
Abstract
We observed the ultrastructural distribution of MAGP-36 by immunoelectron microscopy in human and bovine tissues. MAGP-36 was present in microfibrils associated with tropoelastin in skin, aorta, and spleen. It was not detected in microfibrils from the ocular zonule and kidney mesangium that were not associated with tropoelastin. In skin, MAGP-36 was present in both early immature elastic fibers and mature elastic fibers. In mature elastic fibers, MAGP-36 was localized around amorphous elastic cores at the elastin-microfibril interface and in electron-dense bundles. Localization of MAGP-36 in elastic fibers coincided with the distribution of lysyl oxidase, an enzyme that plays a pivotal role in the deposition of tropoelastin. These findings suggest that MAGP-36 may be involved in elastogenesis. (J Histochem Cytochem 47:1049–1056, 1999)
M
In addition to its major component, fibrillin, many microfibril-associated proteins have been identified. These include MAGP-1 and 2 (Henderson et al. 1996; Finnis and Gibson 1997), emilin (Bressan et al. 1993), fibulin-2 (Reinhardt et al. 1996), amyloid P (Li et al. 1991), and lysyl oxidase (Kagan et al. 1986). MAGP-36 was initially discovered in porcine aorta and is believed to be one of these microfibril-associated proteins (Kobayashi et al. 1989, 1994). However, the precise distribution of this protein and its functions are still obscure.
MAGP-36 has recently received much attention because its human homologue, AAAP-40 (MAGP-3), is immunoreactive with IgG purified from the serum and aortic wall of patients with abdominal aortic aneurysms (AAAs) (Tilson 1995; Gregory et al. 1996; Xia et al. 1996; Hirose et al. 1997). Autoimmunity against AAAP-40 might play a role in the pathogenesis of AAAs. AAAP-40 also shares sequences with the envelope glycoprotein of the Ebola virus (Tilson et al. 1996). Another human homologue of MAGP-36, MFAP-4, is commonly deleted in Smith-Magenis syndrome (SMS) (Zhao et al. 1995). In addition, human MAGP-36 is the target protein for Tranilast, a therapeutic drug for hypertrophic scars and keloids (Hiromi Furuichi, personal communication). Hypertrophic scars are known to cause excessive collagen accumulation and deposition of fine immature elastic fibers in the dermis (Russell et al. 1995; Roten et al. 1996). These findings suggest that MAGP-36 may play an important role in the organization of connective tissue. To address this question, we describe the ultrastructural localization of MAGP-36 in skin, aorta, spleen, and kidney. Our results indicate that the distribution of MAGP-36 is matched exactly by the distribution of tropoelastin.
Materials and Methods
Antibodies
An antiserum was prepared from rabbits immunized with purified porcine MAGP-36 as previously described (Kobayashi et al. 1989). MAbs to tropoelastin and fibrillin-1 were supplied by Elastic Products Company (Owensville, MO), and NeoMarkers (Fremont CA), respectively. The rabbit antiserum to bovine fibrillin was supplied by LSL (Tokyo, Japan). Goat anti-rabbit IgG antibodies and goat anti-mouse IgG antibodies conjugated with colloidal gold particles were supplied by British Biocell (Cardiff, UK).
Sample Preparation
Specimens of hypertrophic scar tissue from a 19-year-old woman were obtained during surgery. Samples of normal human skin were taken from the marginal part of the surgical specimen after informed consent was obtained from the patient. Specimens of other tissues from adult cows were obtained from a local slaughterhouse. Tissues for postembedding immunoelectron microscopy were fixed in 4% formaldehyde-0.2% glutaraldehyde in 0.1 M phosphate buffer, pH 7.4, for 5 hr at 4C, washed extensively with 0.1 M phosphate buffer, dehydrated through a graded series of ethanol-water mixtures at −20C, and embedded in LR White. The resin was polymerized at 40C for 48 hr under vacuum. For pre-embedding immunolabeling, tissue samples were fixed in 4% formaldehyde in 0.1 M phosphate buffer, pH 7.4, for 5 hr at 4C, rinsed twice with 0.1 M phosphate buffer, cryoprotected in 20% sucrose at 4C, and frozen with OCT compound in dry ice-ethanol. Some samples were also fixed in 4% formaldehyde-2.5% glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4, for 2 hr at 4C and postfixed in cacodylate-buffered 1% OsO4 for 1 hr at 4C. The samples were washed in buffer, 50% ethanol and then stained en bloc with 2% uranyl acetate in 50% methanol for 1 hr at 4C. After dehydration through a graded series of ethanol-water mixture and propylene oxide, these samples were embedded in Epon 812.
Immunolabeling
Postembedding immunoelectron microscopy was conducted according to Atkinson et al. (1989). Ultrathin sections were collected on uncoated nickel grids, blocked with PBS-milk-Tween (PMT) for 30 min, and incubated with appropriate dilutions (1:1000 for MAGP-36, 1:100 for tropoelastin, and 1:200 for fibrillin) of antibodies in PMT overnight at 4C. The sections were then washed several times in PBS-BSA-Tween (PBT), incubated for 1 hr with a diluted solution (1:20) of goat anti-rabbit IgG antibodies or goat anti-mouse IgG antibodies conjugated with colloidal gold particles, rinsed in PBT followed by distilled water, and stained for EM. For double labeling experiments, a mixture of rabbit serum and mouse MAbs was used as primary antibody. Secondary antibodies, 20-nm gold-conjugated goat anti-rabbit IgG antibody, and 10-nm gold-conjugated goat IgG anti-mouse antibody, were applied separately for 1 hr each.
