distribution pattern in microvilli and filopodium-like protrusions in immunoelectron tomography. In addition, MUC1 and ezrin showed spatial association, because several 10-nm gold particles used to decorate ezrin were seen in the vicinity close to the clusters of 5-nm gold particles decorating MUC1. Therefore, MUC1 appears to be associated with ezrin, but the nature of this association requires further study.
Keywords
M
The interactions between cell membrane structures and the cytoskeleton have an essential role in various cell functions, including coordination of cell morphology, migration, and adhesion. Several protein complexes exist that mediate interactions between cell adhesion molecules and the cytoskeleton. These include protein complexes associated with integrins in focal adhesions (e.g., α-actinin, vinculin, talin, paxillin, and focal adhesion kinase) and cadherins in cell-cell adherence junctions (β-catenin/plakoglobin–α-catenin complex) (Cowin and Burke 1996; Yamada and Geiger 1997). The ERM (ezrin, radixin, moesin) protein family members are among the proteins that have been suggested to link the cell membrane to the cytoskeleton (Algrain et al. 1993; Turunen et al. 1994; Bretscher et al. 1997; Tsukita et al. 1997; Vaheri et al. 1997). Ezrin has been localized between the plasma membrane and the core structure formed of actin filaments in microvilli (Pakkanen et al. 1987; Franck et al. 1993). Ezrin family members associate with cell adhesion molecules such as CD44, ICAM-1 and ICAM-2 (Helander et al. 1996; Hirao et al. 1996; Sainio et al. 1997; Heiska et al. 1998; Yonemura et al. 1998). Ezrin interacts with ICAM-2 in activated T-cell lymphocytes (Helander et al. 1996), and it should be noted that MUC1 has also been shown to be expressed by activated T-cell lymphocytes (Agrawal et al. 1998).
Other membrane-associated molecules, including MUC1, share ezrin's predilection for microvilli (Parry et al. 1990). It has recently been shown that another mucin, CD43, binds to ERM proteins in vitro (Yonemura et al. 1998). CD43 is an endothelial and leukocyte mucin believed to be involved in cell-cell interaction after lymphocyte activation (Thomas 1989; Wang et al. 1992; Giordanengo et al. 1995). It has been suggested that the cytoplasmic domain of transmembrane mucins binds to the cytoskeleton (Parry et al. 1990; Gendler and Spicer 1995). An unexpected link with the cytoskeleton may occur via catenins. MUC1 has been reported to interact with the cell-cell adherence junction proteins β-catenin and plakoglobin (Yamamoto et al. 1997), which are known to interact with the cell adhesion protein cadherin. α-Catenin links β-catenin and plakoglobin to actin filaments. Catenins may not be the only cytoskeletal proteins regulating the localization of MUC1. There also appear to be two discrete motifs in MUC1 that determine its apical localization. The first is located in the extracellular domain and the second is in the juxta-membrane region of the cytoplasmic domain (Pemberton et al. 1996). This allows several ways to regulate the localization of MUC1.
We now show that ezrin and MUC1 have a similar localization in MCF-7 cells and are enriched in microvilli and filopodium-like protrusions, but differ in peripheral membrane areas. We have not been able to show direct molecular interaction between MUC1 and ezrin. However, the similar distribution of MUC1 and ezrin in immunoelectron tomography suggests that MUC1 and ezrin are associated, perhaps indirectly, with each other in certain cell surface protrusions. A distinct localization of MUC1 was observed in the tips of filopodial protrusions, suggesting a role in transient adhesion to the substratum.
Materials and Methods
Cells and Antibodies
MCF-7az breast carcinoma cells were chosen for this study because this cell line expresses high amounts of MUC1 in immunofluorescence microscopy. The majority of breast carcinoma cell lines express MUC1, but MCF-7az cells were chosen because of their intensity of staining and the presence of structures hereafter referred to as filopodial protrusions, which contain large amounts of MUC1 at their tips. The U251-MG glioma cell line, which does not show endogenous MUC1 expression, was used to express exogenous MUC1 cDNA. MCF-7az and U251-MG cells were cultured in MEM supplemented with 10% fetal calf serum (FCS) medium. Antibodies used in this study were a polyclonal rabbit antibody to human ezrin (Ez-Bl-III) (Pakkanen et al. 1987), a monoclonal mouse antibody to human ezrin (3C12) (Helander et al. 1996), a monoclonal mouse antibody to human actin (Sigma; St Louis, MO) and MUSE11, a monoclonal mouse antibody to human MUC1 (kindly provided by Dr. Yuri Hinoda, Sapporo Medical University, Japan). MUSE11 recognizes an epitope (PDTRPAPG) corresponding to a 20-amino-acid repeat in the tandem repeat domain of the MUC1 core protein (Hinoda et al. 1993). Secondary antibodies were TRITC-conjugated donkey anti-mouse antibody and FITC-conjugated swine anti-rabbit or rabbit anti-mouse antibody (Jackson ImmunoResearch; West Grove, PA). All antibodies used in this study have been previously characterized.
