
Abstract
The pacemaker of the heart, the sinoatrial (SA) node, is characterized by unique electrical coupling properties. To investigate the contribution of gap junction organization and composition to these properties, the spatial pattern of expression of three gap junctional proteins, connexin45 (Cx45), connexin40 (Cx40), and connexin43 (Cx43), was investigated by immunocytochemistry combined with confocal microscopy. The SA nodal regions of rabbits were dissected and rapidly frozen. Serial cryosections were double labeled for Cx45 and Cx43 and for Cx40 and Cx43, using pairs of antibody probes raised in different species. Dual-channel scanning confocal microscopy was applied to allow simultaneous visualization of the different connexins. Cx45 and Cx40, but not Cx43, were expressed in the central SA node. The major part of the SA nodal-crista terminalis border revealed a sharply demarcated boundary between Cx43-expressing myocytes of the crista terminalis and Cx45/Cx40-expressing myocytes of the node. On the endocardial side, however, a transitional zone between the crista terminalis and the periphery of the node was detected in which Cx43 and Cx45 expression merged. These distinct patterns of connexin compartmentation and merger identified suggest a morphological basis for minimization of contact between the tissues, thereby restricting the hyperpolarizing influence of the atrial muscle on the SA node while maintaining a communication route for directed exit of the impulse into the crista terminalis.
T
The morphological structures responsible for intercellular coupling in the heart are gap junctions, clusters of transmembrane channels that span the plasma membranes of neighboring myocytes (Saffitz et al. 1995; Gros and Jongsma 1996; Severs et al. 1996). Gap junctions between SA nodal cells are small and sparse compared with those in other heart tissues, consistent with relatively poor coupling (Masson-Pevet et al. 1979; De Mazière et al. 1992). Apart from the size and number of gap junctions, however, current evidence suggests that the type of gap junction protein (connexin) expressed is a key determinant of conduction properties in different tissues of the heart (Saffitz et al. 1994, 1995; Gros and Jongsma 1996; Severs et al. 1996). Gap junction channels made from different connexin types in vitro are reported to show distinct unitary conductance, ionic selectivity, and molecular permeability properties (White et al. 1995; Bruzzone et al. 1996; White and Bruzzone 1996). Most studies report that the major gap junction protein of the heart, connexin43 (Cx43), is undetectable in the central SA node (ten Velde et al. 1992; Oosthoek et al. 1993; Davis et al. 1994, 1995), although several reports claim the presence of this connexin in at least some nodal cells (Anumonwo et al. 1992; Trabka-Janik et al. 1994; Kwong et al. 1998). Few studies have investigated the possible presence of other connexin types. Although connexin40 (Cx40) and connexin45 (Cx45) have been reported in the SA node of canine and human heart (Davis et al. 1994, 1995; Kwong et al. 1998), recent results with an alternative antibody to Cx45 indicate a need to reevaluate earlier findings on this connexin (Coppen et al. 1998).
To advance our understanding of how the expression of different connexin types may contribute to the specialized properties of the SA node, this study set out to identify which connexin or connexins are expressed in SA nodal myocytes in the rabbit and to determine how the connexin expression patterns of the SA node and surrounding atrial myocytes are spatially connected. We were particularly interested in establishing whether, by simultaneous visualization of different connexin types combined with the high resolution afforded by confocal microscopy, evidence might be found for the existence of gradients of coupling at the SA node-crista terminalis border. A prerequisite for this study was a set of reliable connexin-specific antibody probes, raised in different species to permit specific secondary detection in double label experiments and suitable for application in the rabbit (Coppen et al. 1998; Yeh et al. 1998). With the aid of this approach, we now demonstrate that Cx45 is a major connexin of the rabbit SA node and that distinct boundaries and a region of merger exists between the Cx45-expressing myocytes of the node and the Cx43-expressing myocytes of the surrounding atrium. These findings provide new insights into the structural basis for the electrophysiological properties of the SA node.
Materials and Methods
Animals and Tissue Preparation
