
Abstract
Histamine (HA), contained in the enterochromaffin-like (ECL) cells of the gastric mucosa in animals, plays an important role in gastric acid secretion, although methods for its exact morphological localization are still lacking. We used a pre-embedding indirect immunoperoxidase approach to define the fine structural localization of HA in rat oxyntic mucosa that was fixed with a glutaraldehyde-based fixative and HA monoclonal antibodies (MAbs AHA-1 and 2). Transmission electron microscopy showed that the peroxidase end-product not only was concentrated in the cores of cytoplasmic granules but also was distributed to a high degree in the cytoplasm peripheral to the granules of the ECL cells. These results suggest that in ECL cells HA is enzymatically synthesized in the cytoplasm, then is transported and stored in the cores of the granules before its release from the basal lamina. The present HA immunoelectron microscopic method with MAbs would be applicable more generally to the ultrastructural identification of HA-containing cells. (J Histochem Cytochem 47:1031–1038, 1999)
Histamine (HA) is a ubiquitous chemical mediator that acts as a regulator of gastric acid secretion, a modulator of inflammatory reactions, and a neurotransmitter (Kahlson and Rosengren 1968; Soll and Walsh 1979; Schwartz et al. 1980). Also recently known is the fact that HA has been implicated in several other functions, including cell growth and differentiation (Hakanson et al. 1976; Panula et al. 1984; Tilly et al. 1990; West 1990; Nissinen et al. 1992). Although there have been many investigations at the light microscopic level by Panula and co-workers (Panula et al. 1984, 1988; Nissinen et al. 1992; Nissinen and Panula 1993, 1995), only a few reports regarding immunoelectron microscopic study for HA are presently available (Login et al. 1992; Nissinen and Panula 1993). This is most likely due to the lack of an ultrastructural detection method for HA that can be applied generally.
A large number of argyrophilic endocrine cells, contained in the oxyntic glands of rat stomach, synthesize and store large amounts of HA (Hakanson et al. 1976). These cells, designated enterochromaffinlike (ECL) cells by Hakanson and Owman (1966, 1967, 1969), do not normally possess demonstrable amounts of serotonin (5-hydroxytryptamine; 5-HT) or exhibit chromaffin and argentaffin reactivity, as do the true enterochromaffin (argentaffin) cells located predominantly in the pyloric glands. Recent studies have provided evidence that the ECL cells are the only HA-containing epithelial endocrine cell type in rat gastric glands, as shown by autoradiographic studies with radioactive HA precursors (Rubin and Schwartz 1979) and experiments using a density centrifugation technique (Soll et al. 1981). These results also appear to be supported by the localization of histidine decarboxylase-like immunoreactivity (Kubota et al. 1984). Recently, Nissinen and Panula (1993) provided direct evidence for cellular localization of HA, in ECL cells only, by an immunoelectron microscopic study with polyclonal HA antiserum. One would expect the vesicular granules, characteristic of ECL cells, to store HA. However, in the subcellular examination in their study, HA was distributed diffusely in the cytoplasm but not in the granules. This might be a consequence of their method of fixing tissues with the carbodiimide-based solution in the immunoelectron microscopic study (Nissinen and Panula 1993). In their immunocy-tochemical studies, however, carbodiimide was an unavoidable tissue fixative, because the HA antiserum was produced against the carbodiimide-conjugated HA (Panula et al. 1988; Nissinen and Panula 1993). In general, in immunocytochemical studies of small molecules, the best results can be expected when the epitope of the hapten-protein conjugate used for immunization closely resembles the fixed tissue antigen because the antibody can precisely recognize not only the antigen structure but also, in part, crosslinking sites with carrier proteins (Buijs et al. 1984; Onteniente et al. 1984; Seguela et al. 1984; Panula et al. 1988; Fujiwara et al. 1994; Fujiwara and Masuyama 1995).
In our recent study of monoclonal antibodies (MAbs AHA-1–5) against glutaraldehyde-conjugated HA, we identified two MAbs (AHA-1 and 2) that gave the best immunocytochemical results at the light microscopic level when glutaraldehyde was used as a tissue fixative (Fujiwara et al. 1997). These successes prompted us to further examine the possibility of the use of immunoelectron microscopy for precise cellular localization of HA in rat oxyntic glands with these MAbs.