Immunocytochemistry/Confocal Microscopy
Cells were grown to 50–70% confluence as described above, fixed for 15 min with 3.7% paraformaldehyde in PBS at room temperature (RT), and permeabilized for 10 min with 0.05% Triton X-100 in PBS. The Ez-Bl-III antibody was used at 1:50–1:100 dilutions, 3C12 at 1:100 dilution, and MUSE11 at 1:25 dilution in PBS. In some experiments, during secondary antibody incubation, cells were stained with rhodamine-conjugated phalloidin (Molecular Probes; Eugene, OR) or Oregon Green-conjugated DNase I (Molecular Probes) to reveal F-actin and G-actin, respectively. After incubation with the secondary antibodies, the coverslips were mounted on glass slides using Syva Microtrak mounting fluid (Behring Diagnostics; Cupertino, CA). Cells were examined by conventional fluorescence microscopy using an Olympus BH-2 microscope (Olympus America; Melville, NY) or by confocal microscopy with a confocal 410 Invert Laser Scan Microscope (Carl Zeiss; Oberkochen, Germany). The printed images from the confocal data were processed using the Apple Macintosh Adobe Photoshop program.
MUC1 cDNA (kindly provided by Dr. Sandra J. Gendler; Mayo Clinic, Scottsdale, AZ) was expressed in the pCDNA3 vector (Invitrogen; San Diego, CA) and transfected by the lipofectin method (GIBCO BRL Life Technologies; Gaithersburg, MD) to U-251 MG cells. After 2 days of growth, the transfected cells were fixed with paraformaldehyde for study by immunofluorescence.
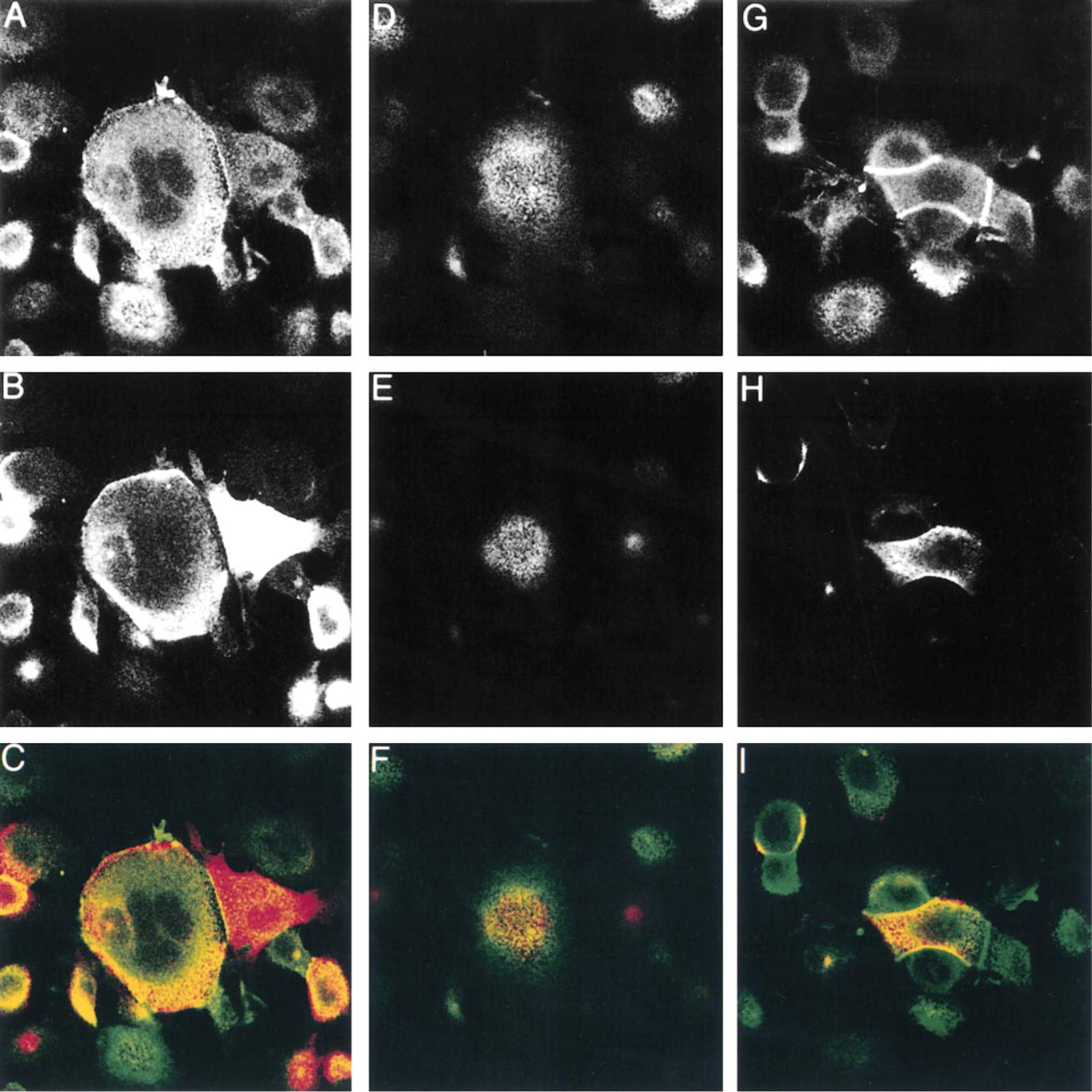
Partial co-localization of MUC1 and ezrin. Confocal microscopy was used to determine the degree of co-localization between MUC1 and ezrin. MCF-7az cells were double stained with MUSE11 (1:25 dilution) and Ez-Bl-III (1:50 dilution) antibodies and visualized with TRITC-conjugated donkey anti-mouse and FITC-conjugated swine anti-rabbit secondary antibodies (1:70 dilution), respectively. Pictures were taken from the basal (
Immunoelectron Microscopy
MCF-7az cells were grown on poly-
Immunoelectron Tomography
Electron micrographs of tilt series were collected with 3° increments from + 60 to –60 degrees. The resulting negatives were scanned with a Umax PowerLook 2000 scanner. Images were stored in JPEG format and aligned using our Jpeganim software (Engelhardt and Ruokolainen 1996; Engelhardt in press) for Silicon Graphics workstations (SGI). 3D reconstructions were produced from the aligned pictures by both weighted back projection and the maximum entropy method. The resultant volumes were examined with 3D viewing using BOB (public domain program; University of Minnesota, Graphic Visualization Laboratory, Army High Performance Computing Research Center), and animations for 3D visualization were produced using our FUNCS program for SGI computers (Engelhardt and Ruokolainen 1996; Engelhardt 2000).