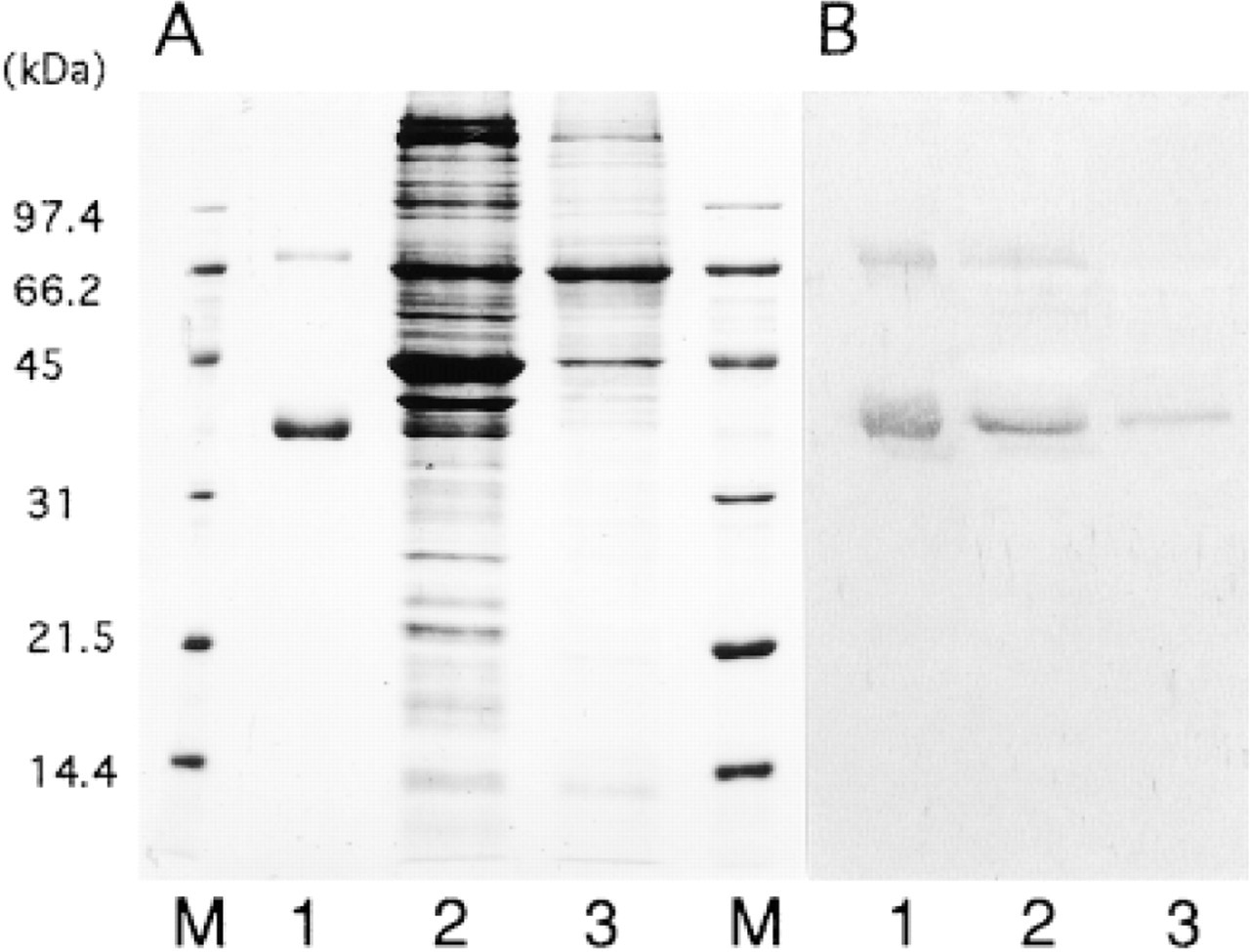
Immunoblotting of bovine aorta and human skin with anti-MAGP-36 antibody. (A) SDS-PAGE using a 12% acrylamide gel stained with Coomassie Blue. (B) Immunoblot stained with anti-MAGP-36 antibody. Lane 1, purified MAGP-36; Lane 2, crude extract from bovine aorta; Lane 3, crude extract from human skin; Lane M, markers.
