
Abstract
Integration of vascular endothelial function relies on multiple signaling mechanisms, including direct cell-cell communication through gap junctions. Gap junction proteins expressed in the endothelium include connexin37, connexin40, and connexin43. To investigate whether individual endothelial cells in vivo express all three connexin types and, if so, whether multiple connexins are assembled into the same gap junction plaque, we used affinity-purified connexin-specific antibodies raised in three different species to permit multiple-label immunoconfocal and immunoelectron microscopy in the rat main pulmonary artery. Immunoconfocal microscopy showed a high incidence of co-localization between connexin43 and connexin40, but lower incidences of co-localization between connexin37 and connexin40 or connexin43. Immunoelectron microscopy revealed that 83% of gap junction profiles contained all three connexins, with the proportion of connexin40 labeling being significantly higher than that of connexin37 or connexin43. The presence of three different connexin types of distinct properties in vitro provides potential for complex regulation and functional differentiation of endothelial intercellular communication properties in vivo.
I
The gap junction channel is constructed from a pair of hemichannels termed connexons, each connexon being assembled from six connexin molecules. The connexins form a multigene family of conserved proteins, at least 13 members of which are expressed in mammalian cells (Bruzzone et al. 1996; Kumar and Gilula 1996). Because cells stably transfected with cDNAs encoding different connexins show distinctive properties (e.g., permeability and unitary channel conductance) (Veenstra et al. 1994; Elfgang et al. 1995; Mazzoleni et al. 1996;Veenstra 1996; Cao et al. 1998), the precise functional properties of gap junctions in vivo are believed to depend in part on the specific connexins present.
In common with many other tissues, endothelia may express more than one connexin type (Christ et al. 1996; Severs et al. 1996; van Rijen et al. 1997; Yeh et al. 1997). Earlier reports identified connexin43 (Cx43) and connexin37 (Cx37) as the principal connexins in endothelial cells from a variety of sources (Larson et al. 1990; Pepper et al. 1992; Reed et al. 1993; Little et al. 1995). In addition, connexin40 (Cx40) has been reported in some endothelia, e.g., those of the rat aorta, small muscular arteries in heart and lung, and pial arterioles in brain (Bastide et al. 1993; Bruzzone et al. 1993; Little et al. 1995). Recent studies on cultured cells emphasize that the pattern of connexin expression in vitro is profoundly influenced by cell density, mechanical load, growth, mitosis, and the presence of exogenous cytokines (Pepper and Meda 1992; Larson et al. 1997; Xie et al. 1997; Cowan et al. 1998; van Rijen et al. 1998). Our recent work on intact arterial tissues demonstrates at least some intrinsic variation in endothelial connexin expression patterns according to vascular site (Yeh et al. 1997). Specifically, in the rat, the coronary artery endothelium expresses Cx40 and Cx37 but no detectable Cx43 (Yeh et al. 1997), whereas the endothelia of the aorta (Yeh et al. 1998) and the pulmonary artery (Ko et al. 1998) express all three of these connexins. In this last study, we reported data from single label experiments on pulmonary arterial endothelium using antibodies raised in different species, as a first step in establishing the feasibility of developing double and triple labeling for the simultaneous visualization of up to three connexin types (Ko et al. 1998). The present study builds on these findings by reporting new information on the precise connexin make-up at the level of the individual gap junction from correlative multiple-labeling immunoconfocal and immunoelectron microscopy applied to pulmonary artery endothelium.
Materials and Methods
Tissue Preparation
All studies were conducted on the main pulmonary artery obtained from adult male Sprague-Dawley rats (320-450 g). Samples for immunoconfocal microscopy were obtained by rapidly dissecting arterial samples from animals sacrificed by dislocation of the neck. After rinsing with PBS containing heparin (10 U/ml), the samples were quickly cut into two segments, frozen immediately in isopentane cooled with liquid nitrogen, and stored in liquid nitrogen. Ten-μm-thick cryosections of the arterial segments were cut transversely, mounted on poly-L-lysine-coated slides, and dried overnight in a −20C freezer before immunolabeling. Samples for postembedding immunogold thin-section electron microscopy were prepared by perfusion fixation. The rats were anesthetized by IP injection of midalozam-hypnorm and perfusion-fixed with 2% paraformaldehyde in PBS retrogradely via the abdominal aorta for 15 min, and the main pulmonary arteries dissected out. Fixation of small tissue samples was continued in the same solution for a further 15 min at room temperature (RT). The fixed samples were dehydrated in 30% ethanol at 4C for 30 min, followed by 50% and 70% ethanol at −20C for 30 min and 1 hr, respectively, and then 90% and absolute ethanol three times at −30C with 1 hr for each step. Infiltration with Lowicryl K4M (Agar Scientific; Stansted, UK) was carried out using 1:1 and then 2:1 Lowicryl:ethanol mixtures, followed by pure Lowicryl K4M overnight. After being placed in a new batch of Lowicryl K4M for 1 hr, the samples were embedded in fresh Lowicryl K4M in gelatin capsules and polymerized with UV light at −30C for 16 hr, and then at RT for 72 hr in a Balzers FSU 010 low temperature-embedding unit. Preparation of rat tissues was conducted in accordance with the United Kingdom Animals (Scientific Procedures) Act, 1986.