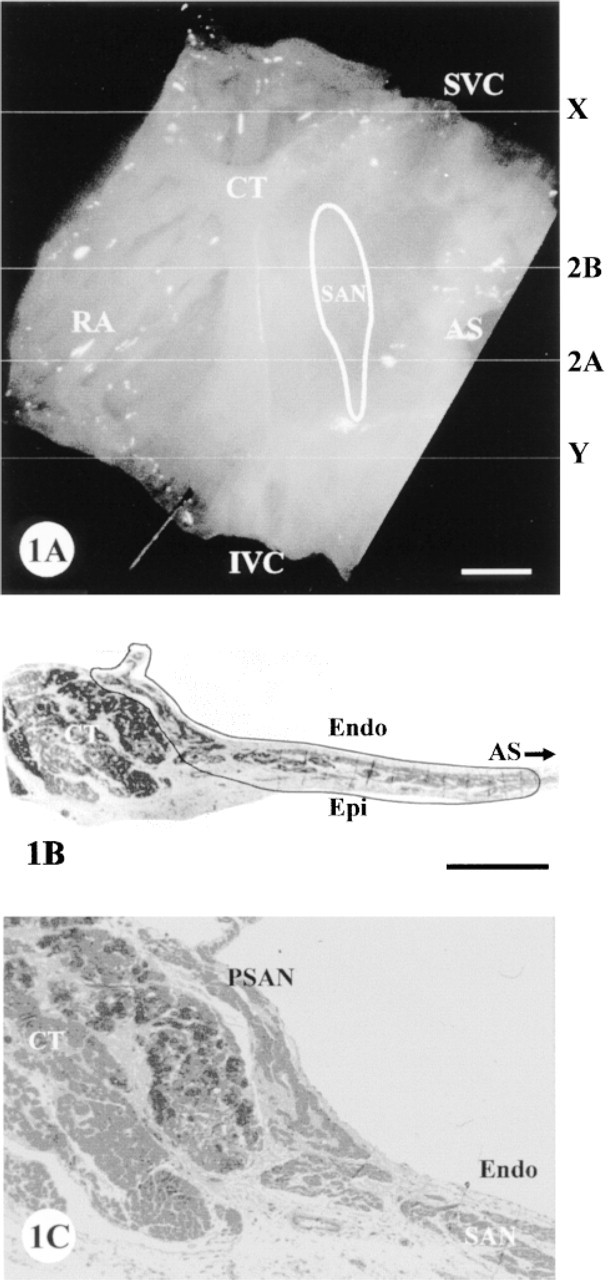
Rabbits weighing 1.5-2.0 kg were anesthetized with IV sodium pentobarbital (30–40 mg/kg). The chest was opened and the heart was rapidly excised and placed in KrebsRinger solution at 34C. To dissect SA nodal preparations for cryosectioning and connexin localization, the following procedure was used. The right atrium was separated from the remainder of the heart and opened by a longitudinal incision in the free wall to expose the endocardial surface. With the preparation pinned to a silicon base, the atrium was trimmed to leave a preparation approximately 15 × 15 mm, which included the SA node and some of the surrounding atrial muscle. The SA node is located in the intercaval region between the superior and inferior venae cavae (Figure 1). Laterally, it is bounded by the atrial septum on one side and the crista terminalis and the atrial appendage on the other. Towards the superior vena cava, a thick muscle bundle (the central pectinate muscle) projects from the crista terminalis towards the atrial appendage. The leading pacemaker site in the SA node is typically located ∼1 mm caudal to the lower edge of this muscle bundle and 0.5-1.0 mm from the crista terminalis. All animal procedures were conducted in accordance with the United Kingdom Animals (Scientific Procedures) Act, 1986.
Dissected SA node preparations, still pinned to their base, were immersed in melted gelatin. The preparation was raised to allow penetration of the gelatin beneath the tissue. After brief cooling at 4C to set the gelatin, the silicon rubber and pins were removed and the preparation frozen at −50C in isopentane for subsequent serial cryosectioning.
Supporting histological observations were made on Evans blue-stained cryosections, and on semithin sections of epoxy resin-embedded tissue after Toluidine Blue staining. For the latter, the tissue was fixed in phosphate-buffered 2% paraformaldehyde/2.5% glutaraldehyde for 1 hr, rinsed in phosphate buffer, postfixed in 1% osmium tetroxide (phosphate-buffered) for 1 hr, dehydrated, and embedded in epoxy resin.
For Western blot analysis, the SA node, crista terminalis, and atrial septum were carefully dissected under a binocular microscope and rapidly frozen in liquid nitrogen. Samples of left ventricle and right ventricle were frozen in parallel as controls.
Antibodies
Polyclonal antibodies against Cx45 and Cx40 were raised in guinea pig. The anti-Cx45 antiserum Q14E (GP42) and the anti-Cx40 antiserum V15K (GP318) were prepared, purified and characterized as described previously (Coppen et al. 1998; Yeh et al. 1998). For immunofluorescence work, the anti-Cx45 antibody (hereafter referred to as GP42) was used at 1 in 100 dilution, and the anti-Cx40 antibody (GP318) at 1:250 dilution. Detection of Cx43 was performed using a commercially available monoclonal antibody (raised against residues 252–270 of rat Cx43 (Chemicon; Harrow, UK) at 1:1000 dilution for both Western blot analysis and immunofluorescence.
(
For Western blotting, alkaline phosphatase-conjugated secondary antibody to mouse IgGs (used at 1:5000 dilution) was purchased from Pierce & Warriner (Chester, UK). All the fluorophore-conjugated (CY3 and CY5) secondary antibodies for immunoconfocal localization were purchased from Chemicon (used at 1:500 dilution). The fluorophore-conjugated antibodies were adsorbed against IgGs from a range of species before use to ensure specificity and avoid crossreactivity in double labeling experiments.
Sample Preparation and Western Blotting
For Western blotting, whole tissue homogenates were prepared by pulverizing the frozen specimens under liquid nitrogen using a mortar and pestle. After lysing the powdered frozen tissues in SB20 (20% SDS, 0.15 mol/liter Tris, pH 6.8), an aliquot was removed for protein estimation (BioRad DC protein assay) and 2-mercaptoethanol added to the remainder to a final concentration of 2.5%.
Ten μg of total protein per lane was run on 12.5% SDS-PAGE and electrophoretically transferred overnight to PVDF membrane at constant voltage. Transfer was assessed by Ponceau S staining and the membrane blocked with TBS/0.1% Tween 20 with 5% dried milk powder. The membrane was incubated with primary antibody, diluted in TBS/0.1% Tween/1% BSA, for 1 hr at room temperature (RT), washed, and incubated with alkaline phosphatase-conjugated secondary antibody, diluted in TBS/0.1% Tween/10% normal goat serum, for 1 hr. The membrane was washed and the enzyme activity revealed using a freshly prepared NBT/BCIP substrate solution (Pierce and Warriner).