Results
Localization of MUC1 and Ezrin to Similar Structures in MCF-7 Cells
A number of different cell lines were screened for endogenous expression of MUC1 and ezrin. We chose to study the localization of MUC1 and ezrin in MCF-7az cells because of the intense expression of MUC1. In double staining of cultured MCF-7az cells, both MUC1 and ezrin were seen in microvilli and in peripheral membrane areas, as earlier reported for MUC1 in mammary epithelial cells (Parry et al. 1990). The co-localization of MUC1 and ezrin was only partial in subconfluent cells (Figures 1 and 2). Typically, membrane areas with significant overlapping of the two proteins were seen, although the distribution differed somewhat, especially in the peripheral membrane areas. In addition, the MUC1-containing microvilli were located in smaller apical areas (topmost apical region) than ezrin-containing microvilli (Figures 1E and 1F). When the cells became spread and flattened, MUC1 and ezrin were largely localized similarly in the microvilli (Figure 2). MUC1 could also be seen in some cells in the cell–cell contact region, but not as typically as ezrin (Figures 1 and 2). The localization of ezrin to the cell–cell contact region could be detected by both polyclonal (Ez-Bl-III) and monoclonal (3C12) antibodies to ezrin (not shown). When U251-MG glioma cells, which do not produce endogenous MUC1, were transfected with MUC1 cDNA, a widely dispersed co-localization of ezrin and MUC1 was seen in the microvilli (Figure 3). In addition, in MCF-7az cells vacuoles staining for both MUC1 and ezrin were seen (Figure 2, insets). Some vacuoles stained intensely for both MUC1 and ezrin, but vacuoles staining only for ezrin were also seen (not shown). These structures might represent the intercellular lumina or vacuolar apical compartments (VACs) described in the literature (Vega-Salas et al. 1987). Almost without exception, vacuoles that stained for MUC1 also strained for ezrin.