Materials and Methods
Antibodies
The MAbs AHA-1 and 2 (IgG1 subisotype) were produced against HA conjugated to bovine serum albumin (BSA) using glutaraldehyde (GA) and sodium borohydride (Fujiwara et al. 1997). The specificity of the immunocytochemical method that utilized MAb AHA-1 has been demonstrated to be specific to HA and not reactive to any of the amino acids and peptides with N-terminal histidine (Fujiwara et al. 1997). In HA immunocytochemistry, the immunoreaction was completely abolished by preabsorption of the MAb with GA-conjugated HA but not with GA-conjugated L-histidine.
Animals
Normal adult male Wistar rats (Otsubo Experimental Animals; Nagasaki, Japan), body weight 200–250 g, were used in this study. They were housed in temperature- and light-controlled rooms (21 ± 1C and 12L:12D) and had free access to standard food and tapwater.
Immunocytochemistry (ICC)
Under sodium pentobarbital (Abbott Labs; North Chicago, IL) anesthesia (60 mg/kg), 10 rats were perfused intracardially with PBS at 50 ml/min for 2 min at room temperature (RT) and then with a freshly prepared solution of 2.5% GA in 10 mM phosphate buffer, pH 7.2, for 6 min. Stomachs were quickly excised and immersed overnight at 4C in the same fixative. They were cut into 50-μm-thick sections with a Microslicer (Dosaka EM; Kyoto, Japan) and the sections were treated for 10 min with 1.0% H2O2 to inhibit endogenous peroxidase, 0.2% NaBH4 for 10 min, and incubated at 4C for 48 hr with primary MAb AHA-1 or -2 at concentrations ranging from 10 to 50 ng/ml of 50 mM Tris-HCl buffer, pH 7.4, containing 0.86% NaCl (TBS) [the concentration of which was determined by the conventional sandwich ELISA using chromatographically purified mouse IgG (Zymed; San Francisco, CA) as a standard]. Sections were then incubated with a goat anti-mouse IgG/Fab' labeled with horseradish peroxidase (HRP) (MBL; Nagoya, Japan) 1:200 for 12 hr at 4C. After rinsing with TBS, the HRP was revealed for 5–10 min with diaminobenzidine and H2O2. The HRP substrate consisted of 10 mg of 3, 3′-diaminobenzidine tetrachloride (Sigma; St Louis, MO) dissolved in 20 ml of 50 mM Tris buffer, pH 7.4, supplemented with 20 of 30% H2O2 (Graham and Karnovsky 1966). The samples were rinsed first with Tris-HCl buffer, pH 7.4, and then with cacodylate buffer, pH 7.4, for 30 min at RT.
Electron Microscopy
The color-developed specimens prepared as above were then fixed with 1.0% osmium tetroxide in 50 mM cacodylate buffer, pH 7.4, for 1 hr at RT and dehydrated in a series of graded ethanol solutions. After immersion in propylene oxide (Nacalai Tesque; Kyoto, Japan) (three times for 10 min each), the samples were immersed in a mixture (1:1) of propylene oxide and Epon 812 resin (Taab; Reading, UK) overnight and embedded in Epon 812 resin in a routine manner. The regions to be studied were cut with a 2-mm diameter punch, mounted on Epon blocks, and sectioned on a horizontal plane into ultrathin sections, which were then immediately observed in a 100 CX Jeol electron microscope.
For quantification, images of electron microscopic photographs (both sample and control specimens) taken under constant conditions were digitized into a computer through a video-camera head (ITC-370M; Olympus-Ikegami, Tokyo, Japan) according to the method of Shin et al. (1995). The optical density of the cores of the granules, but not including the light halo, the mitochondria, and the nucleus in the ECL cells of the photographs, was measured by a computer-assisted image analyzer (Olympus-SP500; Olympus, Tokyo, Japan) under constant conditions.
Control Experiments
In the HA immunocytochemistry study, the specificity of immunostaining was ascertained by incubating control sections with (a) the secondary antiserum alone, (b) type-matched MAb (IgG1) anti-penicillin (30–80 ng/ml; Cosmo, Tokyo, Japan), and (c) MAb AHA-1 or -2 preabsorbed with histamine-glutaraldehyde-human serum albumin (HA-GA-HSA) conjugate at a concentration of 2 μg/ml.
