
Abstract
The polyamines (PAs) are ubiquitous polycationic metabolites in eukaryotic and prokaryotic cells and are believed to be intimately involved in the regulation of DNA, RNA, and protein biosynthesis, the exact function of which remains unclear, mainly because of a lack of knowledge of PA subcellular localization. In this study, using immunoelectron microscopy, we have demonstrated that PAs are predominantly located on free and attached ribosomes of the rough endoplasmic reticulum in the neurons of the lateral reticular nucleus of rat medulla oblongata. The nuclei, axons, and nerve endings were devoid of PA. This suggests that PAs are one of the components of biologically active ribosomes, being closely involved in the translation processes of protein biosynthesis.
Keywords
The present study demonstrated that PAs are predominantly located in the ribosomes in the neurons of the central nervous system of rats. However, the possibility that PAs are located in sites other than the ribosomes in cells could not be excluded, because it is known that PAs have effects on membrane proteins such as ion channels and receptors (for reviews see Schuber 1989; Rock and Macdonald 1995; Williams 1997).
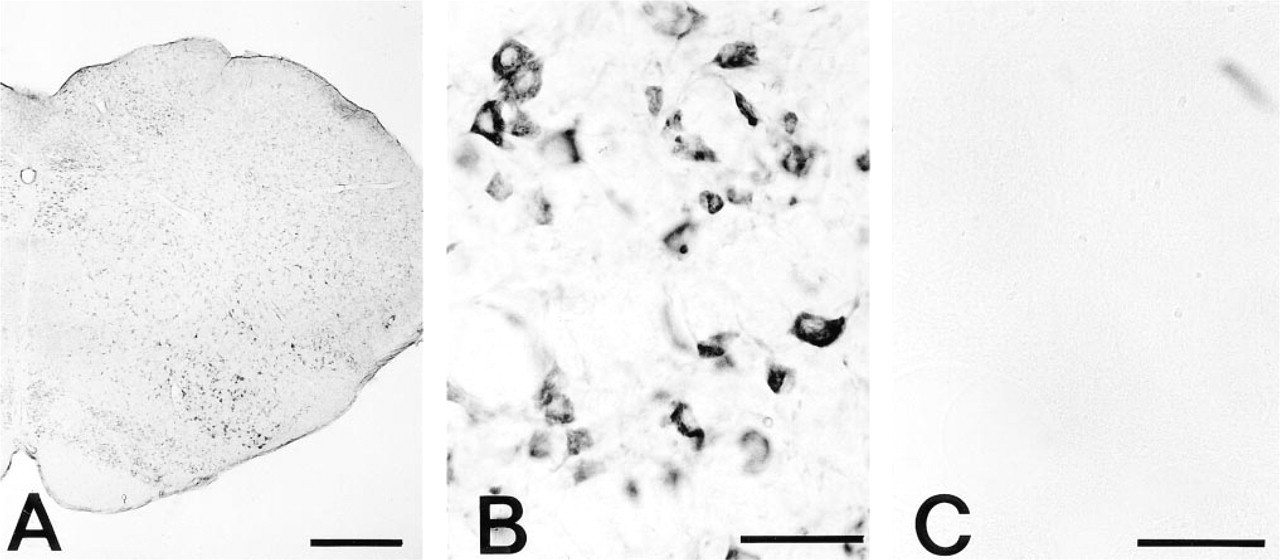
Rat medulla oblongata for PAs by an ICC method using MAb ASPM-29. (
Materials and Methods
Antibody
The monoclonal antibody (MAb) ASPM-29 (IgG1 subisotype) was produced against spermine conjugated to human serum albumin (HSA) using glutaraldehyde (GA) and sodium borohydride (Fujiwara and Masuyama 1995). The specificity of the immunocytochemical method that utilized the ASPM-29 MAb has been characterized extensively (Fujiwara and Masuyama 1995; Fujiwara et al. 1996,1997). In PA immunocytochemistry model experiments with an ELISA binding test, the ASPM-29 MAb detected Spm and Spd but not putrescine, cadaverine, 1,3-diaminopropane, N1, N12 -diacetylspermine, N-monoacetylspermine, N1, N8 -diacetylspermidine, N1 -acetyspermidine, N8 -acetylspermidine, N1, N4 -diacetylputrescine, N-monoacetylputrescine, ornithine, lysine, arginine, glycine, γ-aminobutyric acid, or histamine (Fujiwara and Masuyama 1995). In PA immunocytochemistry the immunoreaction was completely abolished by preabsorption of the MAb with either Spm or Spd but not with putrescine (Fujiwara and Masuyama 1995; Fujiwara et al. 1996,1997).