Anti-connexin Antibodies and Secondary Antibody Detection Systems
Three primary antibodies were used for immunohistocytochemical detection of the gap junction proteins Cx37, Cx40, and Cx43. Those against Cx37 and Cx40 were affinity-purified polyclonal antisera raised in rabbits and guinea pigs, respectively. The Cx37 antiserum Y16Y(R4) was raised against a synthetic peptide corresponding to residues 266-281 of the cytoplasmic C-terminal tail of the rat Cx37 sequence. The Cx40 antiserum V15K(GP319) was produced against a synthetic peptide corresponding to residues 256-270 of the cytoplasmic C-terminal tail of the rat Cx40 sequence. These antibodies were affinity-purified and fully characterized by Western blotting and immunolabeling of transfected cells (Yeh et al. 1998). For Cx43, we used a commercially available mouse anti-Cx43 monoclonal antibody (MAb) directed against residues 252-270 of the rat Cx43 sequence (Chemicon; Harrow, UK).
For immunoconfocal microscopy, the working dilution of the anti-Cx37 antibody was 1:60 (0.5 μg/ml) and that of the anti-Cx40 antibody was 1:100 (0.3 μg/ml). The working solution of the commercial anti-Cx43 antibody was 1:1000. The secondary antibody detection systems used were donkey anti-mouse, anti-guinea pig, and anti-rabbit immunoglobulins conjugated to either Cy3 or Cy5 (Chemicon; dilution 1:500). Both these primary antibodies and secondary antibody detection systems were diluted in 0.5% bovine serum albumin (BSA) in PBS.
For immunoelectron microscopy, the optimal dilution of the anti-Cx37 antibody was 1:30 (1 μg/ml) and that of the anti-Cx40 antibody was 1:100 (0.3 μg/ml), used in 0.5% BSA in PBS. The working solution of the commercial anti-Cx43 antibody was 1:500. The secondary antibody detection systems used were 5-nm gold-goat anti-rabbit complexes, 10-nm gold-goat anti-guinea pig complexes, and 15-nm gold-goat anti-mouse complexes (BioCell; Cardiff, UK). All secondary antibodies were diluted 1:50 in PBS.
Immunoconfocal Microscopy
Cryosections were blocked with 0.5% BSA in PBS for 30 min and incubated in the primary antibody of choice. Double labeling for Cx37 with Cx40, or for Cx37 with Cx43, was done by first incubating sections with the anti-Cx37 primary antibody overnight at RT and then with the anti-Cx40 primary antibody for 1 hr or with the anti-Cx43 primary antibody for 2 hr at 37C. For Cx40/Cx43 double labeling, the sections were incubated in a mixture of anti-Cx40 and anti-Cx43 antibodies overnight at RT. The sections were washed in PBS and incubated in the matching secondary antibody or mixture of secondary antibodies for 1 hr at RT before final washing and mounting. Negative controls included (a) omission of the primary antibody, and (b) for double labeling, using each primary antibody with both matching and non-matching secondary antibodies. All secondary antibodies were confirmed to be species-specific to their individual primary antibody. Peptide inhibition controls were performed for the polyclonal antibodies (Yeh 1997).
The immunofluorescence-labeled sections were examined by confocal laser scanning microscopy using a Leica TCS 4D equipped with an argon-krypton laser and fitted with the appropriate filter blocks for detection of Cy3 and Cy5 fluorescence. The images were taken using simultaneous dual-channel scanning and were transformed into projection views using sets of five consecutive single optical sections taken at l-μm intervals. All specimens were examined within 24 hr of immunolabeling.
Five dual-channel projection images from each of five rats were randomly selected to evaluate the distribution and co-localization of connexins on pulmonary arterial endothelial gap junctions. The dual-channel images were further split into two separate single-channel images corresponding to each of the connexin types. Fifty immunolabeled spots were randomly selected from each dual-channel image and the component connexins of each spot were determined by analyzing the corresponding split images. A spot on the dual-channel image that was visualized as a corresponding spot on each of the two split images (i.e., both connexins were labeled) was classified as showing co-localization. If a spot was present on only one of the pair of split images, the spot was classified as containing only the single connexin type identified. In each set of dual-channel and corresponding split images, the percentage of spots showing co-localization was determined. A further analysis involved taking those spots that were positive for a given connexin type and determining the proportion of these spots that were also positive for the second connexin type. Data are presented as mean percent values ± SEM.