Results
Immunoelectron microscopic study of HA was carried out employing a pre-embedding method using 50-μm Microslicer sections of the rat gastric fundus and employing MAb AHA-1 or -2 produced against glutaraldehyde-conjugated HA (Fujiwara et al. 1997). Both MAbs AHA-1 and -2 produced intense immunostaining of certain cells of the rat stomach when the tissues were fixed with 2.5-5.0% glutaraldehyde (GA), followed by reduction of tissue sections by NaBH4, this being in agreement with the results in the previous study using paraffin sections (Fujiwara et al. 1997). However, tissue fixation with a mixture of 2.5% GA plus 4% paraformaldehyde (PFA) combined with the tissue reduction resulted in only moderate immunostaining in the cells by MAb AHA-2, and none at all by MAb AHA-1. Furthermore, neither MAb AHA-1 nor MAb 2 produced any immunostaining in the cells when 5% carbodiimide, 5% formalin, or 4% para-formaldehyde was used as a tissue fixative, as shown in our previous study (Fujiwara et al. 1997).
HA immunoreactivity was detected by light microscopy mainly in the cells scattered in the basal half of the gastric glands (oxyntic glands) of the gastric fundus and corpus, and in a few cells in the neck portion of the glands (Figure 1A). The HA-immunoreactive cells varied in shape and size and often had one or two processes invading the bases of the adjacent cells. However, no HA-immunoreactive cells were detected in other areas of the stomach, such as pyloric glands, antrum, or other parts of the gastric glands. The mast cells around the blood vessels in the submucosa were very weakly stained by the MAb (Figure 1A), this being in agreement with previous results reported by Login et al. (1992) and us (Fujiwara et al. 1997). No immunoreaction occurred with the MAb AHA-1 or -2 preabsorbed with HA-GA-HSA conjugates (2 μg/ml) (Figure 1C), with anti-penicillin MAb, or with the secondary antiserum alone.
Immunoelectron Microscopy
Among endocrine cells in rat gastric glands, ECL cells and A-like cells were predominantly observed, most of which in the basal half of the mucosa were in contact with the basal lamina of the glands (Figures 2A and 3A). Immunoelectron microscopic examination of the basal parts of the gastric glands showed identical results for MAb AHA-1 and MAb AHA-2: predominant distribution of HA in the ECL cells, which were identified on the basis of their characteristic ultrastructural features; many vesicular granules located in the basal part of the cell, varying in size within the same cell; and a wide electron-lucent halo separating the osmophilic core from the limiting membrane (Forssmann et al. 1969). Some of the labeled cells in the neck portion of the gastric glands were also examined by electron microscopy, revealing that the cells tested were all ECL cells, not intraepithelial mast cells (not shown). There was a marked tendency for higher immunoreactivity in the specimens fixed with GA alone than with a mixture of 2.5% GA plus 4% PFA. On the other hand, better morphology was apparent with a mixed fixative than with GA alone. All 65 of the ECL cells and 50 of those found in the specimens were immunoreactive with MAb AHA-1 and MAb 2, respectively (Figures 2A and 2B).
Immunoreactive end deposits were highly concentrated in the cores of various sizes of granules (100-500-nm diameter). They were also diffusely distributed throughout the cytoplasm, especially near the surroundings of the ECL cell granules (Figure 3A). In addition, the processes (cytoplasm) characteristic of the ECL cells were positive for HA immunoreactivity (Figure 2B) and were not seen to make contact with the lumen of the gland. However, the nuclei and mitochondria were devoid of immunoreactivity (Figures 2A, 2B, and 3A). The sections stained with the preabsorbed MAb AHA-2 or the anti-penicillin MAb did not show any HA immunoreactivity (Figure 3B). A computer-assisted image analysis of the electron microscopic photographs (Figures 3A and 3B) was carried out by measuring the optical density of the cores of the granules and mitochondria in the ECL cells (Figure 4). The values obtained for the cores of the granules in the sample and control specimens were 184.4 ± 28.3 (SE) and 91 ± 16.4, respectively, with a statistically significant difference (p <0.001), revealing that the cores of the granules were significantly labeled (Figure 4). In contrast, the mitochondria were unlabeled, because no significant difference (p >0.05) was evident between the sample (127.1 ± 8.1) and control (113.7 ± 12.9) (Figure 4). In addition, A-like cells in the gastric glands were analyzed in the same manner, and we found no labeling of the granules in the cells of sample specimens (not shown). None of the other endocrine cells occasionally detected in the specimens, such as D-, P-, and D1-cells, were immunoreactive with both the MAbs (not shown).