Animals
Normal adult male Wistar rats (Otsubo Experimental Animals; Nagasaki, Japan), body weight 200–250 g, were used in this study. They were housed in temperature- and lightcontrolled rooms (21 ± 1C and 12L:12D) and had free access to standard food and tapwater.
Immunocytochemistry (ICC)
Under sodium pentobarbital (Abbott Laboratories; North Chicago, IL) (60 mg/kg) anesthesia, 10 rats were perfused intracardially with PBS at 50 ml/min for 2 min at room temperature (RT), and then with a freshly prepared solution of 2.5% GA in 10 mM phosphate buffer at pH 7.2 for 6 min. Brains were quickly excised and immersed overnight at 4C in the same fixative. They were cut into 50-μm-thick sections with a Microslicer (Dosaka EM; Kyoto, Japan) through all levels of the lower parts of the medulla oblongata. The sections were treated with 0.2% NaBH4 for 10 min and incubated at 4C for 48 hr with primary MAb ASPM-29 at concentrations ranging from 10 to 50 ng/ml of 50 mM Tris-HCl buffer, pH 7.4, containing 0.86% NaCl (TBS) [the concentration of which was determined by the conventional “sandwich ELISA” using chromatographically purified mouse IgG (Zymed; San Francisco, CA) as a standard]. Sections were then incubated with a goat anti-mouse IgG/Fab' labeled with horseradish peroxidase (HRP) (MBL; Nagoya, Japan) 1:200 for 12 hr at 4C. After rinsing with TBS, the HRP was