Immunofluorescent Labeling
Frozen sections (10 μm thick) were cut from four frozen SA nodal preparations. The sections were mounted on poly-L-lysine-coated glass slides, which were then stored at −65C until use. The sections were fixed by immersing the slides in methanol at −20C for 5 min and were then washed three times with PBS. Blocking was carried out for 1 hr with 1% bovine serum albumin (BSA) in PBS before incubation with primary antibody (diluted in 1% BSA in PBS) for 2 hr (GP42 and GP318) or 1 hr (monoclonal anti-Cx43). After washing five times with PBS over 30 min, the sections were incubated with CY3-conjugated secondary antibodies (diluted in 1% BSA in PBS) for 1 hr. The slides were washed a further five times with PBS over 30 min, mounted with Citifluor mounting medium (Agar; Essex, UK), and the coverslips sealed with clear nail polish.
For double labeling of Cx45 and Cx43 and of Cx40 and Cx43, the sections were incubated sequentially with the primary antibodies, the monoclonal anti-Cx43 being applied second. The secondary antibodies (CY3-conjugated anti-guinea pig IgG and FITC-conjugated anti-mouse IgG) were applied together in the presence of 10% normal rabbit serum. Double labeling for Cx40 and Cx45 was not feasible because both antibodies were raised in the same species.
For peptide inhibition experiments, the antibodies were incubated with 50 μg/ml peptide at RT for 30 min before application to the tissue sections or Western blots. Controls for the immunofluorescent labeling experiments were (a) omission of the primary antibody (one or both in the case of double labeling), (b) using the inappropriate secondary antibody for each individual primary antibody, and (c) using a single primary followed by both secondary antibodies.
Immunolabeled sections were examined by confocal laser scanning microscopy using a Leica TCS 4D instrument equipped with an argon/krypton laser and fitted with the appropriate filter blocks for detection of CY3 and FITC fluorescence. The images recorded were projections of five consecutive single optical sections taken at 0.5-μm intervals.
Results
Isolated SA node preparations from four rabbits were serially cryosectioned in a plane perpendicular to the crista terminalis, as illustrated in Figure 1A. The serial sections were taken through the zone marked X to Y (Figure 1A. Toluidine Blue staining of selected semithin sections of epoxy resin-embedded tissue helped to determine the histological orientation and the extent of the SA node tissue (Figures 1B and 1C). The SA node tissue appears lighter in color than the atrial muscle and occupies the thin intercaval region with an extension onto the endocardial surface of the crista terminalis (the periphery of the SA node; Figure 1C).

Cx43 distribution in survey views assembled from montages of confocal micrographs of representative sections taken from the inferior side of the SA node (
On imaging Cx43 labeling at low magnification by confocal microscopy, the SA nodal region was readily identifiable as a conspicuous immunonegative zone between the strongly Cx43-positive crista terminalis and the atrial myocardium on the septal side (Figures 2A and 2B). The boundary between Cx43-positive and Cx43-negative zones was sharply delineated, and although some bundles of Cx43-expressing atrial myocytes penetrated the nodal region, appearing as “islands” in sectional view (Figure 2A, no extensive interdigitation between the zones was apparent. The lack of Cx43 in the SA node compared with the crista terminalis, atrial septum, and ventricular myocardium was confirmed by Western blotting (Figure 3).
The typical extent of the Cx43-negative region, as reconstructed from serial sections, is indicated in Figure 1A. Survey montage views such as those in Figure 2 were used for mapping the features of connexin expression observed at high magnification throughout this region. The montages illustrated in Figures 2A and 2B come from sections across the inferior edge and center of the SA node, respectively. Because multiple montages of this type were used for mapping connexin distribution in detail, an idealized diagram summarizing the montage features (Figure 4) is used here to illustrate the locations of all the high-magnification panels presented in Figures 5–8, which have been drawn from different representative sections through the nodal region.
Dual-channel imaging of double labeled preparations was used to permit high-resolution simultaneous visualization of Cx45 with Cx43 and of Cx40 with Cx43. With this approach, close inspection throughout the Cx43-negative SA nodal region revealed extensive, widely distributed Cx45 labeling in the form of small but sharply defined spots (Figure 5A, and a similar pattern of labeling was observed for Cx40 (Figure 5B. The relative amounts of Cx45 and Cx40 labeling in the SA node varied from one heart to the next, but the overall level of labeling for these connexins was always substantially lower than that observed for Cx43 in the surrounding atrial myocardium (Figure 5A, inset). The labeling for Cx45 and Cx40 differed from that of Cx43 by being widely dispersed rather than aggregated into intercalated disk clusters and by comprising individual spots of much smaller size.