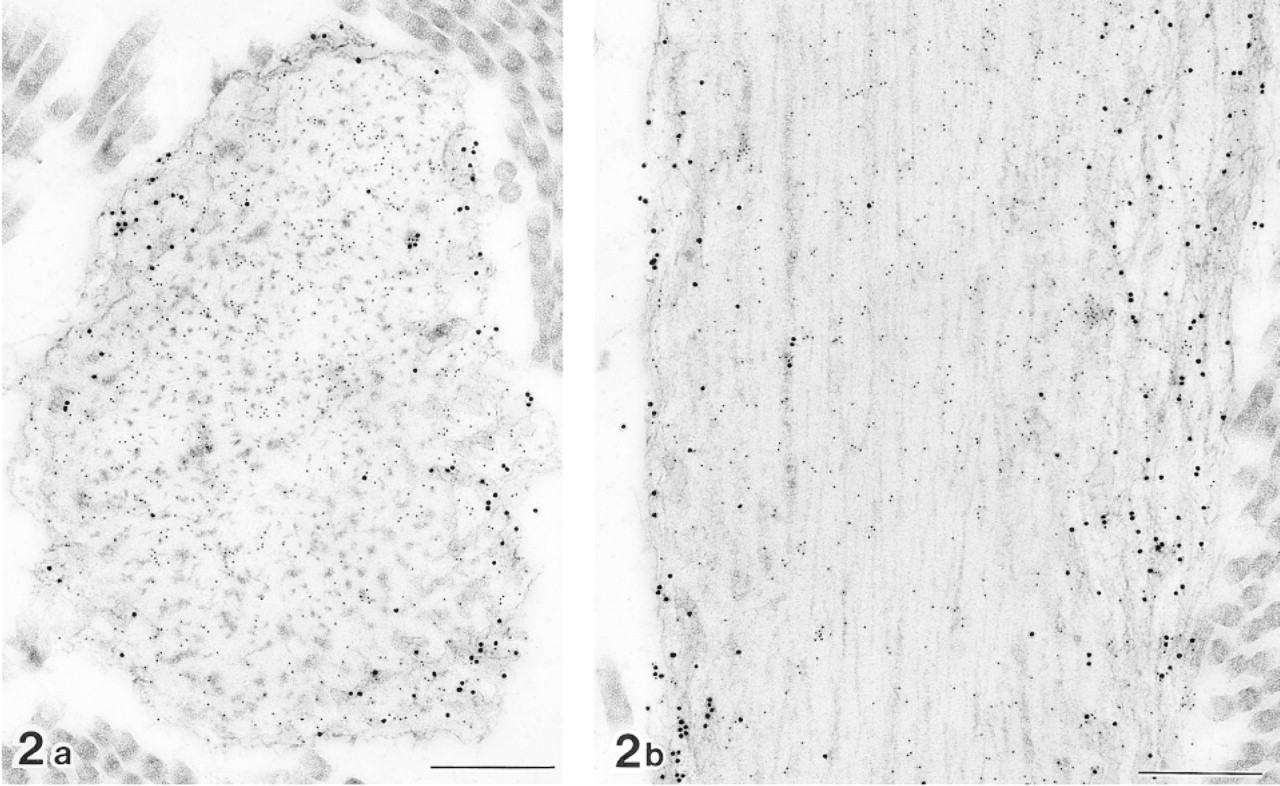
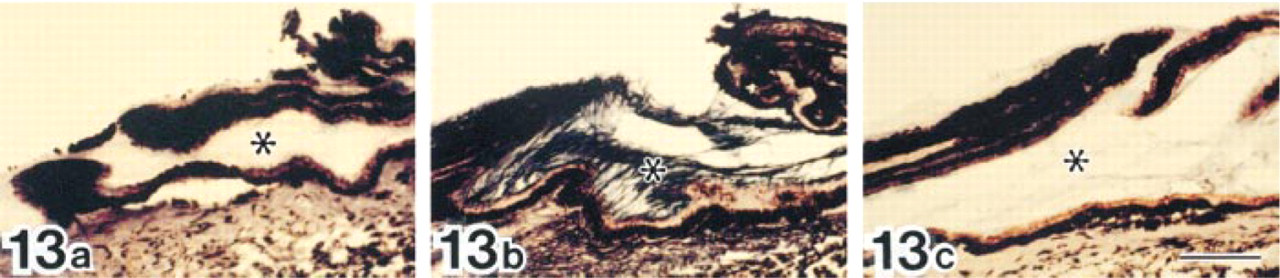
Cross (a) and longitudinal (b) sections of elastic fibers in normal human skin after double labeling with anti-tropoelastin antibody (10-nm gold particles) and anti-MAGP-36 antibody (20-nm gold particles). MAGP-36 antibody labeled with 20-nm gold particles is present at the surface of amorphous elastic core and electron-dense bundles at the outer periphery of elastic core. Bars = 0.5 μm.
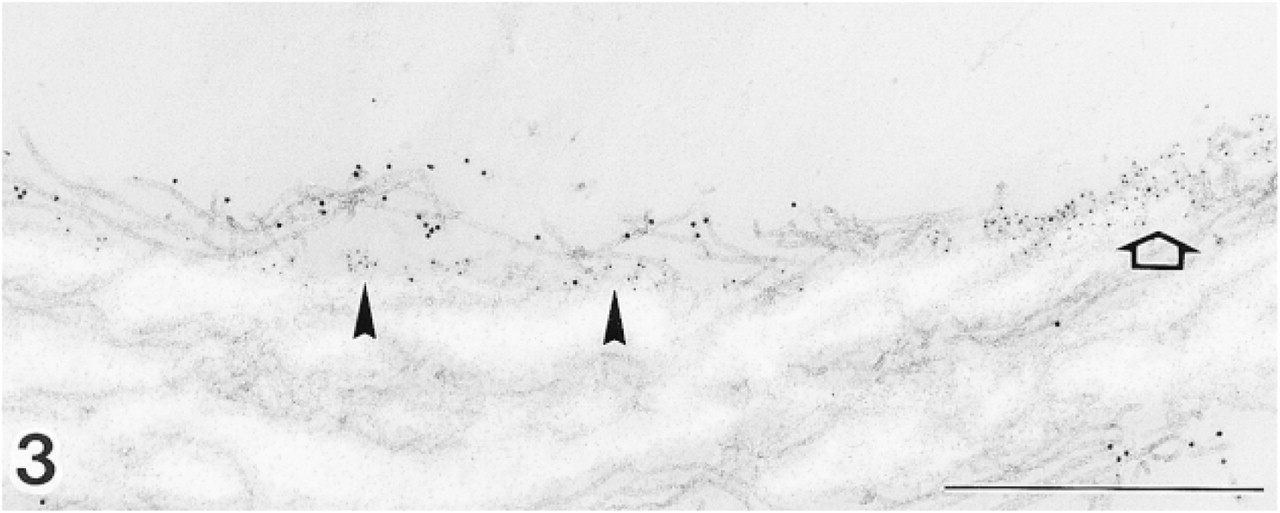
Pre-embedding labeling with anti-MAGP-36 antibody (5-nm gold particles) and anti-fibrillin-1 antibody (10-nm gold particles) on elastic fiber of normal human skin. Labeling for MAGP-36 is preferentially located at the surface of elastic cores and the interface of elastin and microfibrils (arrowheads). Intense labeling covers regions of the surface of elastic core in which microfibrils had apparently been stripped off (arrow). The microfibrils away from the elastin bundle do not bind the antibody for MAGP-36. All microfibrils are immunoreactive with anti-fibrillin-1 antibody. Bar = 0.5 μm.