Postembedding Immunogold Thin-section Transmission Electron Microscopy
Ultrathin sections of the Lowicryl K4M-embedded specimens were prepared using glass knives. The sections were picked up on nickel grids and incubated at RT successively in 1% BSA in PBS for 5 min, 1% gelatin in PBS for 10 min, and 0.02 M glycine in PBS for 3 min. Sections were immunolabeled as follows. For single labeling, the sections were incubated in the primary antibody, washed with PBS, and then labeled with the appropriate matching secondary antibody-gold complex. The optimal incubation periods determined for the anti-Cx37, anti-Cx40, and anti-Cx43 primary antibody treatments were overnight, 2 hr, and 4 hr, respectively. For double labeling of Cx37 with Cx40 and of Cx37 with Cx43, sections were incubated in anti-Cx37 primary antibody overnight and then in anti-Cx40 primary antibody for 2 hr or in anti-Cx43 primary antibody for 4 hr. For double labeling of Cx40 with Cx43, the two primary antibodies were used as a mixture, with a treatment period of 4 hr. For triple labeling, sections were first incubated in anti-Cx37 primary antibody overnight and then in a mixture of anti-Cx40 and anti-Cx43 primary antibodies for 4 hr. For double and triple labeling, the secondary antibodies were applied as mixtures. An incubation period of 1-1.5 hr was used for the secondary antibody step for single, double, and triple labeling protocols. All immunolabeling steps were carried out at RT. After immunolabeling, the sections were washed with PBS, incubated in 1.25% glutaraldehyde for 3 min, further washed with distilled water, dried, and then stained with uranyl acetate and lead citrate. All sections were examined in either the Philips EM301 or the Hitachi 900 electron microscope. Negative controls consisted of (a) omission of primary antibodies, (b) application of irrelevant primary antibodies or serum [mouse monoclonal anti-dystrophin MAb (Stevenson et al. 1998), rabbit polyclonal anti-desmoplakin antibody (Angst et al. 1997), and guinea pig preimmune serum)]; and (c) single primary antibody followed by double and triple secondary antibody incubation (i.e., matching and nonmatching secondaries). All irrelevant primary antibodies and serum were demonstrated not to label gap junctions, and all three secondary antibodies were confirmed to be specific to their individual primary antibodies and showed no crossreaction in the multiple-labeling experiments.
To evaluate the distribution and co-localization of gold label for the different connexins in individual endothelial gap junction plaques, 35 gold-labeled gap junctions were randomly selected from five triple labeled sections. For each gap junction, gold markers of each size were counted and the proportions of each calculated. Results are presented as mean percent values ± SEM, and data were compared using the Wilcoxon rank-sum test with statistical significance defined as p<0.05.
Results
Immunoconfocal Microscopy
By immunoconfocal microscopy, rat main pulmonary artery endothelial gap junctions were visualized as distinct punctate labeling at the luminal side of the internal elastic laminae (Figure 1). Whereas Cx40 and Cx43 endothelial labeling was abundant and evenly distributed, Cx37 labeling was less marked and was heterogeneously distributed. The spatial distribution of each pair of the three connexin types, as assessed by double labeling and dual-channel recording, showed a high incidence of co-localization between Cx40 and Cx43 spots (78.4 ± 0.64%) but only lower incidences of co-localized Cx37 and Cx40 spots (44.8 ± 1.78%) and of Cx37 and Cx43 (38.7 ± 1.88%) spots. Analysis of the proportions of co-localized and single labeled spots for each connexin type demonstrated that most Cx37-positive spots were also positive for Cx40 (84.4 ± 0.56%) and for Cx43 (75.7 ± 1%). The majority of Cx40-positive spots were also positive for Cx43 (88.5 ± 0.32%), although only about half the Cx40-positive spots were also positive for Cx37 (49.9 ± 2%). For Cx43 spots, a pattern was observed similar to that of Cx40 (87.2 ± 2.7% positive for Cx40 and 44.4 ± 11.4% positive for Cx37).