revealed for 5–10 min with diaminobenzidine and H2O2. The HRP substrate consisted of 10 mg of 3,3′-diaminobenzidine tetrachloride (Sigma; St Louis, MO) dissolved in 20 ml of 50 mM Tris buffer at pH 7.4 supplemented with 20 μl of 30% H2O2. The samples were rinsed first with Tris-HCl buffer, pH 7.4, and then with cacodylate buffer, pH 7.4, for 30 min at RT.
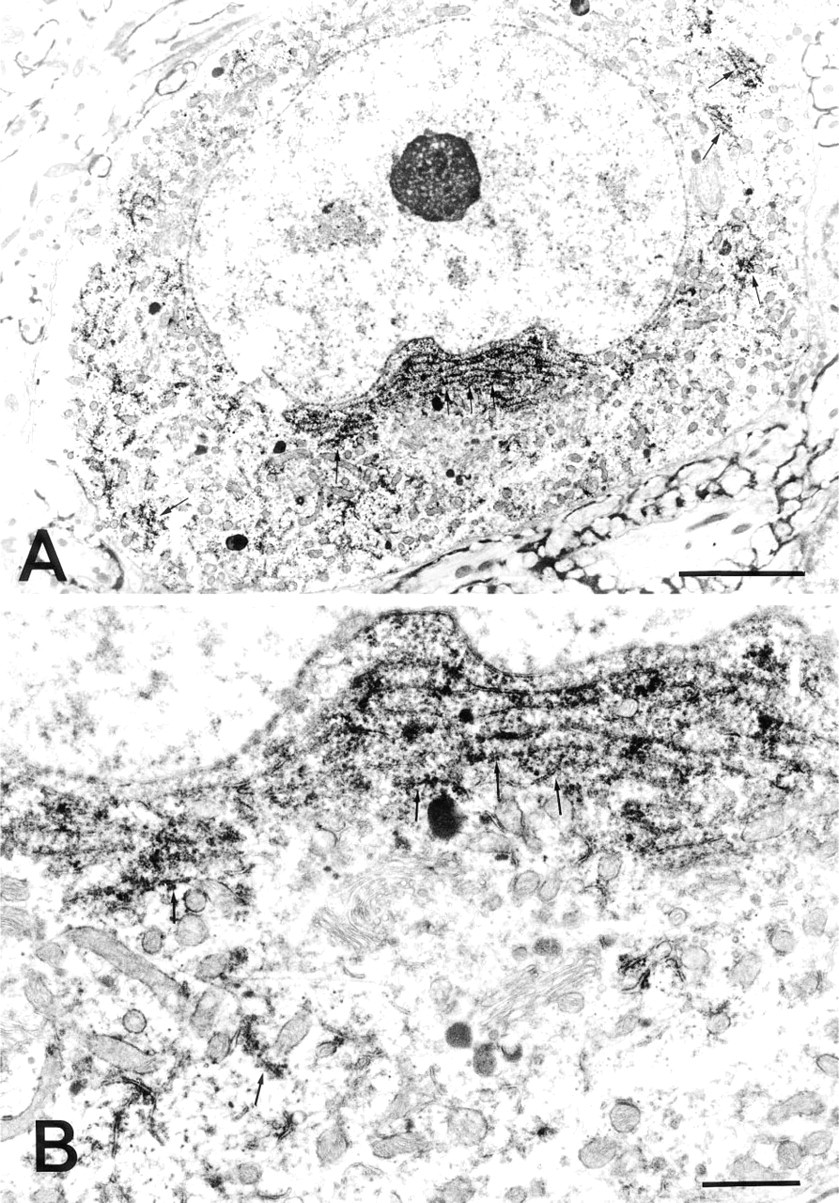
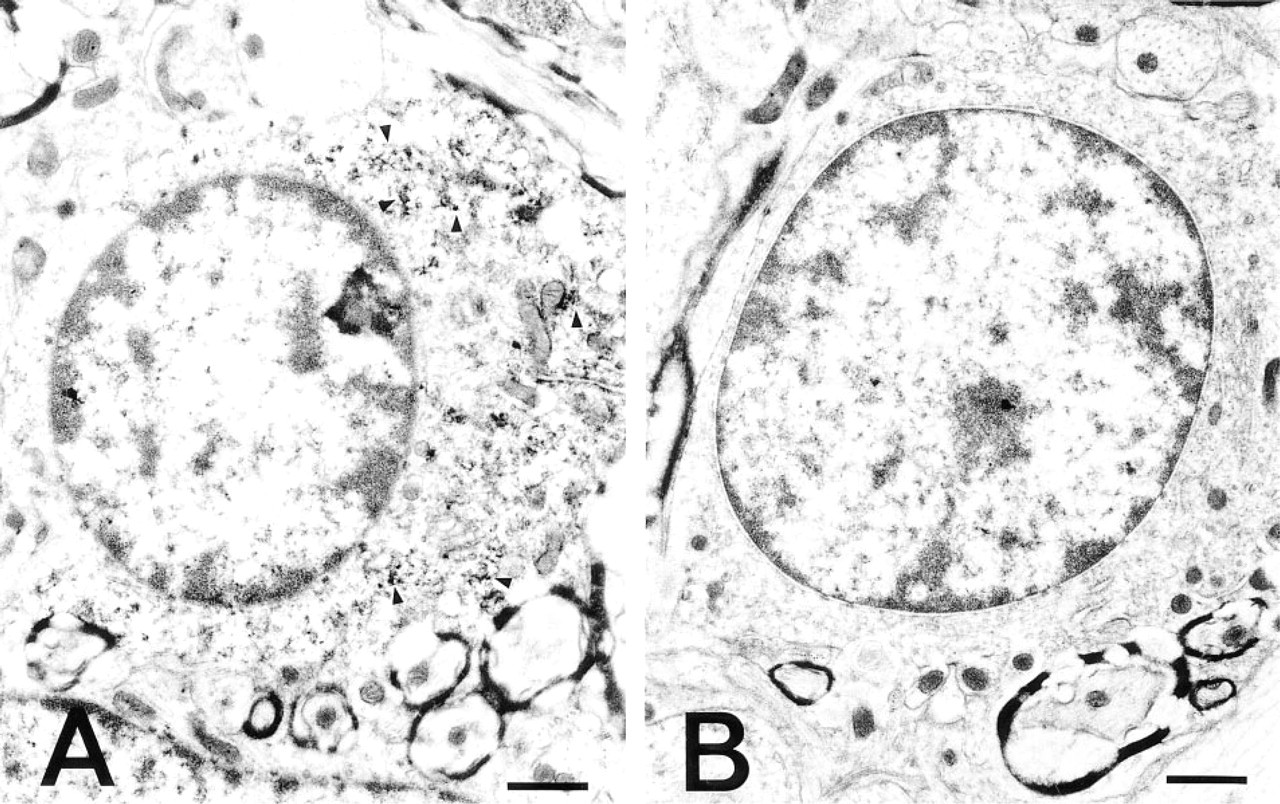
Immunocytochemical localization of PAs in a nerve cell in the lateral reticular nucleus of the medulla oblongata. (
Electron Microscopy
The color-developed specimens prepared as above were then fixed with 1.0% osmium tetraoxide in 50 mM cacodylate buffer, pH 7.4, for 1 hr at RT and dehydrated in a series of graded ethanol solutions. After immersion in propylene oxide (Nacalai Tesque; Kyoto, Japan) (three times for 10 min each), the samples were immersed in a mixture (1:1) of propylene oxide and Epon 812 resin (Taab Laboratories; Reading, Berks, UK) overnight and embedded in Epon 812 resin in a routine way. The regions to be studied were cut with a 2-mm diameter punch, mounted on Epon blocks, and sectioned on a horizontal plane into ultrathin sections, which were then immediately observed in a 100 CX Jeol electron microscope.