Longitudinal section of elastic lamina of bovine aorta after labeling with anti-MAGP-36 antibody. Note the thin (#13 nm) and smooth microfibrils in the elastic core (arrowheads), and thick electron-dense bundles at periphery of the elastic core (arrows). Labeling is preferentially associated with the electron-dense bundles. Bar = 0.5 μm.
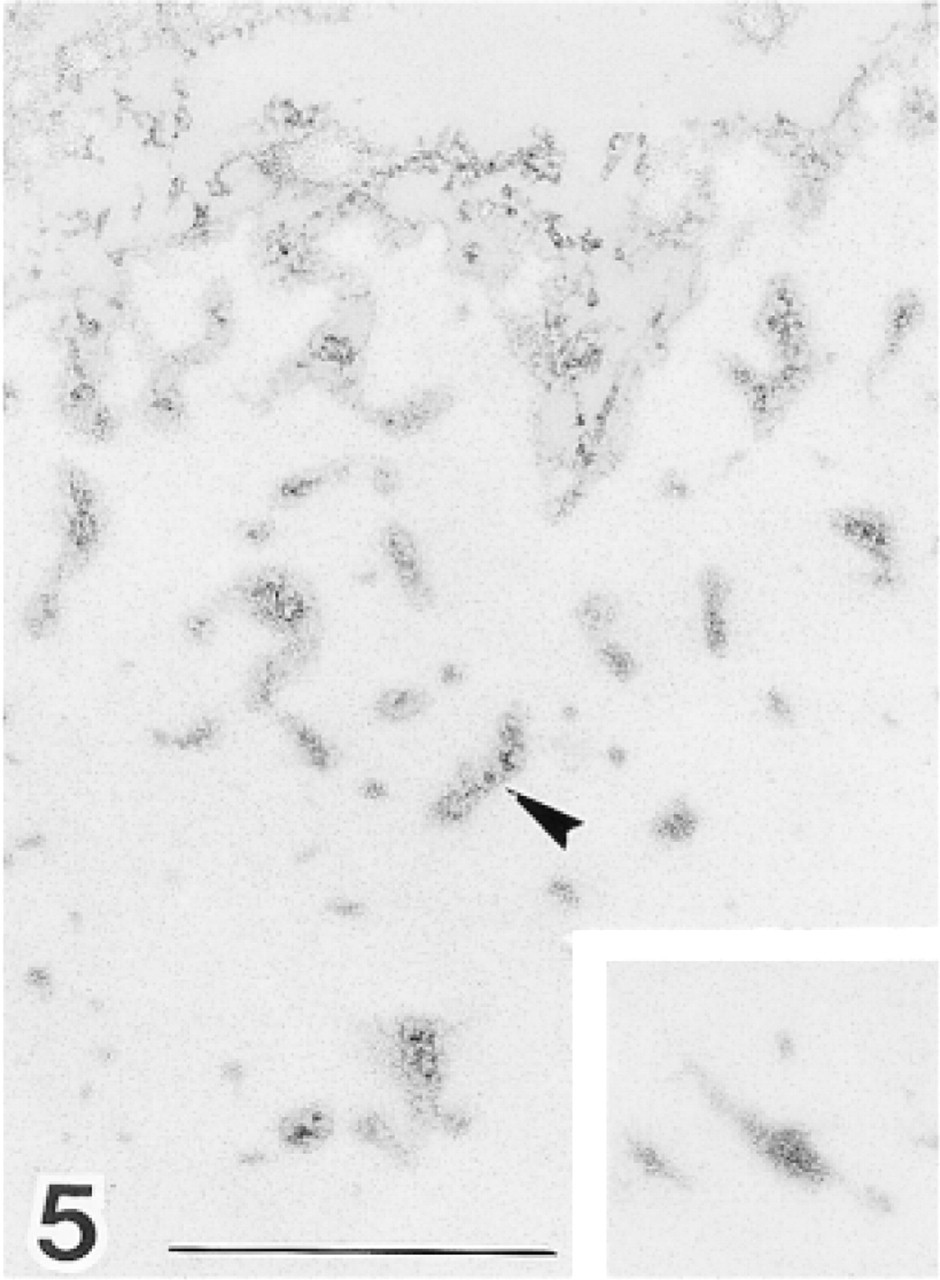
Periphery of elastic lamina in bovine aorta in Epon section. Tubular microfibrils surround the elastic core. Tubular microfibrils and amorphous electron-dense materials are also recognizable in electron-dense bundles distributed at medium density. Electron-dense bundles are packed too densely at the center of the elastic core for the internal structure to be resolved (inset). Bar = 0.5 μm.

Elastic lamina in bovine aorta after labeling with anti-fibrillin antibody. Fibrillin antibody labeled with 20-nm gold particles is present at the electron-dense bundles. Bar = 0.5 μm.
Pre-embedding double labeling experiments were performed on cryosections (15 μm thick) that were mounted on slides. After washing several times in PBS, the sections were blocked with PMT for 30 min and incubated with a mixture of anti-MAGP-36 rabbit serum and anti-fibrillin-1 MAb overnight at 4C. The antibodies were diluted 1:100 for anti-MAGP-36 rabbit serum and to 40 μg/ml for anti-fibrillin-1 MAb in PMT. The sections were then washed several times in PBS and incubated overnight at 4C with a mixture of goat anti-rabbit IgG antibodies conjugated with 5-nm gold particles and goat anti-mouse IgG antibodies conjugated with 10-nm gold particles. The secondary antibodies were diluted 1:20 in PMT. After rinsing several times in PBS, sections were fixed in 2.5% glutaraldehyde in PBS for 30 min, dehydrated, and embedded in Epon 812.