Discussion
Nissinen and Panula (1993) have recently reported an immunoelectron microscopic study of HA in rat gastric glands using polyclonal rabbit HA antibodies produced against the carbodiimide-conjugated HA and also using the carbodiimide-based fixative containing both paraformaldehyde (PFA) and a low concentration of GA. The authors raised the concern that subcellular storage of HA may be sensitive to change during sample processing because of the low molecular weight of HA, leakage from the vesicles, or diffusion of the reaction product (Nissinen and Panula 1993). In their study, however, HA immunoreactivity was not at all evident in the cytoplasmic granules, although it was diffusely distributed throughout the cytoplasm of the ECL cells. GA-based fixation is known to strongly crosslink and immobilize reactive molecules in their native compartments and to be superior to carbodiimide-based fixation for preservation of morphology for light and electron microscopic studies (Pearse 1980; Storm-Mathisen et al. 1983; Geffard et al. 1984; Seguela et al. 1984; Hodgson et al. 1985; Campistron et al. 1986; Ottersen et al. 1986; Fujiwara et al. 1998). Furthermore, an MAb allows the detection of the cellular pools of the antigens more precisely than could be achieved using polyclonal antiserum (Guesdon et al. 1986; Madl et al. 1986; Meyer et al. 1986; Chagnaud et al. 1987; Decavel et al. 1987; Williams and Beitz 1989; Frandoza et al. 1990). The best results in HA immunocytochemistry with the HA MAbs produced in this laboratory can be expected when a GA-based fixative is used (Fujiwara et al. 1997). In the production of the HA MAbs, the HA-protein conjugate used for immunization was prepared with GA (Fujiwara et al. 1997).
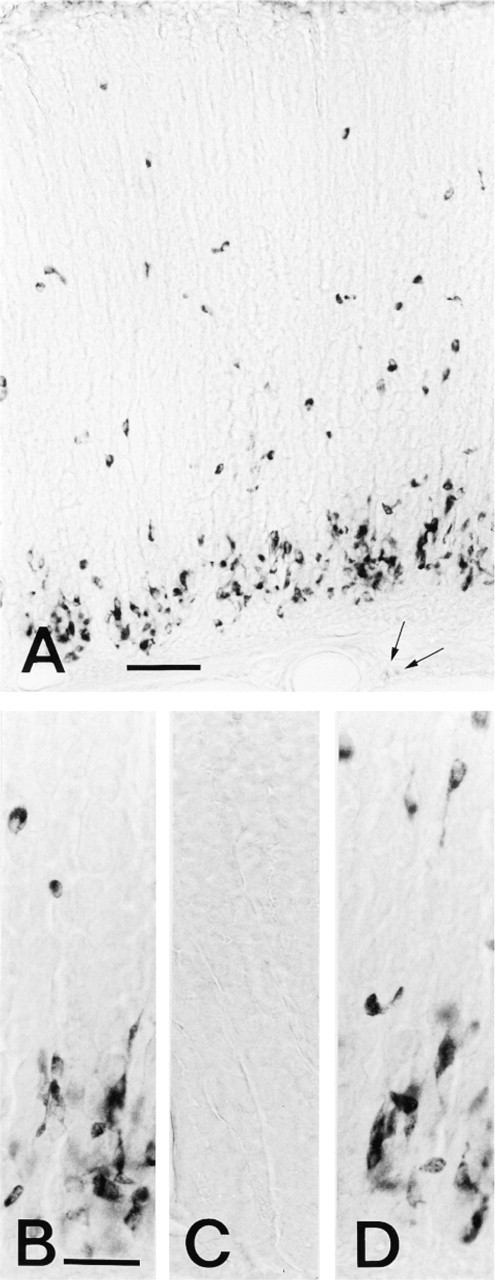
Rat gastric mucosa stained for HA by immunocytochemistry. (A) Section of the corpus of rat stomach stained by MAb AHA-2 (by the indirect method described in Materials and Methods). Strong immunostaining occurs in the cytoplasm of the cells scattered in half of the glands of the mucosa, but very weak staining in mast cells (arrows) of the submucosa. Bar = 0.1 mm. (B-D) Sections of the gastric mucosa reacted with (B) MAb AHA-1, (C) MAb AHA-2 preabsorbed with glutaraldehyde-conjugated HA (HA-GA-HSA, 2 μg/ml), and (D) MAb AHA-2 incubated with L-histidine-GA-HSA conjugate (20 μg/ml). A staining pattern similar to that of MAb AHA-2 was seen (B). HA conjugate completely abolished all staining (C), but L-histidine conjugate does not affect the staining (D). Bars = 50 μm.