For quantification, images of electron microscopic photographs (both sample and control specimens) taken under constant conditions were digitized into a computer through a video camera head (ITC-370M; Olympus-Ikegami, Tokyo, Japan) according to the method of Shin et al. (1995). The optical density of the regions of the neural rough endoplasmic reticulum, nucleus, and nucleolus in the photographs was measured by a computer-assisted image analyzer (Olympus-SP500; Tokyo, Japan) under constant conditions.
Control Experiments
In the PA immunocytochemistry study of the rat brain, the specificity of immunostaining was ascertained by incubating control sections with (a) the secondary antiserum alone, (b) type-matched MAb (IgG1) anti-penicillin (30–80 ng/ml; Cosmo Biological, Tokyo, Japan), and (c) ASPM-29 MAb preabsorbed with Spm at a concentration of 50 μg/ml.
Results
The polyamine immunoelectron microscopic study was developed with the pre-embedding method using 50-μm Microslicer sections of the lower parts of the medulla oblongata. In the present PA immunocytochemistry using 2.5% GA as a fixative no proteolytic pretreatment of the sections was needed before the PA immunoreaction, in contrast to our previous studies using tissues other than central neural tissues (Fujiwara 1994; Fujiwara and Masuyama 1995; Fujiwara et al. 1996). This may indicate that the central neural tissues were loosely fixed with GA and became easily accessible for PA immunoreaction.