All ultrathin sections for electron microscopy were stained for 15–30 min with 2% alcoholic uranyl acetate and 5 min with Reynold's lead citrate before examination in a JEOL 1200EX electron microscope.
Immunohistochemistry was performed on cryosections of formaldehyde-fixed tissue using the avidin-biotin complex (ABC) method. Cryosections (15 μm thick) were saturated with PMT for 30 min and incubated with primary antibodies at 4C overnight. These antibodies were detected with an ABC kit (Vector Laboratories; Burlingame, CA). Alkaline phosphatase activity of tissue-bound ABC complex was visualized with 0.3 mg/ml nitroblue tetrazolium (NBT) and 0.1 mg/ml 5-bromo-4-chloro-3-indolyl phosphate (BCIP) diluted in 0.1 M Tris-HCl buffer (pH 9.5) containing 0.1 M NaCl and 5 mM MgCl2.
Method specificity was confirmed by one or more of the following controls: (a) the sections were processed without primary antibody; (b) the primary antibody was replaced by non-immune serum; or (c) the primary antibody was replaced by antiserum adsorbed with purified MAGP-36 which was excised from the gel. For the double labeling, crossreactions between inappropriate antibodies were also checked by applying each primary antibody separately and linking them with the inappropriate secondary antibody.
Results
Specificity of Antibodies
Antibody specificity was carefully assayed against crude tissue preparations. Anti-MAGP-36 antibody did not react with other protein species because it recognized only two bands corresponding to MAGP-36 and its dimeric form (68 kD) by Western blotting of crude extracts from human skin and bovine aorta (Figure 1) (Kobayashi et al. 1994).
Ultrastructural Localization of MAGP-36 in Normal Skin and Aorta
Strong labeling was observed on elastic fibers in dermis and all layers of aorta, intima, media, and adventitia by immunohistochemistry and postembedding immunoelectron microscopy (Figures 2, 4, 11, and 12). In elastic fibers, labeling for MAGP-36 was associated with their surface and with electron-dense bundles at the outer periphery of their amorphous elastic cores. Although some microfibrils were observed in the interior of the elastic cores, labeling of these microfibrils was sparse.
Electron-dense bundles varied in density in Epon sections (Figure 5). Bundles increased from moderate to high density between the outer periphery of the elastic cores and their centers. Tubular microfibrils and electron-dense amorphous material were present in bundles at the periphery of the elastic core. Specific structures could not be resolved at the center of these cores, where density of bundles was highest. Electron-dense amorphous materials were also observed at the surface of elastic cores, although these were not as clear as in bundles that occurred at moderate density around the core. Fibrillin was also detected by immunoelectron microscopy in the electron-dense bundles (Figure 6).
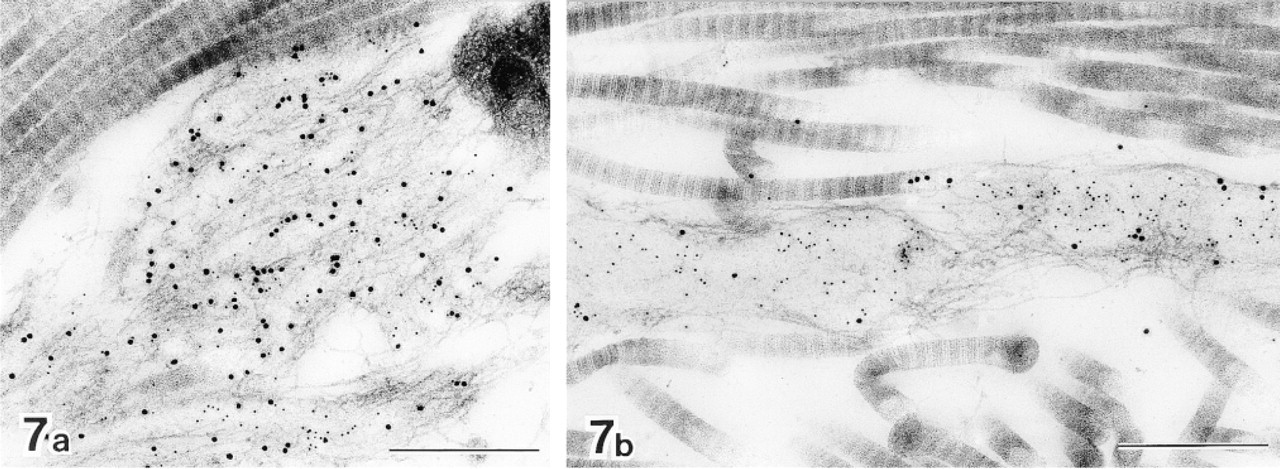
Immature elastic fibers in human hypertrophic scar tissue after double labeling with anti-tropoelastin antibody (10-nm gold particles) and anti-MAGP-36 antibody (20-nm gold particles). In early immature elastic fibers, labeling for tropoelastin is intermingled with the labeling for MAGP-36 (a). In later stages some amorphous elastic deposits are seen. Most labeling for MAGP-36 was located at the intersection of microfibrils with amorphous elastin deposits (b). Bars = 0.5 μm.
With the pre-embedding procedure, labeling for MAGP-36 was detected on the surface of elastic cores and at the elastin-microfibril interface (Figure 3). Intense labeling covered regions of the surface of elastic core in which microfibrils had apparently been stripped off. The microfibrils away from the elastic core did not bind the antibody for MAGP-36.