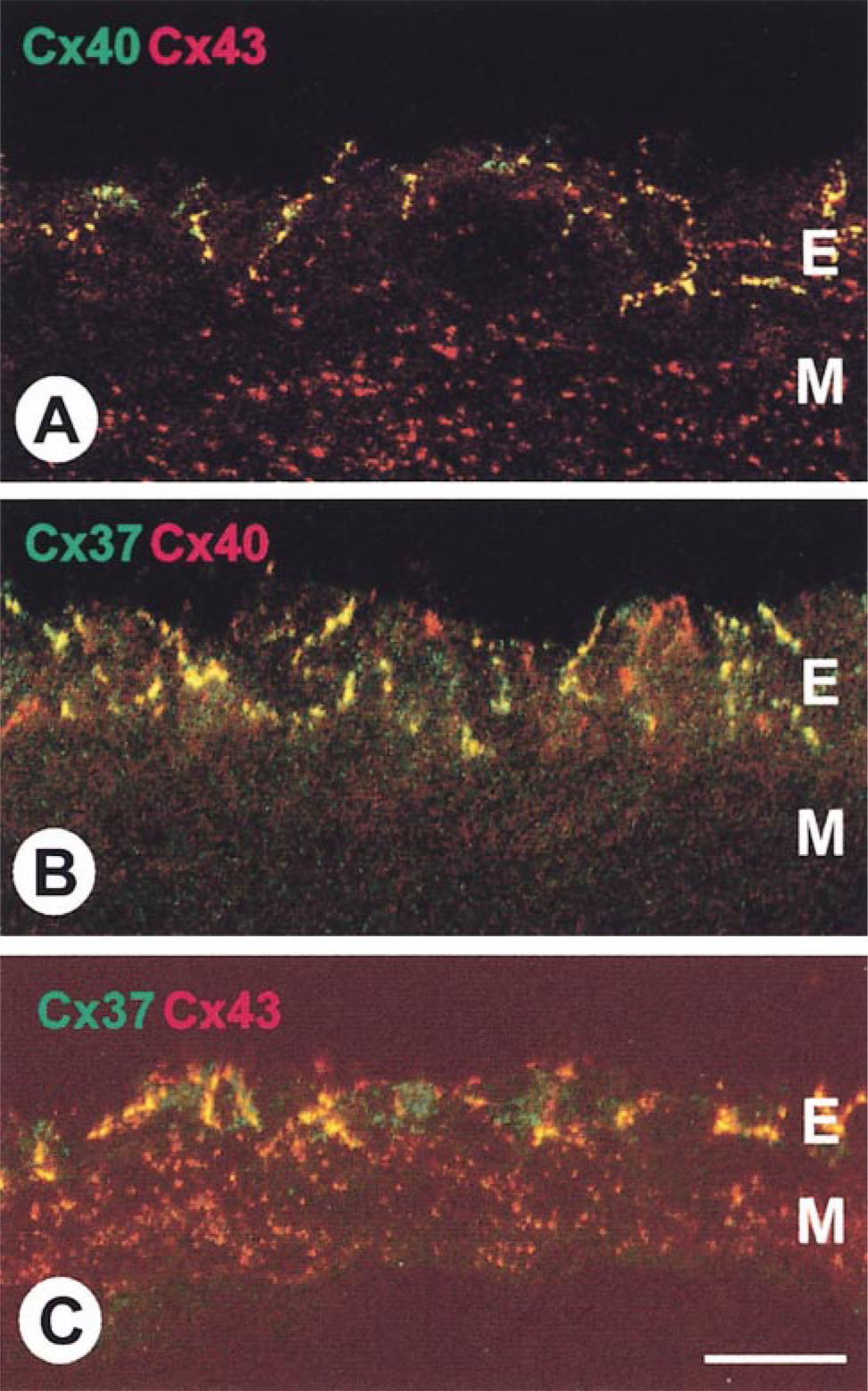
Double labeling for connexin40 (Cx40) and connexin43 (Cx43) (A), connexin37 (Cx37) and connexin40 (B), and connexin37 and connexin43 (C) in rat main pulmonary artery viewed by immunoconfocal microscopy. Yellow spots represent sites at which the signals (red and green fluorescence) for the individual connexins are co-localized. Whereas Cx40 and Cx43 were extensively co-localized in the endothelium (E) (A), Cx37 showed lower incidences of co-localization with Cx40 (B) and Cx43 (C). In the medial layers (M), only Cx43 is expressed, but not Cx37 or Cx40. Bar = 10 μm.
Postembedding Immunogold Thin-section Transmission Electron Microscopy
Thin sections of Lowicryl K4M-embedded rat main pulmonary arteries revealed endothelial gap junctions as typical pentalaminar structures formed from the adjacent cell membranes of neighboring cells (Figures 2 and 3). Under the optimized conditions used, the majority of gap junctions in the single labeled preparations were heavily decorated with gold particles specifically along the junctional membranes, with minimal background labeling elsewhere in the section (Figure 2). Negative controls consistently showed no labeling. The few gap junction profiles that remained unlabeled in the experimental samples may have represented junctions within the section, unexposed at the surface and thus inaccessible to antibodies. The single labeling experiments showed that the three different sizes of gold markers used to discriminate each connexin type were readily distinguishable from one another and therefore suitable for multiple labeling.