In the present immunoelectron microscopic study of the subcellular localization of HA in cells of rat gastric glands, using the HA MAbs and GA-based fixation, HA immunoreactivity was demonstrated only in the ECL cells among epithelial endocrine cell types. This agrees with previous results (Rubin and Schwartz 1979; Soll et al. 1981; Nissinen and Panula 1993) and also with those of localization of histidine decarboxylase-like immunoreactivity (Kubota et al. 1984). Intense HA immunoreactivity was observed in the present immunocytochemistry when MAb AHA-1 or MAb 2 and a fixative of GA were used in combination. In the case of a mixed fixative of 2.5% GA plus 4% PFA, however, the degree of HA immunoreactivity decreased slightly with MAb AHA-2 but disappeared with MAb AHA-1. These phenomena might be due to a change in the HA antigenicity, resulting from a possible chemical reaction of HA in tissues with PFA contained in the mixed fixative, resultant HA derivative(s) of which may be detected by crossreaction of MAb AHA-2, but not by MAb AHA-1. It has already been reported that HA reacts with formalin, yielding a certain compound with weak fluorescence (Falck et al. 1962), and in our previous study MAb AHA-2 showed a much higher affinity to HA-protein conjugates than MAb AHA-1, as assayed by ELISA, although the two MAbs were both monospecific for HA (Fujiwara et al. 1997). In addition, in the samples evaluated subcellularly, fine histological structures were so evident that we easily determined that the immunoreactive end-products were located mostly in the cores of the granules and in the cytoplasm peripheral to the granules of ECL cells, whereas nuclei and mitochondria were devoid of immunoreactivity. These results therefore suggest that in ECL cells HA is synthesized enzymatically through L-histidine with L-histidine decarboxylase in the cytoplasm, and then is transported and stored in the cores of the granules before its release extraglandularly via the basal lamina. This may be supported by the results of Kubota et al. (1984), who revealed by immunocytochemistry, using an antiserum against L-histidine