Ultrastructural Localization of MAGP-36 in Hypertrophic Scars
MAGP-36 was localized in most of the immature elastic fibers observed in hypertrophic scar tissue. Some of these immature elastic fibers did not contain any amorphous elastic deposits, but tropoelastin was detected by immunolabeling in all fibers in which MAGP-36 was present. The amount of tropoelastin varied, depending on the development of the fibers. In early immature elastic fibers, labeling for tropoelastin was intermingled with labeling for MAGP-36. Some labeling for tropoelastin was closely associated with the labeling for MAGP-36 at this stage (Figure 7a). At later stages of development, some amorphous elastin deposits were seen. The majority of gold particles specific for MAGP-36 were located where microfibrils intersected with these elastin deposits (Figure 7b).
Trabeculae in bovine spleen after double labeling with anti-tropoelastin antibody (10-nm gold particles) and anti-MAGP-36 antibody (20-nm gold particles). Typical elastic fibers are seen in the trabeculae. Labeling with antibodies was identical to that observed in normal skin. Bar = 0.5 μm.

Microfibrils in bovine ocular zonule after double labeling with anti-tropoelastin antibody (10-nm gold particles) and anti-MAGP-36 antibody (20-nm gold particles). There is no immunoreactivity in this region. Bar = 0.5 μm.
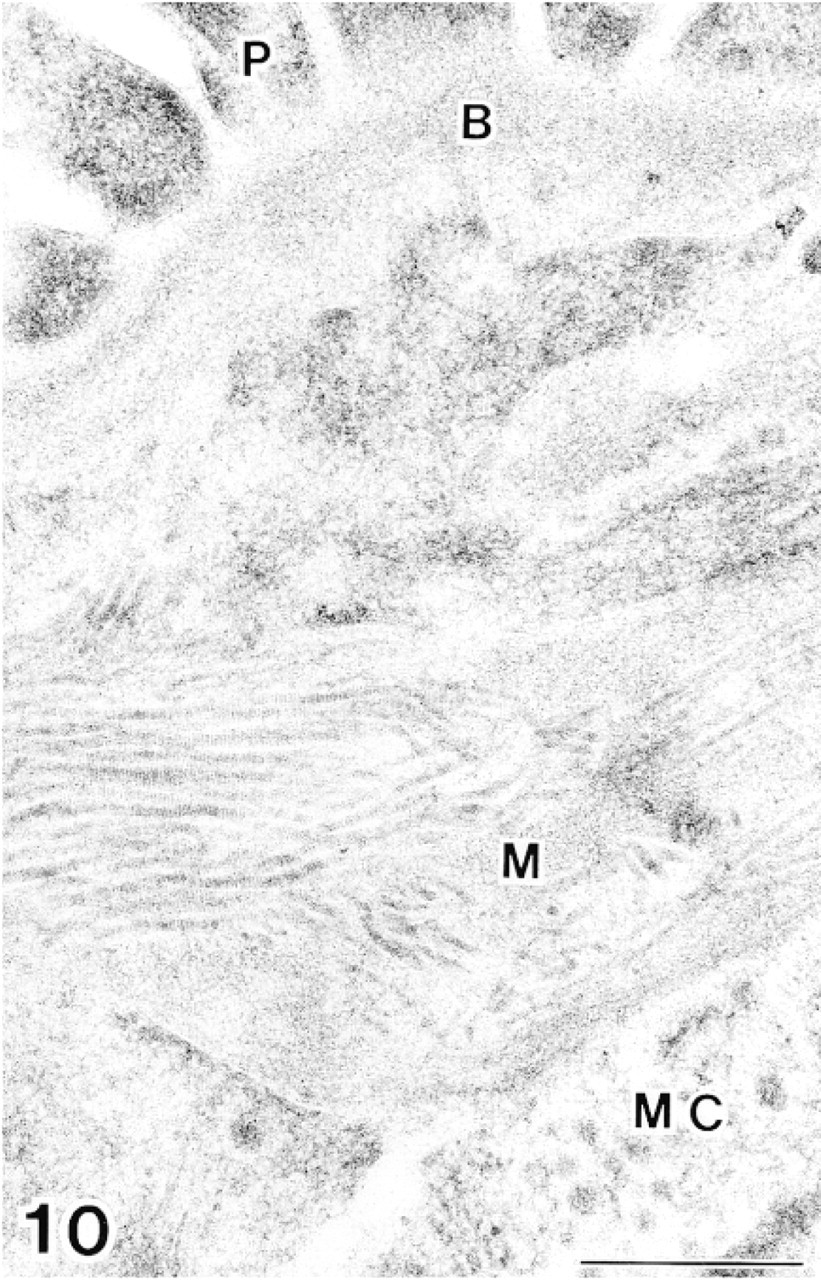
The glomerulus in bovine kidney after double labeling with anti-tropoelastin antibody (10-nm gold particles) and anti-MAGP-36 antibody (20-nm gold particles). There is no immunoreactivity in the mesangium. P, podocyte; B, basal lamina; M, mesangium; MC, mesangial cell. Bar = 0.5 μm.