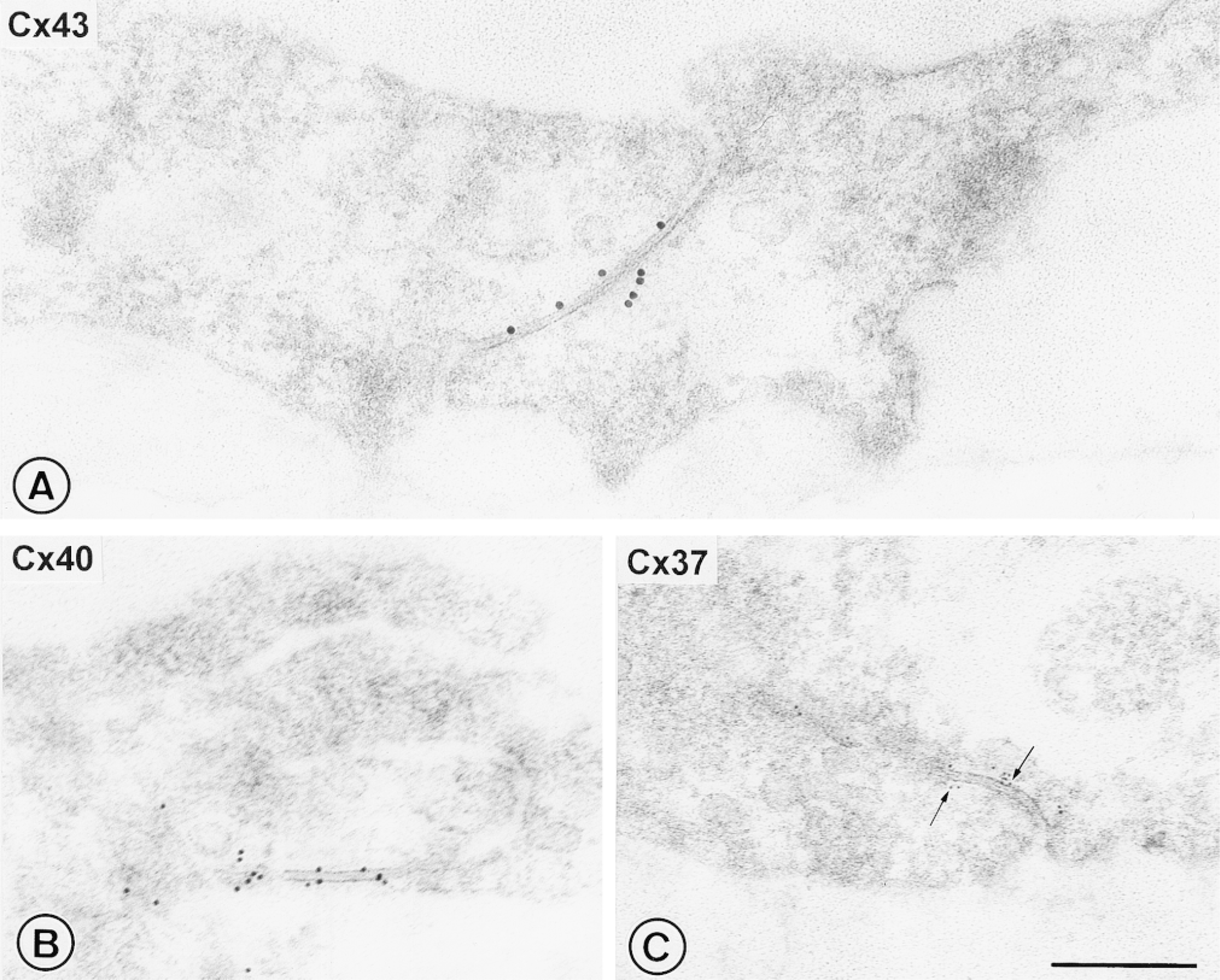
Immunogold transmission electron microscopy of single labeled connexins in sections of Lowicryl-embedded rat main pulmonary artery. The endothelial gap junctions with typical pentalaminar structure were decorated with gold particles specifically along the junctional membranes, with minimal background labeling. The three different sizes of gold markers used to discriminate each connexin type were readily distinguishable from one another. (A) With 15-nm gold for Cx43; (B), with 10-nm gold for Cx40; (C) with 5-nm gold for Cx37 (arrows). Bar = 250 nm.