(
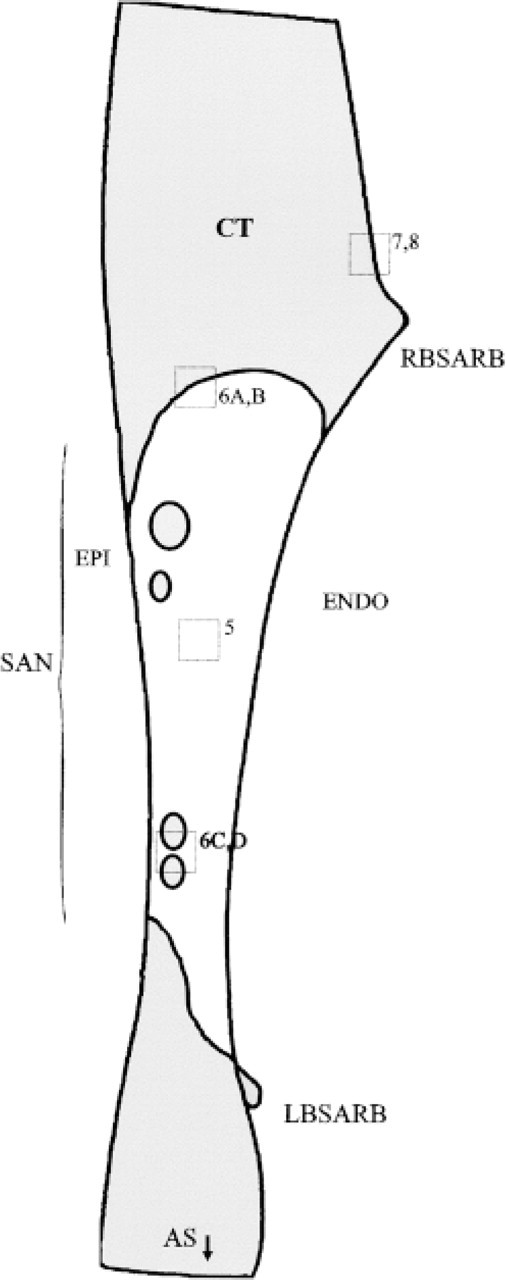
Diagrammatic representation of a typical section through the SA node preparation, based on the combined observations of multiple images from four hearts. The framed areas represent the positions from which high-magnification views are presented in subsequent figures. The central SA node (SAN) is indicated by bracketed zone. CT, crista terminalis; RBSARB, right branch of the SA ring bundle; LBSARB, left branch of the SA ring bundle; AS, atrial septum; Epi, epicardial side; Endo, endocardial side.
The double labeling approach disclosed two distinctive expression patterns at the boundary of Cx43-expressing regions of the crista terminalis. The first, involving a sharp demarcation of Cx43 from both Cx45 and Cx40, characterized the epicardial side of the crista terminalis extending inwards. Here, Cx45 and Cx43 labeling were each confined to discrete, nonoverlapping zones (Figure 6A. A corresponding distinct boundary was present between Cx40- and Cx43-labeled zones (Figure 6B. An intervening, discontinuous connective tissue layer occupied stretches of the boundary (Figures 6A and 6B; see also Figure 1C), although strands of Cx45/Cx40-expressing nodal cells were in places closely approximated to groups of Cx43-expressing cells of the crista terminalis. A similar sharp delineation between Cx43 and Cx45/Cx40 was apparent at the border of the node with “islands” of penetrating myocyte bundles (Figures 6C and 6D). The second distinctive pattern, in contrast to these clearly delineated zones, involved a zone in which Cx43 