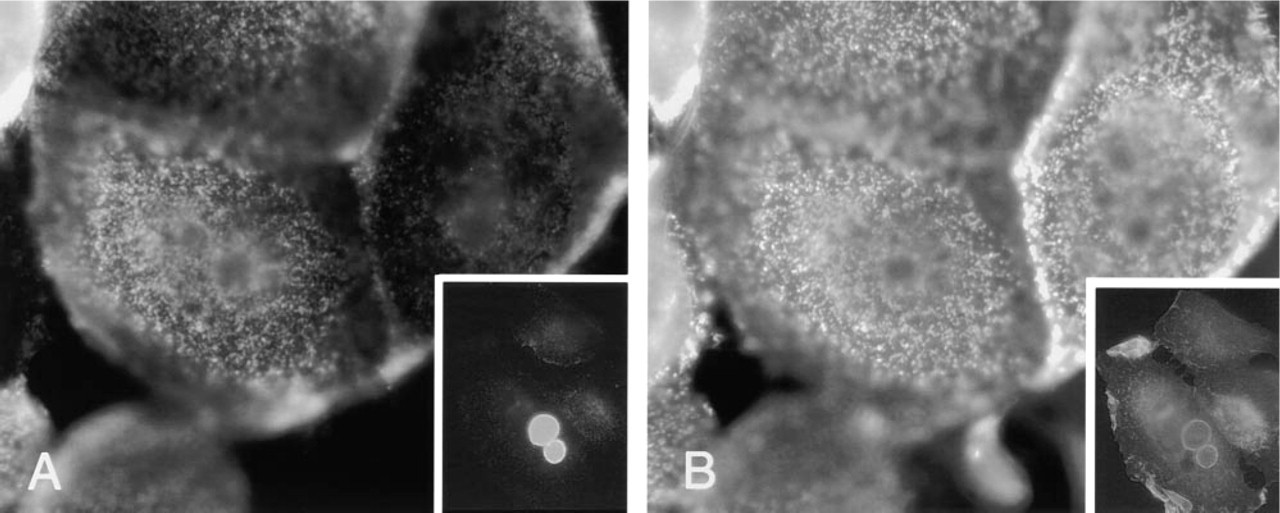
Conventional immunofluorescence photographs of MUC1 and ezrin in MCF7az cells. Confluent MCF-7az cells were double stained for MUC1 (
Transfection experiments of U251-MG glioma cells. Transfected cells were stained for MUC1 (
Localization of MUC1 in the Tips of Filopodial Structures
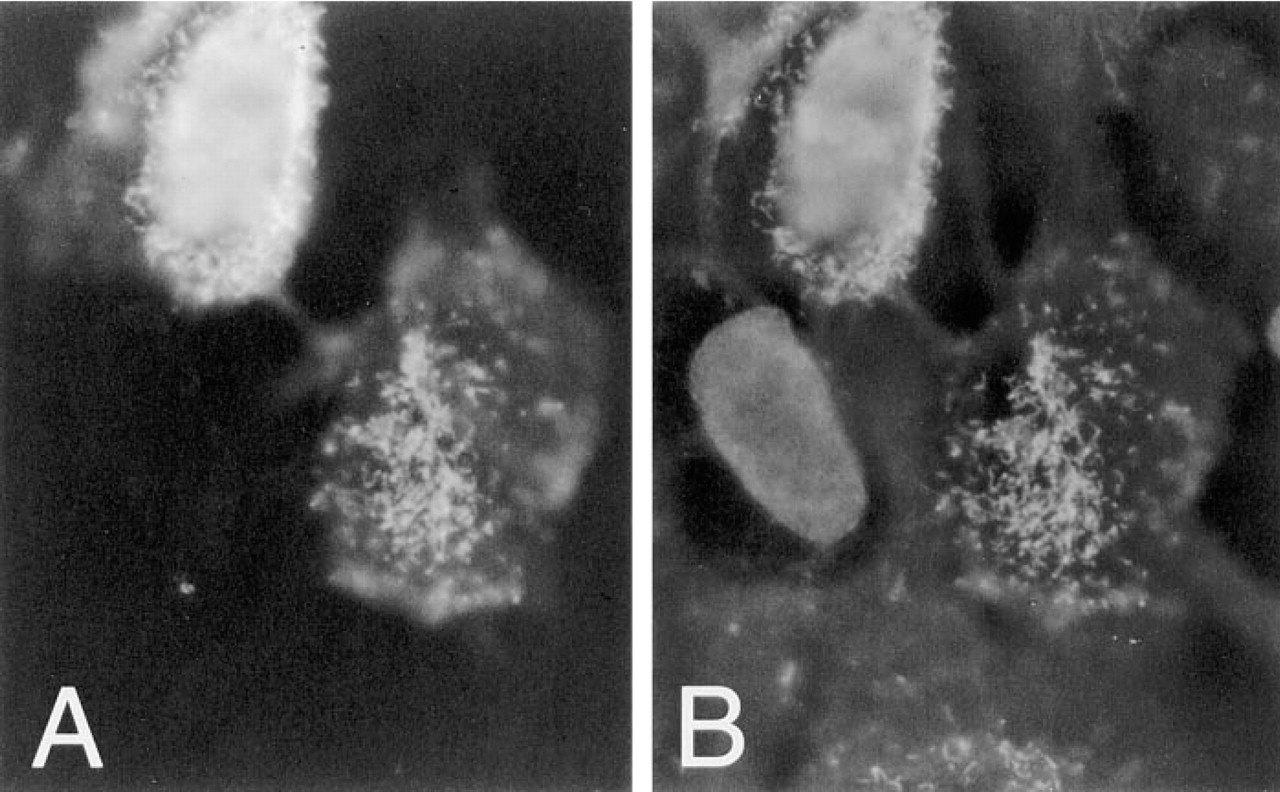
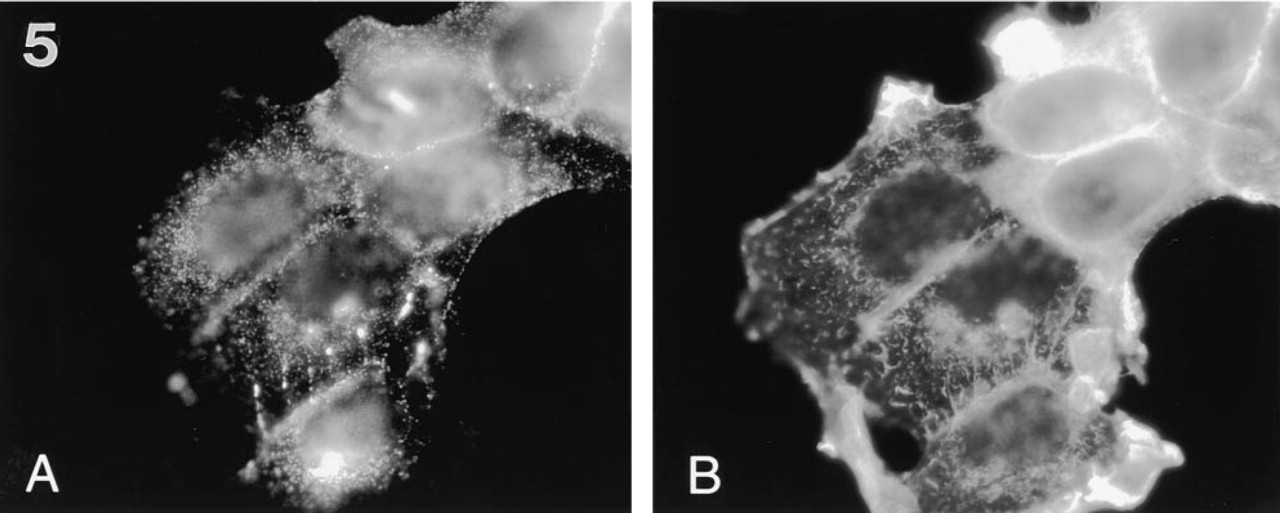
Filopodium-like protrusions that were dominated by MUC1 staining over ezrin staining were seen (Figure 4). The filopodial protrusions, which projected from the cells in the plane of the glass coverslip, resided mainly between the cells in cell clusters (Figure 4). In most cases the location of filopodial protrusions was polarized; they were pointing in the same direction as the leading lamella. The filopodial protrusions were seen only in subconfluent cells, suggesting that they are involved in cell locomotion. These filopodial protrusions were drumstick-like in appearance, containing a narrow stalk and a broader headpiece staining intensely with MUSE11 antibody to MUC1. In double staining experiments, the tip regions of the filopodial protrusions contained small amounts of ezrin (Figure 4B), whereas F-actin was found mostly along the stalk (Figure 5B). The presence of F-actin was monitored by staining with rhodamine-labeled phalloidin. The tips of the filopodial protrusions were located on or close to the basal cell surface, probably attached to the bottom of the growth plate (not shown). The stalk was not always found on the basal surface. The results were controlled by omitting the primary antibodies.