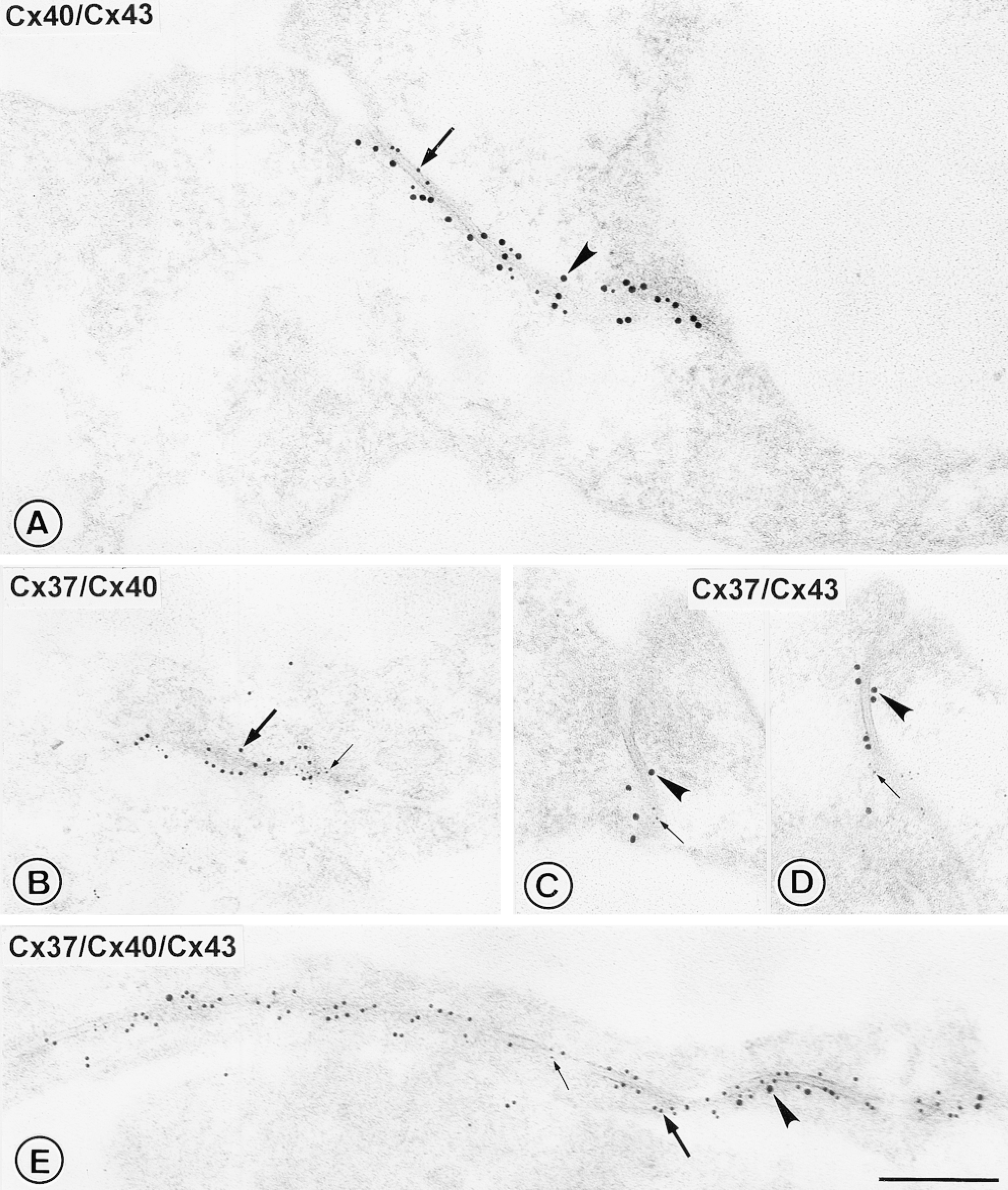
Multiple immunogold labeling for connexins in sections of Lowicryl-embedded rat main pulmonary artery. (A-D) Results of double labeling for pairs of connexin types using the same sizes of markers as used in Figure 2. The pairs of different-sized gold markers were co-localized specifically on the endothelial gap junctions and were readily distinguishable from each other. No segregation of different sizes of markers across the junction, or major lateral aggregation, is apparent. (E) Results of triple labeling. The presence of all three sizes of gold markers labeling individual gap junctions such as this demonstrates that connexins 40, 43, and 37 are organized within the same junction. Arrowheads, Cx43-15-nm gold; thick arrows, Cx40-10-nm gold; thin arrows, Cx37-5-nm gold. Bar = 250 nm.
Double labeling using the same sets of gold markers for each connexin type as those used in single labeling clearly demonstrated co-localization of Cx40 with Cx37, Cx43 with Cx37, and Cx40 with Cx43 within individual gap junctions (Figures 3A-3D). The different-sized gold markers were readily distinguishable and showed no segregation transversely across the junction or major lateral aggregation, suggesting that mixtures of connexins were present more or less evenly throughout each of the junctional membranes.