and Cx45 expression merged and overlapped (two examples are shown in Figures 7 and 8). This co-expressing zone was exclusively on the endocardial surface of the crista terminalis, extending beyond the right branch of the SA ring bundle (Figures 7A and 8A), and was continuous with the Cx45-positive nodal tissue on one side and the Cx43-expressing atrial myocytes of the crista terminalis on the other. Deeper zones of the crista terminalis showed only Cx43 (Figures 7B and 8B). Cx40 was not detectable in the co-expressing zone (Figures 7C and 8C). This general pattern of expression was apparent in all the SA node preparations studied, although some variation in the width of the Cx45-positive region penetrating the crista terminalis was encountered from one heart to the next, as illustrated in the two examples in Figures 7 and 8. Cx45 and Cx43 co-expression was also apparent in the right and left branches of the SA ring bundle.
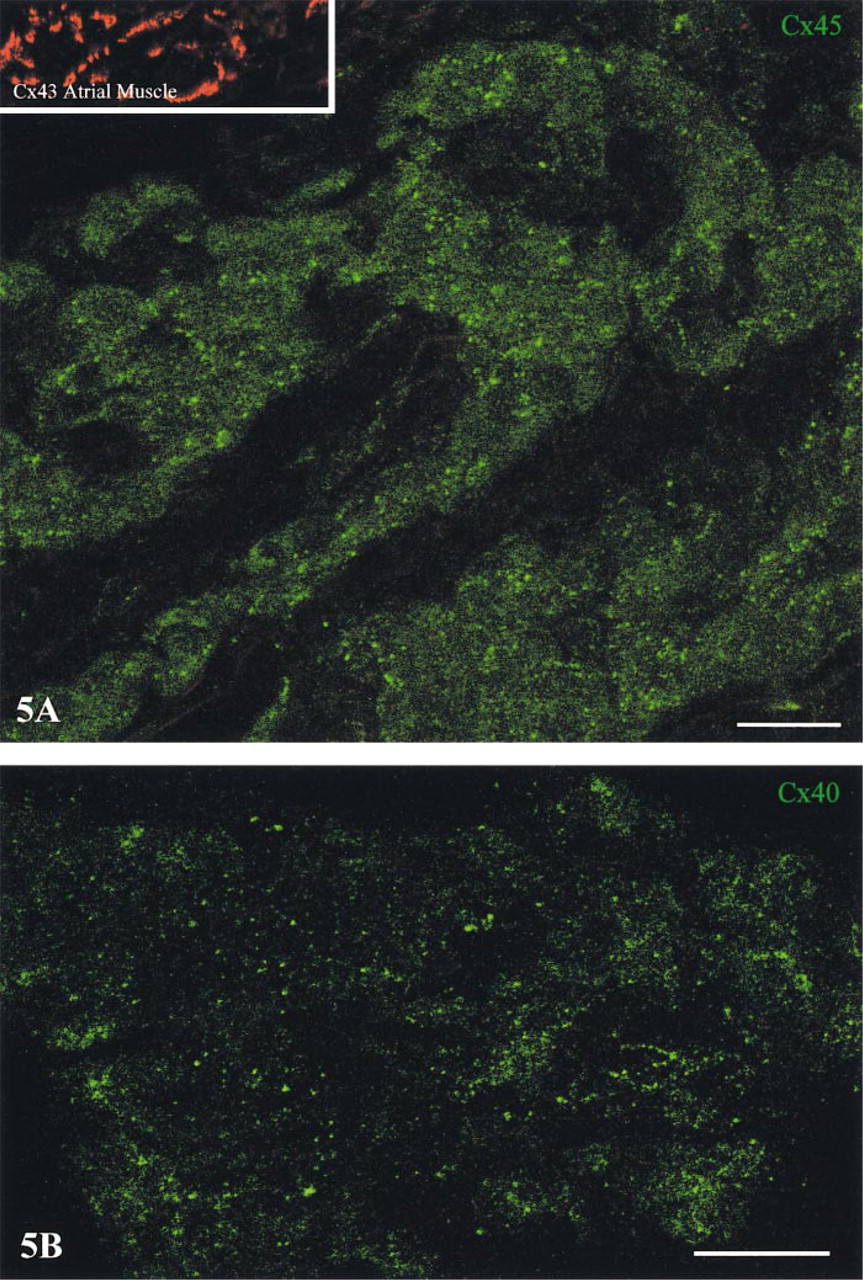
Confocal micrographs showing localization of Cx45 (
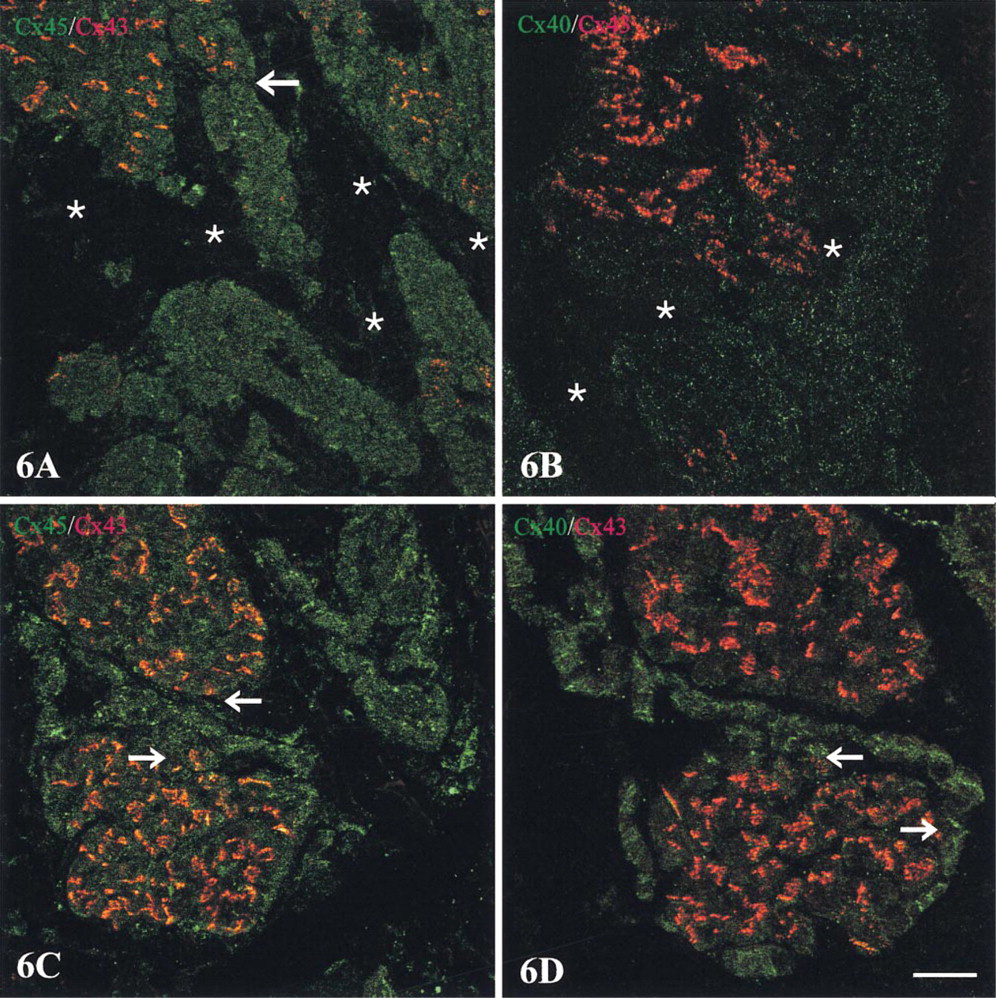
Confocal micrographs of sections through a SA node preparation double labeled for Cx45 with Cx43 and for Cx40 with Cx43. For orientation of the areas of the preparation from which these fields are taken, refer to Figure 4. (
In all experiments, the specificity of the localization was confirmed by peptide inhibition and by controls in which (a) the primary antibody was omitted and (b) treatment with a single primary antibody was followed by the nonmatching secondary antibody or both secondary antibodies.