Immunoelectron Tomography
The localization of MUC1 was studied in whole-mounted microvilli and filopodial structures of MCF-7az cells by immunoelectron tomography. Immunoelectron tomography was used because it allows an unparalleled spatial resolution and provides ultrastructural detail about the specimen under inspection. 3D reconstruction enabled us to distinguish between true co-localization and apparent co-localization caused by superimposing structures. The depth of focus in an electron microscope is quite large, and 3D immunoelectron tomography enabled us to view the microvilli from all possible angles. The microvilli, which mostly projected towards the electron beam, were best visualized in specimen grids in which the Formvar membrane had been intentionally shattered, producing an array of cells in different spatial orientations and, in some cases, leading to an unobstructed view of a microvillus with no superimposing structures or Formvar membranes. Ezrin expression was monitored by 10-nm gold particles coupled to the secondary antibody detecting the polyclonal Ez-Bl-III antibody to ezrin. MUC1 expression was monitored by 5-nm gold particle labeling coupled to the secondary antibody detecting the monoclonal MUSE11 antibody to MUC1. The results were also tested by switching the secondary antibodies between MUC1 and ezrin. Omitting the primary antibody tested the specificity of binding. In these control experiments, binding of gold particles to the specimens was not seen. In addition, 1.4-nm gold particles were used to control for differences in permeability between 10-nm and 5-nm gold particles, but the visualization of 1.4-nm gold particles proved to be very difficult and is not discussed here.
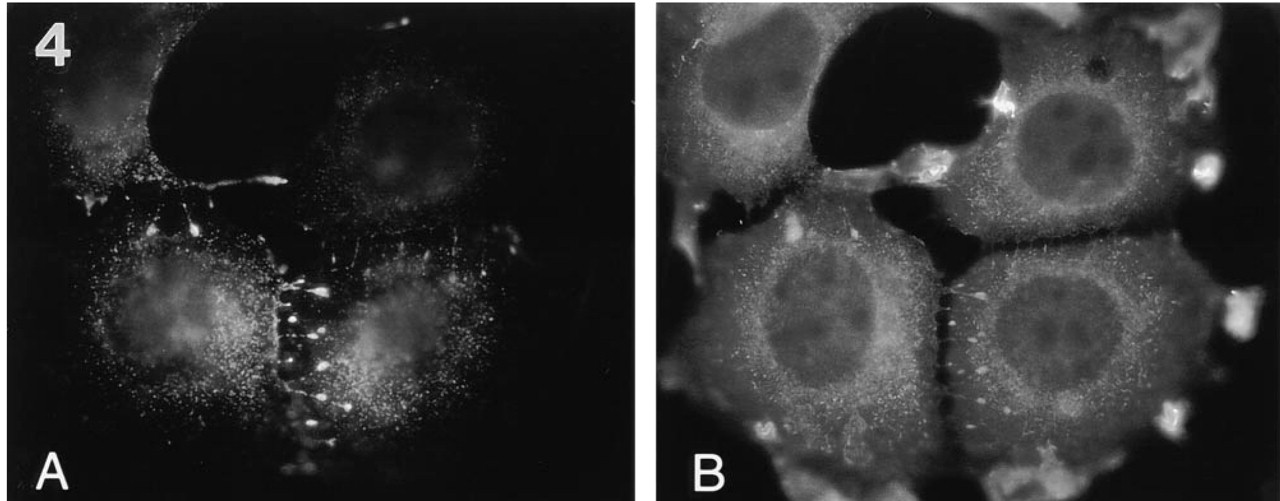
Typical MUC1-containing filopodial protrusions in MCF-7az cells. The cells were stained with MUSE11 antibody (1:25 dilution) (
Localization of MUC1 (
As can be seen in Figures 6 and 8, the similar localization of MUC1 and ezrin is also evident in the immunoelectron tomography pictures. In Figure 6, ezrin, depicted by 5-nm gold particles, is localized to the same patches as MUC1, which is labeled by 10-nm gold particles. In contrast when staining for ezrin and actin was performed a different pattern emerged (Figure 7). As before, ezrin was seen at certain patches along the length of the depicted microvillus. In contrast, actin was seen more widely dispersed along the entire length of the structure. This is congruent with the model proposed for the general structure of microvilli. Furthermore, in 3D immunoelectron tomography, 10-nm gold particles staining for ezrin were seen close to the cell membrane, whereas 5-nm gold particles representing actin were seen closer to the microvillar core. This supports the proposed model for the role and function of ezrin and actin in microvilli.