On triple labeling, 83% of the labeled gap junction membranes viewed demonstrated the presence of all three sizes of gold markers (Figure 3E). Some variation was apparent in the relative amounts of gold markers for the different connexin types (Figure 4). The proportion of Cx40-10-nm gold labeling (52.7 ± 4.0%) was significantly higher than that of Cx37-5-nm gold labeling (20.5 ± 2.6%; p = 0.0001) and Cx43-15-nm gold labeling (26.8 ± 3.8%; p = 0.003). Of the few gap junction membranes containing only one or two sizes of gold markers, one gap junction contained markers for only one connexin type (Cx43) and five contained markers for two connexin types (one Cx37 with Cx40; four Cx40 with Cx43; n = 35). For all multiple-labeling experiments, background labeling was negligible, and double labeling for each pair of primary antibodies with all three secondary antibody-gold complexes confirmed specificity of the localization.
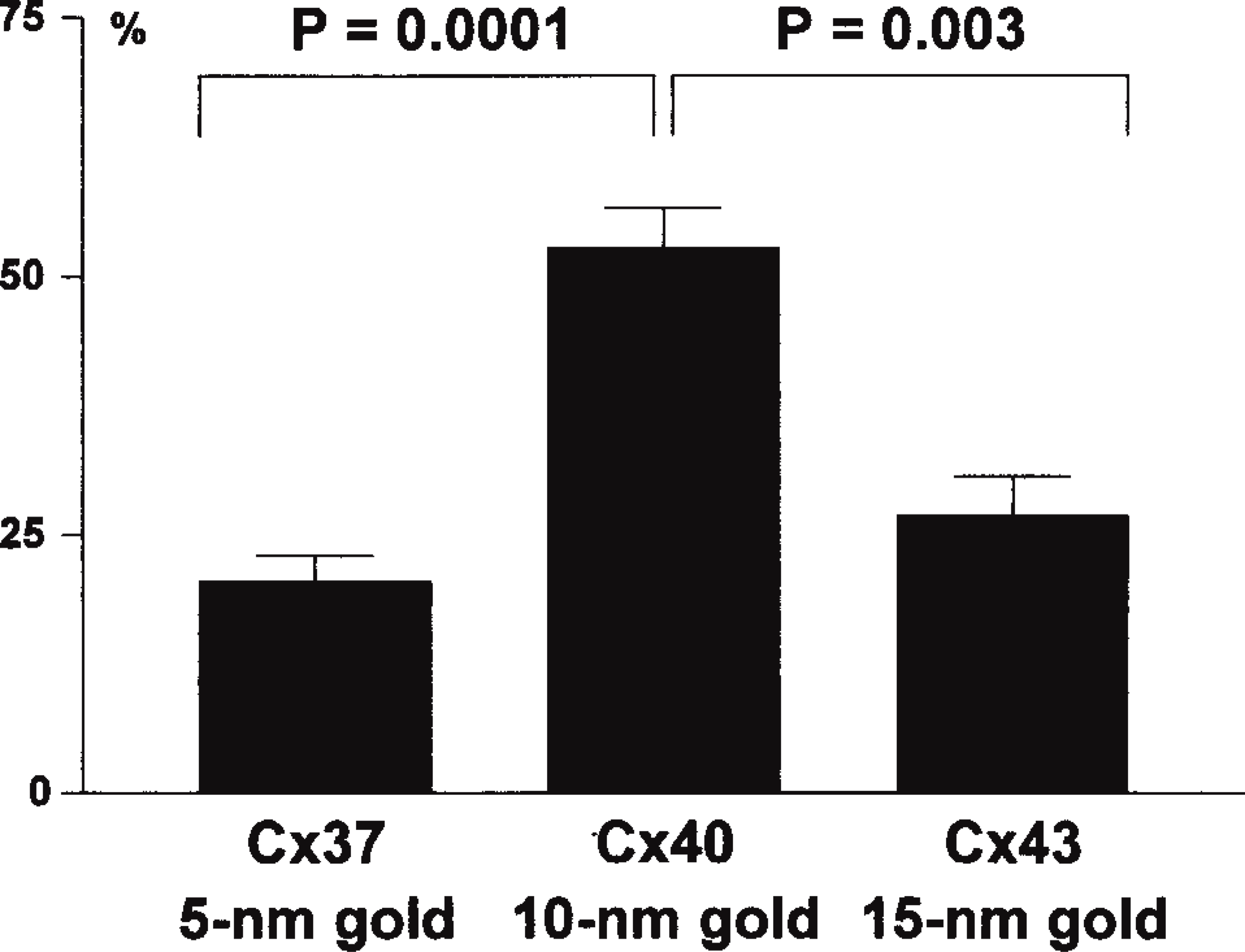
Quantitative analysis of the proportions of gold marker for each connexin type from triple labeled endothelial gap junctions.