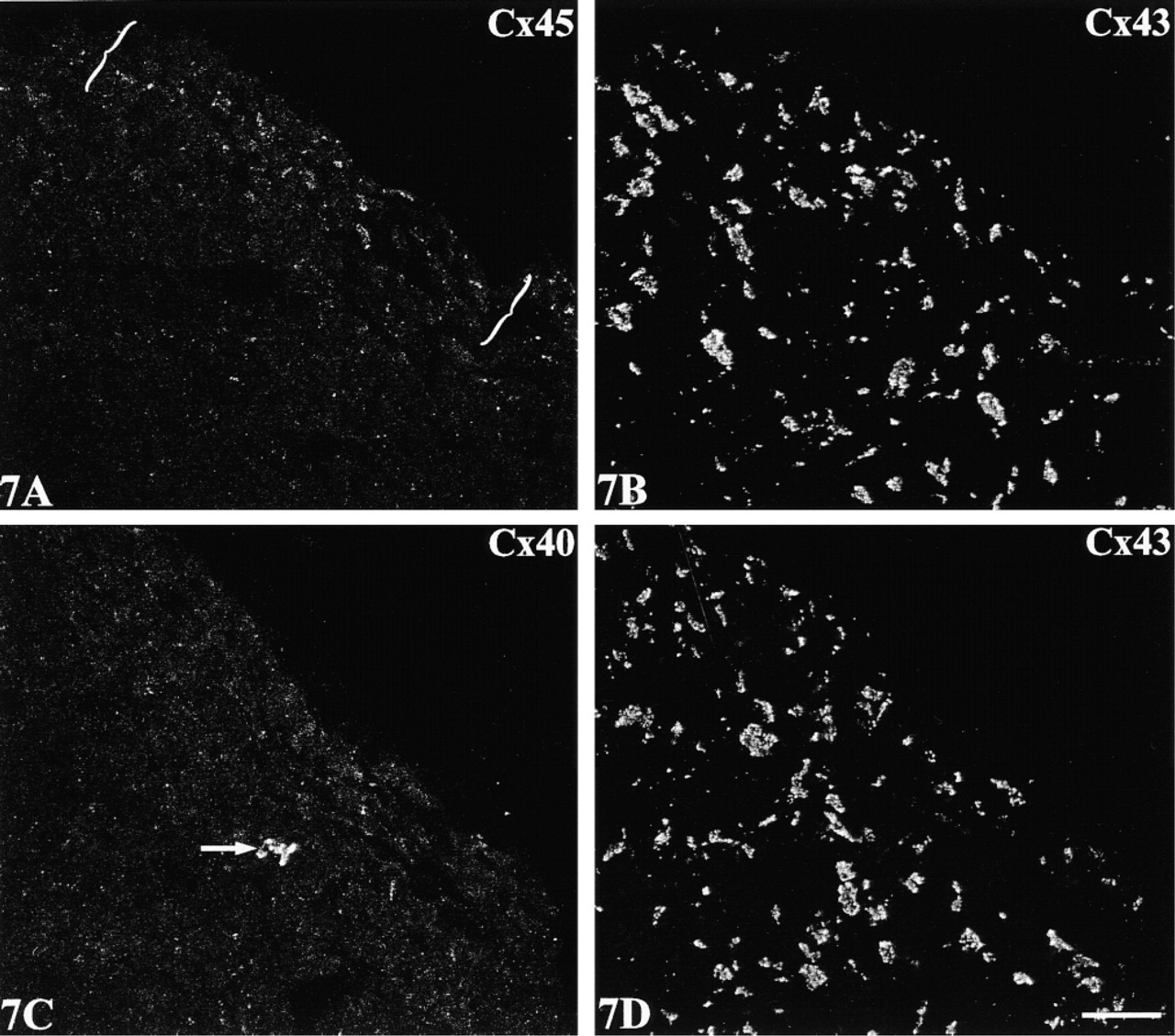
Double label, dual-channel scanning confocal microscopy demonstrates a conspicuous region of co-expression of Cx45 (
Discussion
The principal features of the spatial distribution of connexins 45, 43, and 40 in the SA nodal region of the rabbit, as determined from the double label confocal imaging results, are summarized diagrammatically in Figure 9. Key findings are that Cx45 and Cx40, but not Cx43, are expressed in the central SA node and that distinctive zones of differential connexin expression exist at the SA nodal/crista terminalis border. The border zones are of two types; a sharply demarcated boundary between Cx43 and Cx45/Cx40 and a transitional zone adjacent to the right branch of the SA ring bundle in which Cx43 and Cx45 expression merges. These findings provide new insights into a possible structural basis for the unique electrophysiological properties of the SA node.
Previous studies on connexin distribution in the SA node in different species present a confusing picture. Our finding of a lack of immunodetectable Cx43 in rabbit SA node contrasts with earlier reports in both rabbit (Anumonwo et al. 1992) and hamster (Trabka-Janik et al. 1994) but accords with the consensus in a series of other species, including rat, guinea pig, cow, and human (Van Kempen et al. 1991; ten Velde et al. 1992; Oosthoek et al. 1993; Davis et al. 1994, 1995; Opthof 1994). In dog and human, the SA node was reported to be Cx43-negative but Cx40- and Cx45-positive (Davis et al. 1994, 1995), but a more recent study on dissociated canine SA nodal cells presents a more complex picture, with the majority (∼55%) of nodal cells reported to express only Cx40, approximately one third expressing all three connexins, and the remainder none (Kwong et al. 1998). Although species-related differences in connexin expression profile of the SA node may well exist, technical differences, including reliability and specificity of antibody probes and quality and interpretation of the image data, have undoubtedly contributed to the discrepancies in conclusions from different laboratories.