By light microscopy PA immunoreactivity was seen in the cytoplasm and dendrites of all the nerve cells in the lower part of the medulla oblongata at all levels, whereas the nuclei and axons were devoid of staining. Strong PA immunostaining was noticed in the large nerve cells of the hypoglossal nucleus, nucleus ambiguus, accessorial nucleus, and dorsal motor nucleus of the vagus, and in the reticular nuclei including the gigantocellular reticular nucleus, lateral paragigantocellular nucleus, lateral reticular nucleus, and medullary reticular nucleus (Figure 1A). The patterns of PA immunostaining were either clustered masses or blocklike (Figure 1B). Immunoreactivity was seen to a lesser degree in the small neurons of the inferior olive nuclei, in the nucleus of the solitary tract, in the gracile nucleus, and in the spinal trigeminal nucleus (Figure 1A). Very weak staining or no staining at all was observed in the glial cells and nerve fibers passing through the white matter and entering into the gray matter. No immunoreaction occurred with the ASPM-29 MAb preabsorbed with Spm (Figure 1C), with anti-penicillin MAb, or with the secondary antiserum alone.
Immunoelectron microscopic examination of the neurons in the lateral reticular nucleus showed that PA was unevenly distributed in the cytoplasm and dendrites of the neurons. Variations in the amount of deposit were found from section to section and even within the same ultrathin section. The PA immunoreactivity was specifically and intensely located in the ribosomes attached to the membranes (rER), in the free ribosomes (polysomes) clustered between cisternae, and in the polysomes widely spread throughout the entire cytoplasm (Figures 2A and 2B). In addition, cell organelles at the terminal and in the middle of short dendrites of the neurons were all strongly labeled (Figures 3B and 3C). The nucleus, nucleolus, Golgi cisternae, Golgi vesicles, lysosomes, and mitochondria in the cytoplasm of the perikaryon were devoid of PA staining (Figures 2A and 2B). Among the glial cell types, the oligodendrocytes were immunoreactive with the ASPM-29 MAb, staining the ribosomes (polysomes) in the cytoplasm (Fig. 5A). The axons, both myelinated and unmyelinated, and the synaptic vesicles in the nerve endings around the neurons were completely PA-negative, suggesting that PAs are not axonally transported. The sections stained with the preabsorbed ASPM-29 MAb or the anti-penicillin MAb did not show any PA immunoreaction (Figures 3A and 5B). Electron microscopic photographs of sample and control specimens of subcellular organelles in a neuron (Figures 2A and 3A) were quantitatively analyzed by a computer and revealed that the rER was significantly labeled, in contrast to the nucleolus and nucleus (Figure 4).