decarboxylase, that the enzyme was distributed diffusely throughout the cytoplasm but not in the granules of ECL cells. However, the possibility that the perigranular locations of HA might be a consequence of leakage from granules or diffusion of the core HA during immunocytochemistry could not be excluded because the present method still has limitations that lead to insufficient morphology for the secretory granules of ECL cells, in which the light halo structures around the cores appeared rather empty in the micrographs and the granule membranes were unclear (Figures 3A and 3B). Therefore, more definitive information on the ultrastructural sites of synthesis, storage, and release of HA is needed to postulate the intracellular pathways of HA in ECL cells.
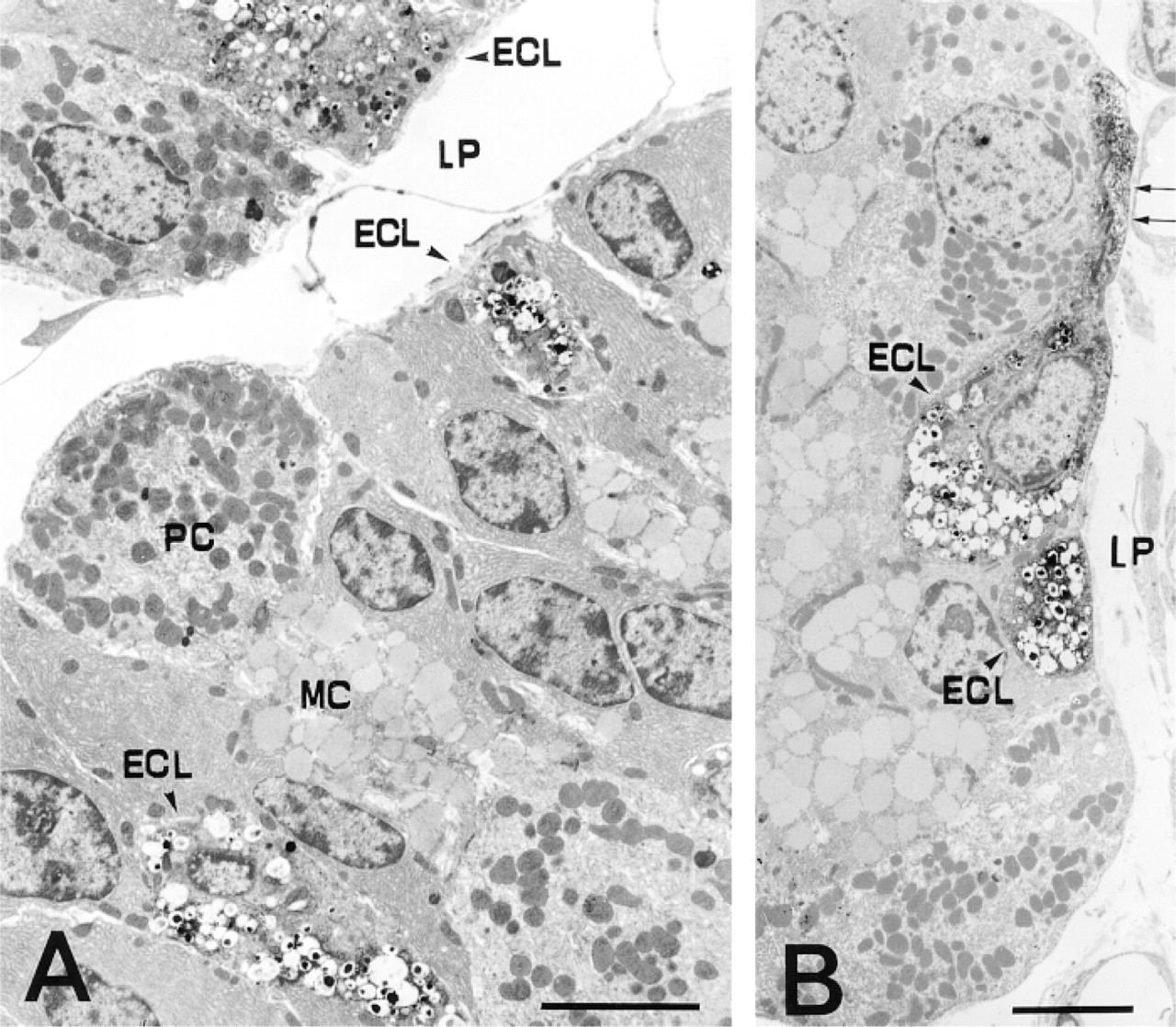
Immunoelectron micrographs of rat gastric mucosa reacted with MAb AHA-2 (A) and MAb AHA-1 (B). (A) Three ECL cells containing many characteristic granules with HA-immunopositive core matrix are seen. Other cells in the glands, such as the parietal cells and mucous cells, lack immunoreactivity. (B) One of the two ECL cells possesses a characteristic process (arrows), which was HA-immunoreactive in the cytoplasm, and the nucleus was nonimmunoreactive with the MAb. ECL, enterochromaffin-like cell; PC, parietal cell; MC, mucous cell; LP, lamina propria. Bars = 5 μm.
In this study, very weak staining or no staining at all was observed in mast cells in the epithelium and around the blood vessels in the submucosa, this being in agreement with the results of Login et al. (1992) and our results using paraffin sections (Fujiwara et al. 1997), although biochemical data show a relatively high concentration of HA in these cells (Login et al. 1987). The present fixation methods might be less able to preserve the association of HA in granules of the mast cells than in the ECL cells. Alternatively, it is possible that binding of the primary or secondary antibodies was impaired because of steric or charge interactions with constituents of the granule matrix of the mast cells (Rabenstein et al. 1987; Diamant 1990; Login et al. 1992).