In a more detailed view of a filopodial structure, the similar localization of ezrin and MUC1 seen in immunofluorescence pictures was also confirmed. Ezrin, labeled by 10-nm gold particles (Figure 8), was distributed along the length of the filopodial structure, accompanied by a cluster of gold-conjugated antibodies representing MUC1 (5-nm gold particles in Figure 8).
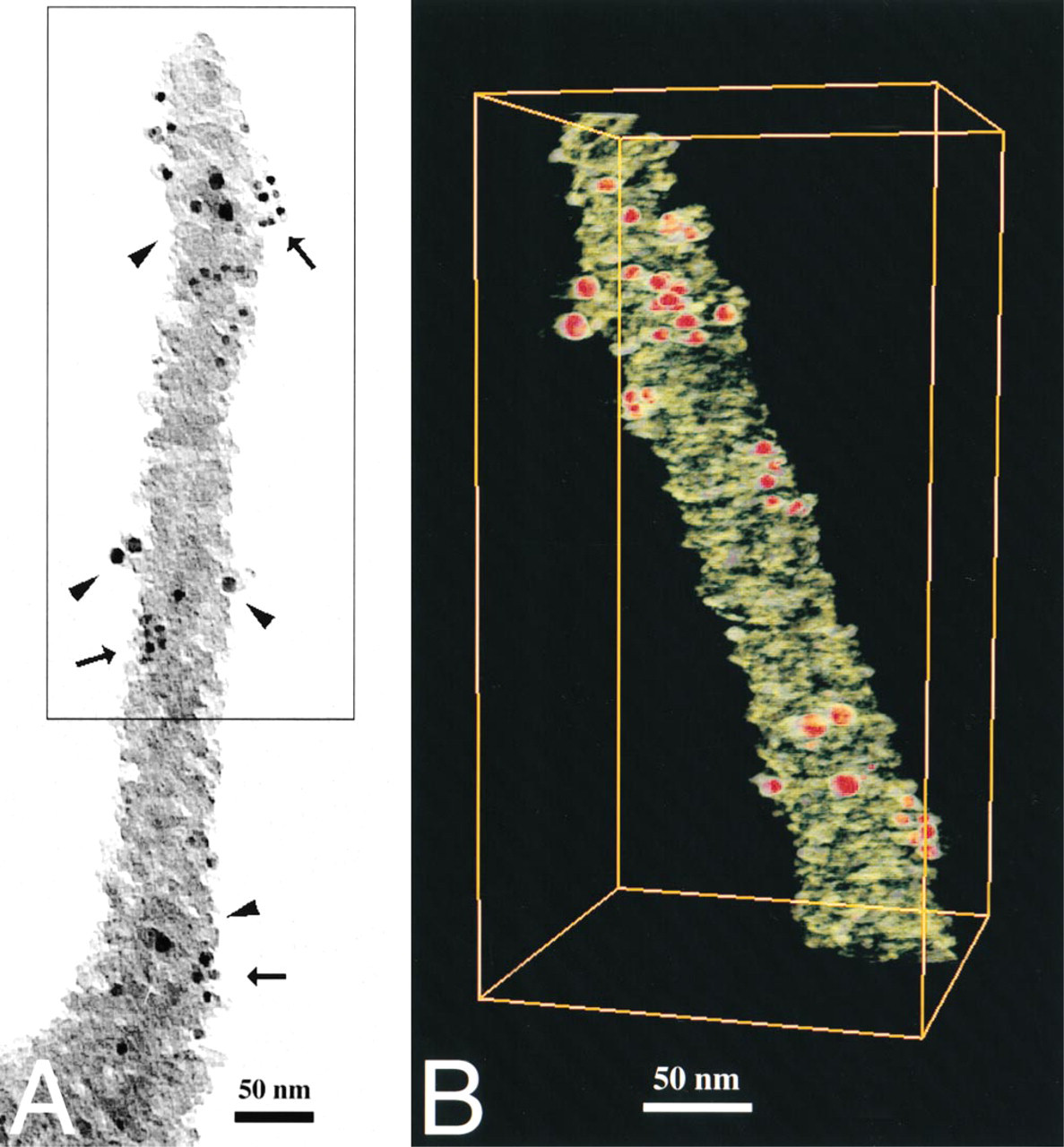
Immunoelectron microscopy and tomography of MUC1 and ezrin in a microvillus. The specimen grid was double stained with MUSE11 (MUC1) and Ez-Bl-III (ezrin) antibodies. For these experiments, the secondary antibodies corresponding to MUC1 and ezrin were switched around. Ezrin expression was monitored by 5-nm gold particles coupled to the secondary antibody detecting the polyclonal Ez-Bl-III antibody to ezrin. MUC1 expression was monitored by 10-nm gold particles coupled to the secondary antibody detecting the monoclonal MUSE11 antibody to MUC1. The localization of the 5-nm particles (arrows in

Immunoelectron tomography of ezrin and actin in a microvillus. The specimen grid was double stained with Ez-Bl-III antibody and monoclonal anti-actin antibody. Ezrin expression was monitored by 10-nm gold particles coupled to the secondary antibody detecting the polyclonal Ez-Bl-III antibody to ezrin, whereas actin is shown by 5-nm gold particles coupled to the secondary antibody detecting the monoclonal anti-actin antibody. Actin (arrows) can be seen along the length of the microvillus, whereas ezrin (arrowheads) is restricted to similar patches along the microvillus, as seen in Figure 6.