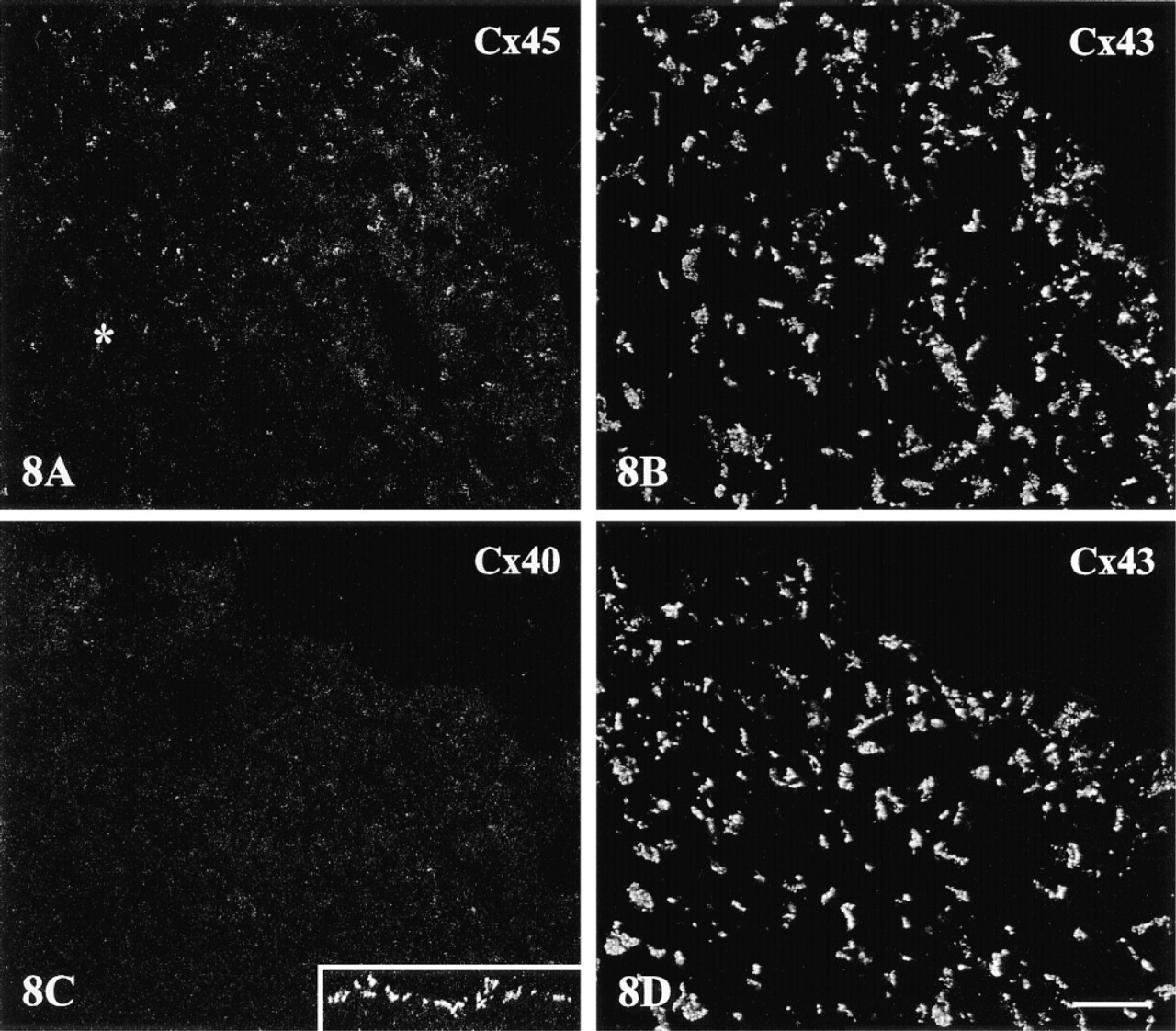
A further example of the merged zone of Cx45/Cx43 expression at the endocardial side of the crista terminalis, to demonstrate reproducibility of the findings and the variation in width of the co-expression zone observed from one heart to the next. Dual imaging of Cx45 (
By using a new, rigorously characterized Cx45 antibody (Coppen et al. 1998) for immunoconfocal microscopy, we now demonstrate that distinct small Cx45 gap junctions are scattered throughout the Cx43-negative region of the rabbit central SA node.
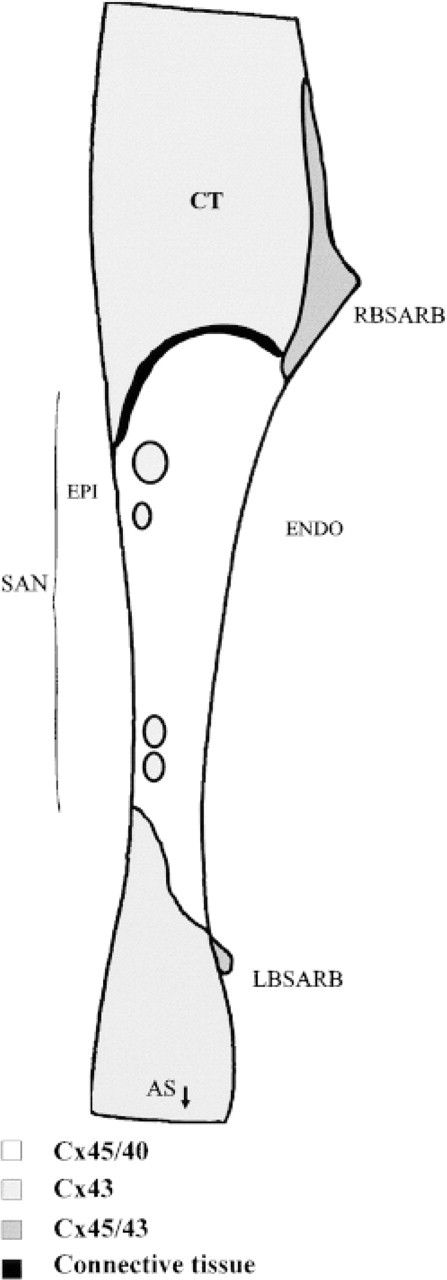
Diagrammatic summary of the spatial patterns of Cx43, Cx40, and Cx45 distribution in the SA node and adjacent regions of the rabbit heart. CT, crista terminalis.