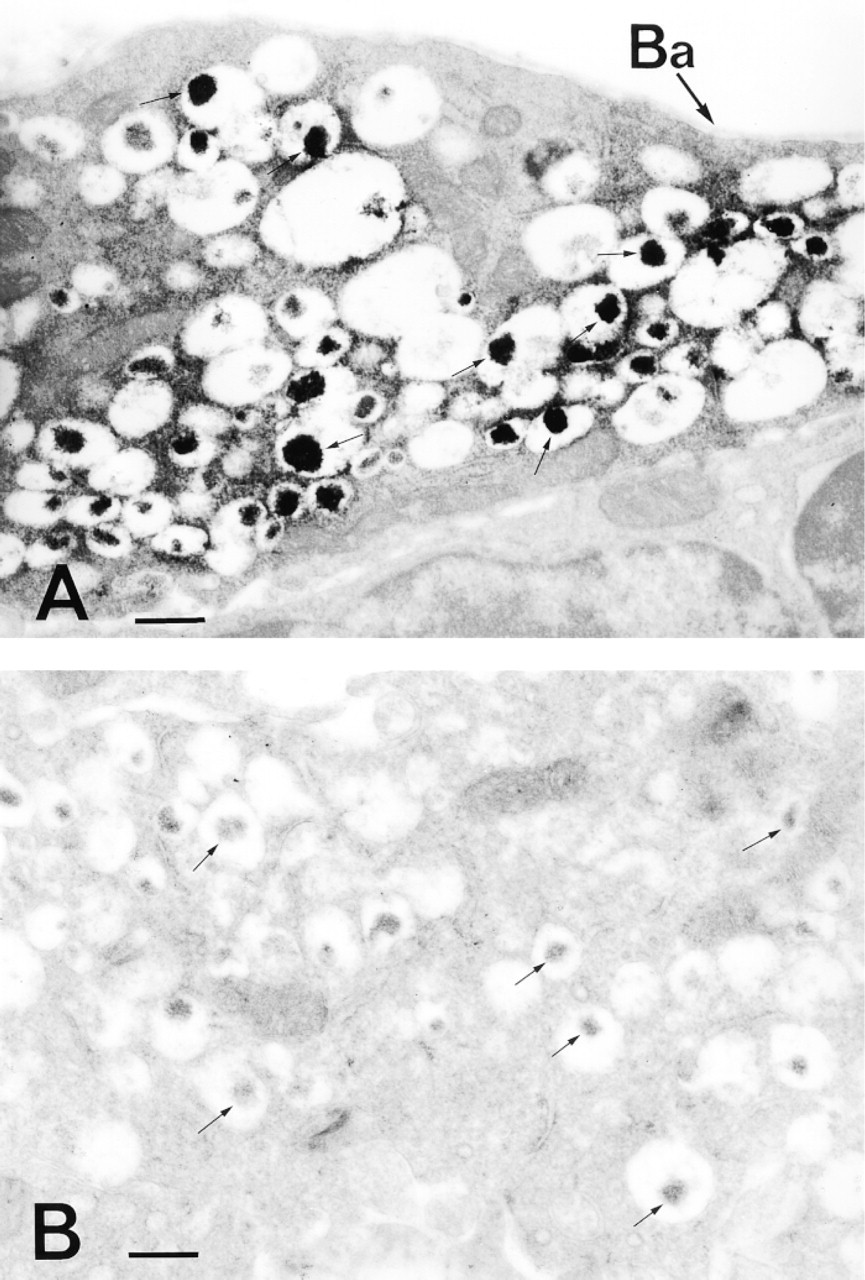
Immunocytochemical localization of HA in an ECL cell. (A) Strongly HA-immunoreactive structures were observed not only in the cores of the granules but also in the cytoplasm peripheral to the granules. The mitochondria lack immunoreactivity. (B) Control for immunocytochemical staining. Immunocytochemistry was carried out in the same way as in A, but HA-GA-HSA conjugate (2 μg/ml) was added to the MAb AHA-2 solution as a control. No labeling occurred in any of the subcellular organelles in the ECL cell. Arrows indicate the cores of the granules. Ba, basal lamina. Bars = 500 nm.
Although HA has been implicated in a variety of biologically important events, such as being the main mediator of allergic reactions and one of the neurotransmitters (Kahlson and Rosengren 1968; Soll and Walsh 1979; Schwartz et al. 1980), immunoelectron microscopic studies of HA have not been undertaken except for the limited studies of Nissinen and Panula (1993) and Login et al. (1992), who used commercial sources of anti-HA sera for subcellular localization of HA in rat peritoneal mast cells. In their study, however, a correlation between a crosslinking agent for HA antigen preparation for the antisera and the tissue fixative used in the immunocytochemistry is unclear. The present HA immunoelectron microscopic method with MAb AHA-1 or MAb AHA-2 allowed the detection of tissue HA antigens fixed with GA-based fixative. Because this fixative is commonly used in electron microscopic studies, this method should be more generally applicable to the ultrastructural identification of HA-containing cells.