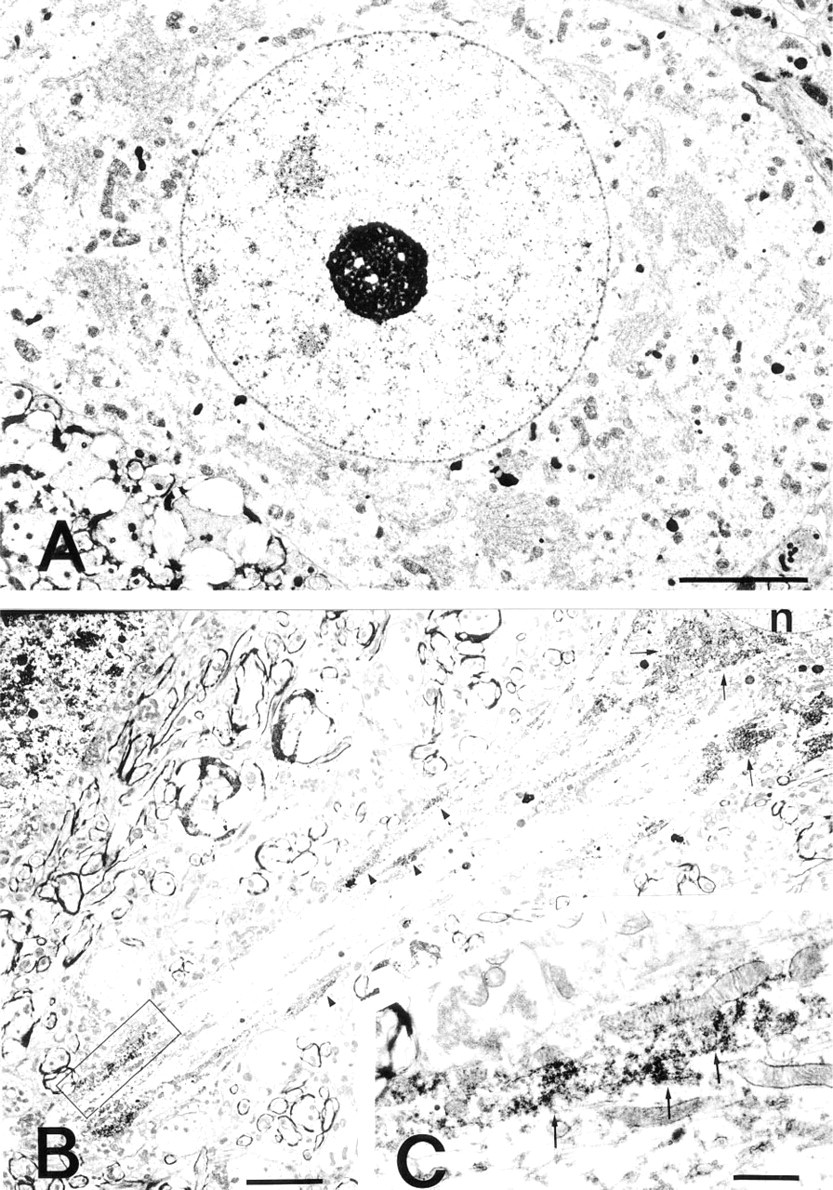
Control for PA immunocytochemical staining and immunocytochemical localization of PAs in the dendrite of a nerve cell. (
Discussion
Since the discovery of Spm phosphate complexes by Leeuwenhoek in 1678, the subcellular localization of PAs has not been previously elucidated.
The present immunoelectron microscopic findings that PAs are predominantly located on free (polysomes) and attached ribosomes of the rough endoplasmic reticulum (Nissl bodies) in the cytoplasm, but not in the nuclei, axons, and nerve endings of neurons of the central nervous system, suggests that PAs are closely involved in the translation processes of protein biosynthesis. PAs may be located close to the sites rich in proteins within the ribosomes because, in our previous study of polyamine immunocytochemistry, protease treatment was indispensable before the first immunoreaction (Fujiwara 1994; Fujiwara and Masuyama 1995; Fujiwara et al. 1996). This is probably due to PA masking resulting from GA bridge formation of the ribosomal proteins, judging from the results of the present immunoelectron microscopic study and also previous PA biochemistry studies (Stevens and Pascoe 1972).
The present light microscopic findings in the medulla oblongata, i.e., that the large neurons with efferent fibers and in the reticular nuclei possessed high amounts of PAs but the small neurons possessed low amounts of PAs (Figure 1A), appear to be in agreement with the commonly accepted idea that Nissl bodies (rER) are larger and more abundant in large neurons than in small ones. The patterns of PA immunostaining were either clustered masses or block-like, just like those of the Nissl bodies (Figure 1B).
Many reports, using extracts derived from a number of cell types, indicate that, in the cell-free systems PAs stimulate a variety of processes of protein biosynthesis, such as stabilization and activation of tRNA (Cohen 1978), stimulation of initiation of translation (Konecki et al. 1975), ensuring fidelity of translation (Abraham 1983), stimulation of the synthesis of specific proteins (Woese and Fox 1977; Lake et al. 1984; Higgins 1995), and stimulation of in vitro reconstitution of ribosomal subunits (Kakegawa et al. 1986a, b). In fact, ribosomes and tRNA isolated from a variety of eukaryotic and prokaryotic cells have been found to contain significant amounts of PAs (Stevens and Pascoe 1972; Cohen 1978). It may be difficult to determine exactly the connection between the PA association and these processes in cells. However, PAs may be partly responsible for the maintenance of their secondary structure, or they may play an active part in the formation of the complexes between ribosomes, mRNA, and the various factors necessary for protein biosynthesis (Kyner et al. 1973; Goyns 1982).