Discussion
We have shown that MUC1 and ezrin are largely localized together in microvilli and, to some degree, to the same cortical cell structures and to filopodia. The localization of ezrin and MUC1 to microvilli, in the entire apical surface, is seen clearly in flattened cells. However, there are also marked differences between MUC1 and ezrin in the distribution patterns. In sub-confluent cells, the localization of MUC1 is more restricted than the localization of ezrin, which is seen over the entire apical cell surface. Often, MUC1 showed a stronger concentration than ezrin at the topmost apical surface.
In immunofluorescence staining, ezrin is found at the cell–cell contact region. However, the presence of ezrin at the cell–cell contact region is controversial (Bretscher et al. 1997). It is possible that ezrin is located in microvillus-like adhesive structures at the cell–cell contact region. MUC1 was also occasionally seen at cell–cell centact sites, in a manner similar to ezrin, but the significance of this finding remains elusive.
The typical localization of MUC1 is its presence in long filopodial protrusions, and a strong MUC1 concentration at the tips of these filopodial structures was observed. The tips formed enlarged structures, which also contained ezrin, whereas F-actin was seen in the stalk. The amount of ezrin did not correspond to the amount of MUC1. This may be due to the intense signal produced by the binding of MUC1 antibody to the extracellular repeats of the MUC1 glycoprotein. The MUC1 epitope detected by MUSE11 antibody is located in tandem repeats on the extracellular side of the membrane (Hinoda et al. 1993). One MUC1 molecule can therefore bind several antibody complexes. This probably explains the presence of 5-nm gold particles in clusters of, on average, 10 particles in the whole-mounted specimens (Figure 8). An interesting finding was that 5-nm gold particle clusters (MUC1) were often accompanied by a single 10-nm particle (ezrin), suggesting the presence of a single ezrin molecule close to MUC1 (Figure 8). This may indicate that ezrin and MUC1 have spatial associations with one another in the cell surface protrusions. This spatial association can also be seen in Figure 6, in which MUC1 is represented by 10-nm gold particles and ezrin by 5-nm gold particles. Other types of filopodial protrusions were also seen, which did not contain any enlarged tip structures and clearly had F-actin along the length of the filopodium (not shown). MUC1 was not seen in these structures.
It has been shown that MUC1 contains two discrete motifs that determine its apical localization. The first is located in the extracellular domain and the second is the Cys-Gln-Cys motif at the junction of the cytoplasmic and transmembrane domains (Pemberton et al. 1996). There is evidence that MUC1 interacts with the cytoskeleton, as shown by disruption of actin filaments with cytochalasin D (Parry et al. 1990). However, it is not known if the cytoskeleton regulates both motifs that determine the apical localization of MUC1. One possible link of MUC1 with the cytoskeleton may be via interaction with catenins. MUC1 is reported to interact with the cell-cell adhesion junction proteins β-catenin and plakoglobin (Yamamoto et al. 1997), which normally interact with E-cadherin, a strong cell adhesion protein. The interaction of MUC1 with catenins also suggests that, in certain situations, MUC1 might replace other cell-adhesion proteins.
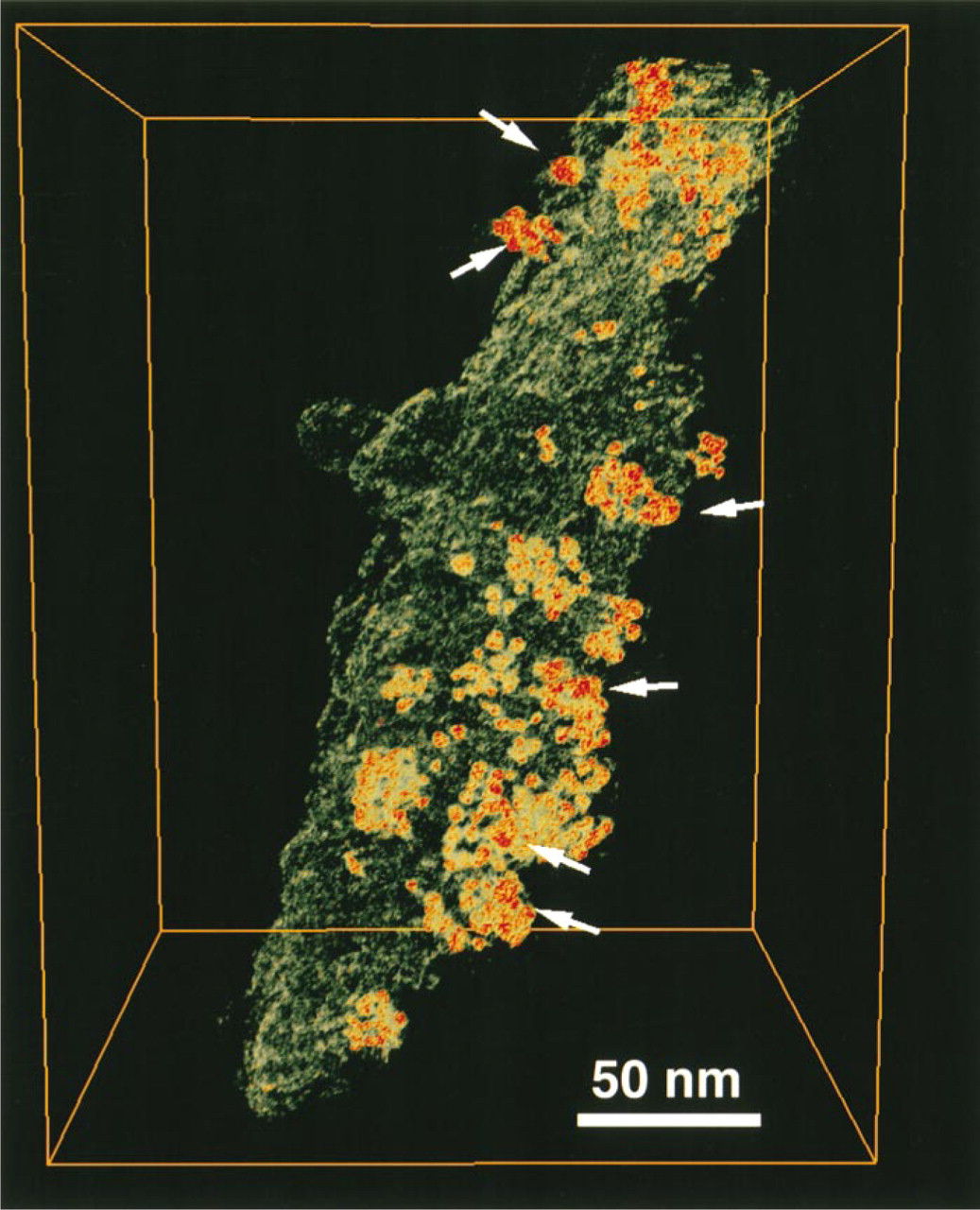
Immunoelectron tomography of MUC1 and ezrin in a filopodial protrusion. The specimen grid was double stained with MUSE11 (MUC1) and Ez-Bl-III (ezrin) antibodies. Ezrin expression was monitored by 10-nm gold particles coupled to the secondary antibody detecting the polyclonal Ez-Bl-III antibody to ezrin. MUC1 expression was monitored by 5-nm gold particles coupled to the secondary antibody detecting the monoclonal MUSE11 antibody to MUC1. Some 10-nm gold particles detecting ezrin (arrows) co-localized with the clusters of 5-nm gold particles detecting MUC1.