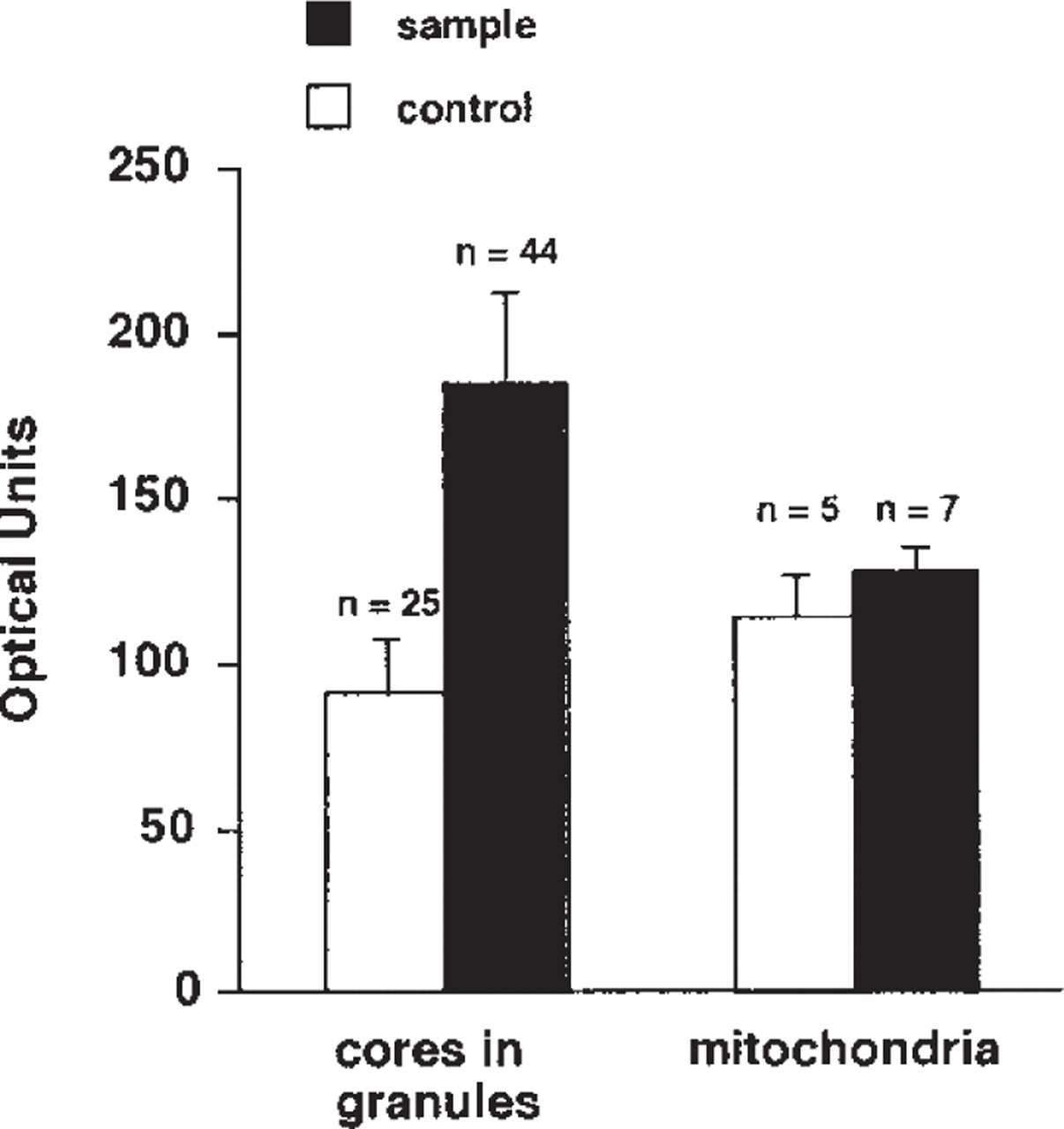
Relative intensity of the immunostaining and control specimens. The regions of the granule cores [44 in the sample specimen (Figure 3A) and 25 in the control (Figure 3B)], but not including the light halo, and in the mitochondria (seven in Figure 3A, five in Figure 3B) were analyzed and treated statistically.
Footnotes
Acknowledgments
We are very grateful to Dr M. Shin for the computer-assisted image analysis, Mr T. Suematsu for technical assistance with electron microscopy, and Prof M. Tiedemann for assistance with editing the manuscript.