Immunolabeled Cx40 gap junctions of similar size and appearance are also present in this region. The dimensions and quantities of the Cx45 and Cx40 spots observed are consistent with the size and frequency of SA nodal cell gap junctions, as revealed by electron microscopy (Masson-Pevet et al. 1979; Bleeker et al. 1980; De Mazière et al. 1992). We therefore conclude that Cx45 and Cx40 are the major connexins of the rabbit SA node. Data from in vitro expression systems suggest that gap junction channels composed of mouse or rat Cx40 have high unitary conductances compared with those made from Cx43 (Traub et al. 1994; Beblo et al. 1995; Bukauskas et al. 1995), whereas Cx45 channels reportedly have low conductance values (Veenstra et al. 1994; Moreno et al. 1995). Given the presence of two connexins (Cx45 and Cx40) of markedly different unitary conductances, the small size and sparsity of the junctions in the SA node, rather than the type of connexin present, appear to be major factors in the very low intercellular coupling conductance of the central pacemaker region demonstrated from electrophysiological studies.
Theoretical models have proposed that successful pacing and conduction of the action potential into the surrounding atrial tissue requires a central zone of weakly coupled cells, surrounded by a ring across which a gradient of increased coupling occurs (Joyner and Van Capelle 1986). However, the demonstration in earlier studies of sharply defined Cx43-positive and Cx43-negative zones led to questioning of the coupling gradient concept and, instead, a role for interdigitations of atrial muscle into SA nodal tissue was proposed (Oosthoek et al. 1993; ten Velde et al. 1995).
Although some interdigitation was observed in the present study, this feature did not appear to be as pronounced in the rabbit as previously reported in the bovine, human, and guinea pig heart (Oosthoek et al. 1993; ten Velde et al. 1995).
Our ability to image pairs of connexin types simultaneously sheds new light on the possible mechanism by which the pacemaker is protected from the hyperpolarizing influence of the surrounding tissue. Although the border between the SA node and crista terminalis presents a potentially large area of contact, our findings indicate that the two tissues at this site remain compartmentalized in terms of connexin expression and with a discontinuous connective tissue layer, apart from a single conspicuous region of contact characterized by Cx43 and Cx45 expression. Such an arrangement could achieve both minimization of contact between the tissues, thereby restricting the hyperpolarizing influence of the atrial muscle on the SA node, and maintenance of a communication route for directed exit of the impulse into the crista terminalis. The differential patterns of connexin expression we show here, comprising predominantly Cx43 expression in the atrial muscle, Cx43 and Cx45 co-expression in the region of contact (at the periphery of the SA node), and Cx45/Cx40 co-expression in the SA node itself, may thus provide the morphological basis of the gradient of coupling proposed by Joyner and van Capelle (1986). The significance of Cx45/Cx43 co-expression in the right and left branches of the SA ring bundle is unclear but may be related to their common developmental origin (both are remnants of the venous valve in the fetus).
Our ability to demonstrate a transitional zone of connexin expression at the nodal periphery depended critically on the availability of a reliable, specific Cx45 antibody and the ability to undertake double labeling combined with dual-channel scanning for simultaneous visualization of Cx45 and Cx43. Previous studies concluding the absence of a morphological gradient in gap junction coupling did not examine connexins other than Cx43 (Oosthoek et al. 1993; ten Velde et al. 1995). Interpretation of earlier reports that Cx45 is expressed in the SA node of dog and human (Davis et al. 1994, 1995; Kwong et al. 1998) is confounded by the use of an antibody raised against the same sequence as a Cx45 antibody now known to crossreact with Cx43 (Coppen et al. 1998). Such crossreactivity would clearly impede discrimination of any transitional boundary regions.
Mounting evidence suggests that connexin compatibility is a key mechanism for regulating intercellular communication and coupling between tissue compartments of different function. Cx45 has been shown to alter channel properties when transfected into cells that endogenously express Cx43 (Koval et al. 1995). In cells expressing more than one connexin type, a multiplicity of different arrangements involving heterotypic channels, homotypic channels, or heteromeric channels is theoretically possible (Kumar and Gilula 1996; White and Bruzzone 1996). Our finding that Cx43 and Cx45 signals co-localize at a specific zone at the SA nodal/crista terminalis border suggests that at least some individual gap junctions in these zones contain both connexin types. Although we cannot at present determine whether heterotypic Cx45/Cx43 channels are formed in these regions, it has been established from in vitro studies that connexons made from Cx45 are able to dock with connexons made from Cx43 to establish functional heterotypic channels (White et al. 1995; Bruzzone et al. 1996). Some heterotypic combinations of connexon (including those involving Cx43 and Cx45) show asymmetric voltage dependence, a property resulting in formation of rectifying channels (White et al. 1994; Bruzzone et al. 1996). Further investigation is required to determine precisely how the novel connexin expression patterns identified in the SA node and its borders combine with other features of tissue architecture and protein channel expression to determine the intercellular coupling properties of the SA node.
Footnotes
Acknowledgements
Supported by project grants (nos. PG/93136 and PG/97175; NJS) and a program grant (no. RG/96004; MRB) from the British Heart Foundation and by a grant from the Ministry of Education, Science and Culture of Japan.
We thank Dr Robert Gourdie (Medical University of South Carolina) for the anti-Cx40 antiserum and Stephen Rothery for invaluable assistance with animal work and immunoconfocal microscopy.