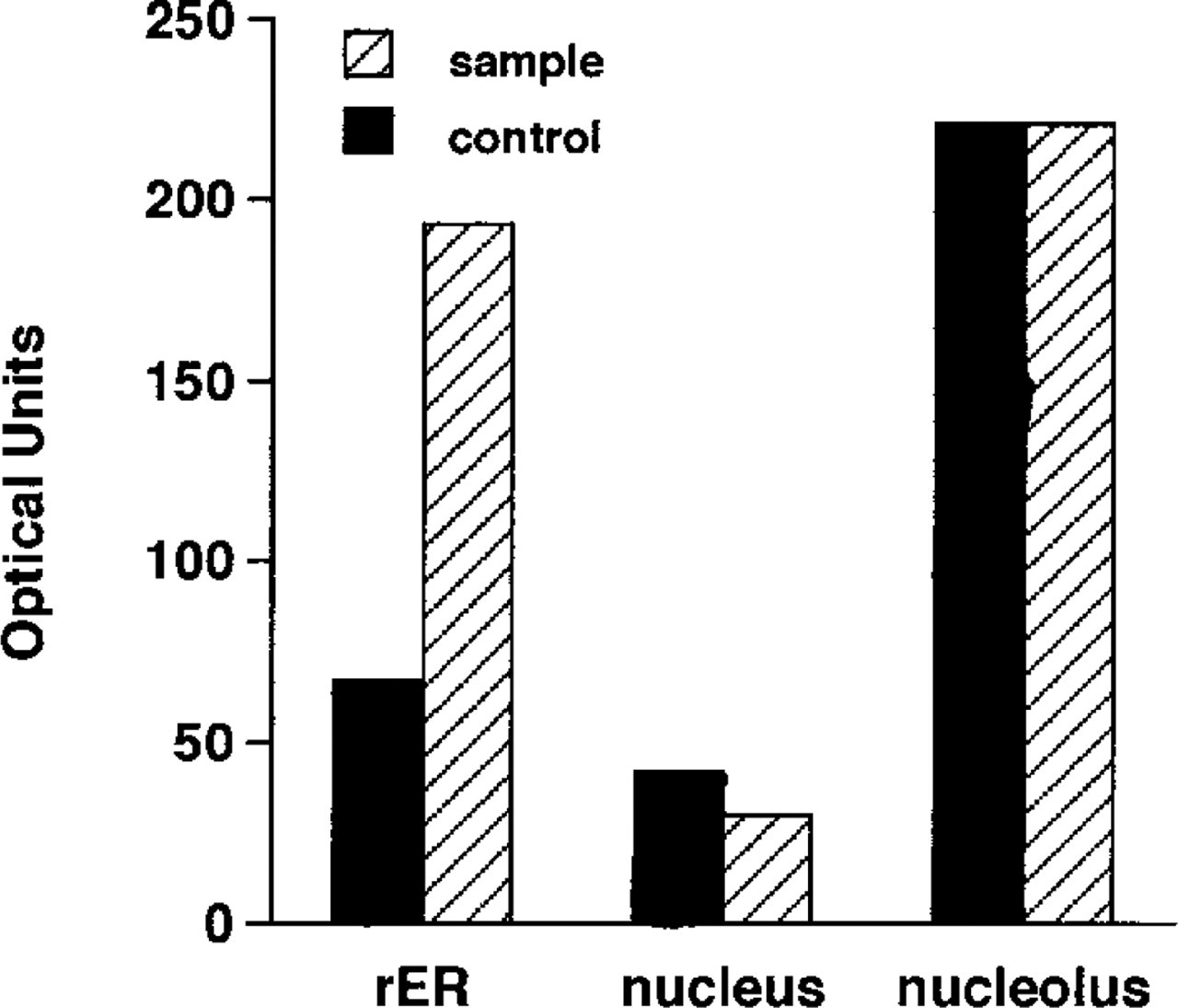
Relative staining intensity of the immunostaining and control specimens. The regions of rER were analyzed in comparison with those of the nucleus and nucleolus, because the size of the attached ribosomes was so small that a precise comparison between sample and control specimens (Figures 2A and 3A) could not be made.