Ultrastructural Localization of MAGP-36 in Other Tissues
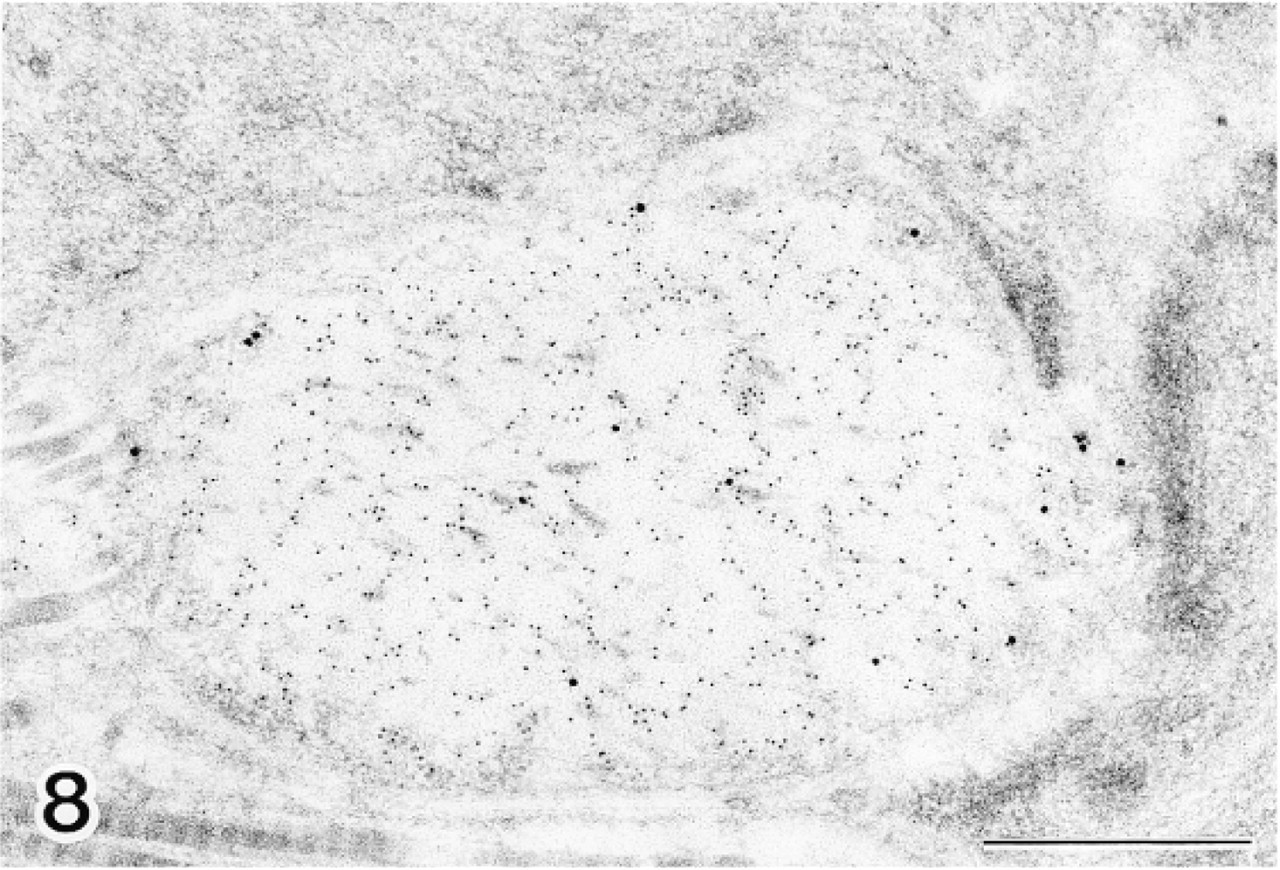
In spleen tissue, labeling of typical elastic fibers with anti-MAGP-36 and anti-tropoelastin antibodies was identical to that observed in normal skin (Figure 8). Labeling for MAGP-36 was localized where micro-fibrils intersect with the amorphous core of these fibers. Labeling for tropoelastin, by contrast, was localized within the amorphous core. In the red pulp region of spleen, labeling was limited for both MAGP-36 and tropoelastin. In kidney, the mesangial region of the glomerulus is known to contain microfibrils that are not associated with elastin. We did not observe labeling with either anti-MAGP-36 or anti-tropoelastin antibody in the mesangial region of the glomerulus (Figure 10). Labeling was also sparse in other regions of kidney for both MAGP-36 and tropoelastin. In the eye, the ocular zonule contained bundles of micro-fibrils in parallel alignment. These microfibrils did not react with either anti-MAGP-36 or anti-tropoelastin antibody (Figures 9 and 13).
Gold deposits were not seen in control specimens prepared using normal rabbit serum or the serum preadsorbed with purified MAGP-36 instead of anti-MAGP-36 antibody. Completely negative results were also obtained when anti-tropoelastin antibody or anti-fibrillin-1 antibody, but not gold-labeled second antibody, was omitted from the immunolocalization protocol described in Materials and Methods. Crossreaction was not detected in control specimens that were stained with combinations of inappropriate antibodies.
Discussion
The results reported here describe the ultrastructural distribution of MAGP-36 in different tissues and its localization during elastogenesis. Among several microfibril-associated proteins, such as fibrillin, emilin, MAGP-1, MAGP-2, fibulin-2, and amyloid P, MAGP-36 is unique in that its distribution is exactly matched by the distribution of tropoelastin (Li et al. 1991; Bressan et al. 1993; Zhang et al. 1994; Gibson et al. 1996; Henderson et al. 1996; Reinhardt et al. 1996; Finnis and Gibson et al. 1997). This distribution suggests that MAGP-36 may be involved in elastogenesis. In skin, MAGP-36 was present in both early immature and mature elastic fibers. In mature elastic fibers, MAGP-36 was localized at the elastin-microfibril interface surrounding the amorphous elastic core and dense bundles of these fibers, but not on the micro-fibrils away from the elastin bundle. This ultrastructural localization coincides with the localization of lysyl oxidase (Kagan et al. 1986), which plays a pivotal role in elastogenesis by crosslinking of newly accreted tropoelastin at the elastin-microfibril interface. MAGP-36 may serve to anchor or align this enzyme with respect to the growing bundle of tropoelastin.