In general, mucins are believed to have anti-adhesion properties (Kim et al. 1996). By virtue of their negatively charged filamentous protruding structure, they can sterically hinder cell aggregation (Wesseling et al. 1996; for review see Kim et al. 1996). On the other hand, mucins may be involved in transient cell adhesion, in which the strength of adhesion is weak (van der Merwe and Barclay 1994). Therefore, mucins, like MUC1, could act both as an anti-adhesion protein and an adhesion protein, depending on the circumstances. The localization of MUC1 to filopodial protrusions, which appear to reach from cell to cell in subconfluent cultures but are abolished when confluence is reached, suggests a role for MUC1 in cell–substratum adhesion of filopodial structures. Filopodia are believed to be transient structures that may not require extensive filamentous actin cytoskeleton for stabilization. In addition, filopodia are not usually involved in forming stable adhesion contacts (Furthmayr et al. 1992). The prominent localization of MUC1 at the filopodial protrusions in MCF-7az cells is consistent with the transient nature of filopodia, and suggests a role for MUC1 in transient cell structures of migrating cells.
There is no evidence, as yet, for a direct association of MUC1 and ezrin in low or physiological salt concentrations, as studied in affinity chromatography, im munoprecipitation experiments, or by the Biacore surface plasmon resonance system with GST fusion protein of the cytoplasmic tail of MUC1 and purified ezrin, or ezrin in cell lysates (not shown). However, the possibility that MUC1 would not associate directly or indirectly with ezrin in certain circumstances is not fully ruled out, e.g., after certain induction signals followed by modification or conformational change in the cytoplasmic domain of MUC1. EBP50 is a protein that appears to link ezrin to anion exchangers in epithelial cells (Reczek et al. 1997) and may be involved in mediating an interaction between MUC1 and ezrin. The juxtamembrane cytoplasmic domain of MUC1 contains a similar positively charged cluster that interacts with the ERM proteins in CD44, CD43 and ICAM-2 (Legg and Isacke 1998; Yonemura et al. 1998). In addition, the immunoelectron tomography of the whole-mounted microvilli (Figures 6 and 7) and filopodial structures (Figure 8) showed the localization of MUC1 and ezrin to these structures with a similar overall distribution pattern of the two proteins. More significantly, ezrin was often seen in close spatial association with MUC1. A 5-nm gold particle cluster, probably labeling a single MUC1 molecule, was accompanied by a single 10-nm gold particle labeling ezrin. This suggests a functional association between MUC1 and ezrin.
In vivo, MUC1 and ezrin have similar localizations in the epithelial tissues. Both proteins are located on the apical surface of the epithelial cells (Berryman et al. 1993; Pemberton et al. 1996; Seregni et al. 1997). In contrast, CD44, an adhesion molecule that interacts with ezrin, is not localized to the apical surface of mature epithelial cells (Isacke 1994). Because ezrin has a prominent localization to the apical surface, other transmembrane proteins are likely to interact with ezrin in mature epithelial cells. Because of the similar localization on the apical surface of epithelial cells, MUC1 and other mucins may be major proteins associated with ezrin.
Footnotes
Acknowledgments
Supported by RIMI/NCRR grant #P20 RR0 11-583 from the National Institutes of Health.
We thank Dr Sandra J. Gendler for the MUC1 cDNA, Dr Yuri Hinoda for the MUSE11 antibody, and Ms Xeuying Wang, Ms Lynn Sydnor, Ms Johnellia Jordan, and Ms Nikki Miller for technical assistance.