Because the present immunoelectron microscopic findings point out a coincidence between ribosomal subunit association in the maturation processes of the ribosomes and the presence of PAs, further polyamine immunocytochemistry and biochemistry are needed to determine whether or not PAs play a role in maturation processes (Jordan and Cullis 1982; Hadjiolov 1985; Sommerville 1986). There may be PA regulation systems to keep the PA concentrations very low in the nucleus. In this way the immature ribosomes may be protected from maturation in the nucleus without binding to the incomplete hnRNA. Biochemical studies by Comolli and Riboni (1980) and by Miret and Goldemberg (1989) have reported that PAs are implicated in ribosomal subunit association. Some in vitro and in vivo experiments have indicated an interaction between Spm and DNA, suggesting that Spm is associated with the nucleus (Shimizu et al. 1965; Flink and Pettijohn 1975; Gosule and Schellman 1976). In the present study, however, no nuclear PA immunoreactivity was evident. This discrepancy may be due to PA redistribution, which probably occurs during the DNA/PA fractionation procedures, or it is likely that some of the suggested actions only reflect the fact that PAs can interact with many acidic macromolecules in vitro and may therefore not be physiologically relevant. There may be additional factors that went undetected in our study because they have a nucleic PA concentration below the detection limit of the present immunoelectron microscopic technique.
Immunocytochemical localization of PAs in the oligodendrocyte. (
We had previously identified the light microscopic localization of PAs and had found that the PAs are abundant in the cytoplasm of neoplastic cells, active protein- or peptide-secreting cells, including exocrine and endocrine cell types, central and peripheral nerve cells, and rapidly proliferative cells in the gastrointestinal epithelia (Fujiwara 1994; Fujiwara and Masuyama 1995; Fujiwara et al. 1996,1997). These results were confirmed by the present study's immunoelectron microscopic findings. All of these cells are characterized by many free or attached ribosomes within the cells. In addition, no PA immunoreactivity was observed in the nuclei of any of the types of cells tested in such polyamine immunocytochemistry studies (Fujiwara 1994; Fujiwara and Masuyama 1995; Fujiwara et al. 1996,1997).
Because many recent reports have also shown that PAs have effects on membrane proteins such as ion channels and receptors (Schuber 1989; Ficker et al. 1994; Rock and Macdonald 1995; Williams 1997), PAs may not be restricted to the ribosomes in cells, as stated in the present study. We are now undertaking a further immunoelectron microscopic investigation into PA localization in neurons employing experimental models of cerebral ischemia in rats. This may be valuable for the elucidation of the biological roles of PAs other than a close association with protein biosynthesis in cells.
The present study suggests that PAs are one of the components of biologically active ribosomes in neurons, thus closely associating with protein biosynthesis. This may indicate an important role of PAs in cell proliferation and growth (Janne et al. 1978; Goyns 1982; Tabor and Tabor 1984; Pegg 1986), in addition to their biological effects on membrane structure and function (for reviews see Schuber 1989; Rock and Macdonald 1995; Williams 1997).
Footnotes
Acknowledgments
We are very grateful to Dr M. Shin for the computerassisted image analysis, Mr T. Suematsu for technical assistance with electron microscopy, and Dr Y. Inoue for valuable discussions throughout this study.