Human dermis incubated with anti-MAGP-36 antibody (a) or normal rabbit serum (b). A positive reaction is found along the outer surface of elastic fibers. Bar = 200 μm.
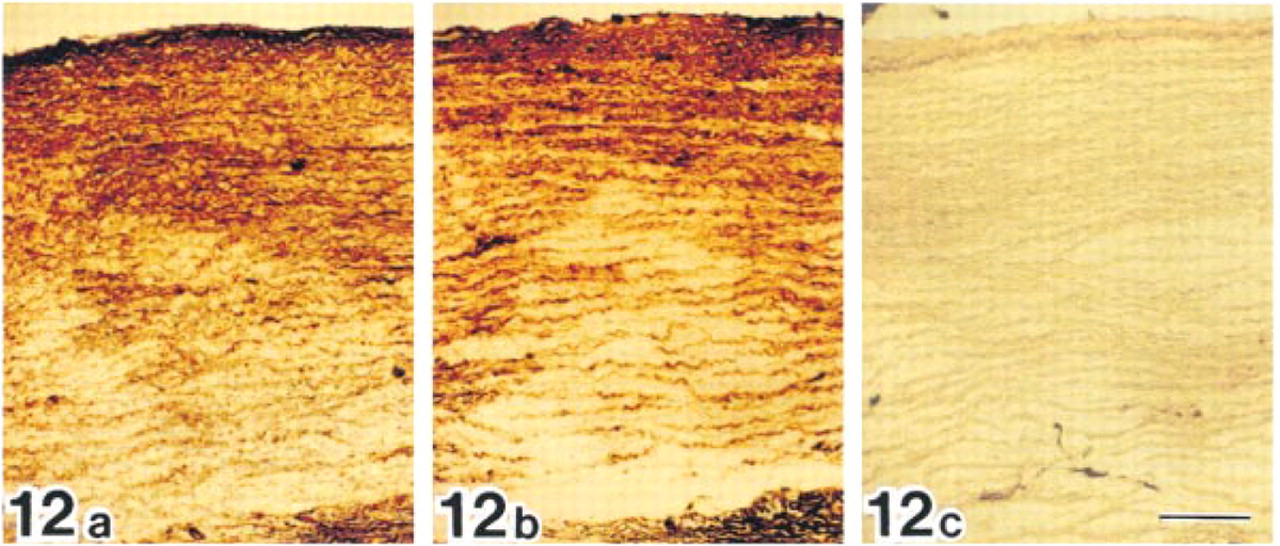
Bovine aorta incubated with anti-MAGP-36 antibody (a), anti-fibrillin-1 antibody (b), or normal rabbit serum (c). Positive reactions are seen along elastic fibers in intima and media after incubation with anti-MAGP-36 antibody or anti-fibrillin-1 antibody. Bar = 200 μm.
Bovine ciliary body incubated with anti-MAGP-36 antibody (a), anti-fibrillin-1 antibody (b), or normal rabbit serum (c). A positive reaction is found in the ocular zonule (∗) after incubation with anti-fibrillin-1 antibody but not with anti-MAGP-36 antibody or normal rabbit serum. Bar = 200 μm.
There are some previous descriptions of electron-dense bundles in elastic fibers (Ross et al. 1969; Varadi 1972; Hashimoto et al. 1975). Some workers interpret the electron-dense bundles as dark-staining microfibrils (Ross et al. 1969; Varadi 1972). Others interpret them as electron-dense granular and amorphous materials rather than microfibrils (Hashimoto et al. 1975). In Epon sections, variations were observed in the density of electron-dense bundles between the periphery of elastic cores and the central portion of the cores. Hashimoto et al. (1975) have suggested that this variation in density may reflect the degree of condensation of electron-dense materials. In bundles spaced at moderate densities, we could recognize some tubular microfibrils and dense amorphous materials.
Immunoelectron microscopy with anti-fibrillin antibody also suggested the presence of microfibrils in the electron-dense bundles. It appears certain that these peripheral dense bundles contain microfibrils and electron-dense amorphous materials. Bressan et al. (1993) have reported similar electron-dense material on the surface of elastic cores where microfibrils and amorphous elastin are in close contact. In our study, labeling for MAGP-36 was preferentially associated with electron-dense bundles and the perimeter of elastic cores where microfibrils were associated with electron-dense materials. We did not observe labeling on microfibrils away from the elastic core or in the central part of elastic cores where microfibrils were not associated with electron-dense materials. These facts suggest that MAGP-36 might be a component of this electron-dense material.
Two human homologues of MAGP-36, MFAP-4 and AAAP-40 (MAGP-3) have been reported (Zhao et al. 1995; Tilson et al. 1996; Xia et al. 1996; Hirose et al. 1997). These proteins share a high level of sequence homology with a fragment of bovine MAGP-36 and show similar molecular weight to bovine MAGP-36 (Kobayashi et al. 1994). However, these proteins are reported to have a more ubiquitous tissue distribution than MAGP-36 by Western blot techniques (Kobayashi et al. 1989). The results from this study demonstrate that MAGP-36 is more widely distributed in tissues and in species than has previously been recognized. MAGP-36 was detected in bovine aorta and spleen and in human skin by immunoelectron microscopy. These facts suggest that MAGP-36 might be identical to MFAP-4 or AAAP-40.
In this study, our findings suggest that MAGP-36 may be involved in elastogenesis. By affecting elastogenesis, MAGP-36 might be related to the pathogenesis of several diseases, such as AAAs, SMS, and hypertrophic scar formation. Further studies are needed to determine the function of MAGP-36 and its relationship to the pathogenesis of these diseases.