Discussion
This study demonstrates that Cx40, Cx43, and Cx37 are simultaneously expressed in endothelial cells of the main pulmonary artery of the rat and that, in this tissue, the majority of individual gap junction plaques contain all three connexins. Although a number of tissues are known to express more than one connexin type [e.g., Cx40 and Cx45 in the cardiac conduction system (Coppen et al. 1998) and Cx46 and Cx50 in lens (Paul et al. 1991)], the extent of connexin diversity in specific types of endothelial cells has remained unclear. Earlier studies by Northern blotting, Western blotting, and immunocytochemistry have shown that Cx37, Cx40, and Cx43 are expressed in diverse vessels, but these studies have provided only limited data for any given endothelial cell type, the various reports differing in the precise connexin profiles identified in endothelial cells of different origins (Larson et al. 1990; Pepper et al. 1992; Bastide et al. 1993; Bruzzone et al. 1993; Reed et al. 1993; Little et al. 1995). Because most previous immunofluorescence work on endothelial connexins has relied on single labeling, it has previously been difficult to ascertain the extent to which multiple connexins are expressed by the same endothelial cell and, importantly, whether endothelial gap junctions are composed of mixed or single connexin types.
To address this issue, the present study has built on our earlier studies (Yeh et al. 1997; Ko et al. 1998) by developing elaborate multiple-labeling techniques for complementary immunoconfocal and immunoelectron microscopy, in particular, triple immunogold labeling at the electron microscopic level. Although correlation of our data from immunoconfocal and electron microscopy gives broad agreement on the overall patterns of distribution and co-localization among the three connexins, comparison of the quantitative data from these two techniques does reveal some apparent discrepancies, particularly in relation to the extent of co-localization of Cx37 with the other connexins. Although fewer than 60% of the immunofluorescent gap junction spots were positive for Cx37, immunoelecpicture tron microscopy indicated that >80% of labeled junction profiles showed gold markers for all three connexin types. This suggests that, for a given set of antibodies, the sensitivity of detection is higher in immunogold electron microscopy than in immunofluorescence confocal microscopy. Absence of detectable signal for one connexin type in double label immunoconfocal microscopy therefore does not exclude the presence of low amounts of that connexin. Furthermore, immunofluorescence localization strictly permits only detection of the target protein; it does not directly visualize the gap junction itself. Although multiple-label immunoconfocal microscopy is certainly valuable in screening for the possible existence of co-localization and estimating its frequency, multiple-label immunogold electron microscopy provides the “gold standard” for definitive localization of two or more connexins to the same morphologically defined gap junction. We suggest that the relative abundance of gold markers for the three connexins, in the order Cx40>Cx43>Cx37, is likely to reflect, at least in part, the relative amounts of the three connexin types within the junction, although differences in antibody affinities and epitope accessibilities, as well as technical aspects (e.g., the order in which primary and secondary antibodies are applied in multiple-labeling experiments and the sizes of the gold markers used for each connexin) all potentially contribute to the absolute values observed. Bearing in mind that even the best antibody probes will never allow 100% labeling efficiency, and that each sectional view of a labeled gap junction represents only a thin slice sampled through a much larger plaque, our finding that >80% of triple labeled junctional profiles show some label for Cx37 will underestimate the true number of junctions that contain at least a trace of Cx37. Therefore, it can be concluded that most if not all endothelial gap junctions of the rat pulmonary artery typically contain all three connexins. Parallel studies indicate a similar pattern of organization in rat aortic endothelial cells (Yeh et al. 1998), suggesting that this pattern may be common to elastic arteries in general.
In contrast to our finding that Cx43 is widely expressed in the endothelium of rat pulmonary artery (this study) and aorta (Yeh et al. 1998), a recent study reports Cx43 signal specifically at sites of turbulent flow in rat aortic endothelium (Gabriels and Paul 1998). The apparent lack of Cx43 at sites distant from turbulent flow, as reported by Gabriels and Paul (1998), may reflect differences in sensitivity of detection among different antibodies and localization techniques. Cx43 may well show elevated expression at sites of turbulent flow, as reported by Gabriels and Paul (1998), but against a relatively lower background expression level elsewhere. Notwithstanding these apparent discrepancies within a given vessel type, the picture now emerging is that the endothelium expresses diverse connexin patterns according to location within the vascular tree (Yeh et al. 1997). For example, in contrast to pulmonary artery and aortic endothelia, the coronary artery endothelium expresses Cx40 and Cx37 but no detectable Cx43 (except towards its junction with the aorta) (Yeh et al. 1997).
In culture systems, expression of Cx37 and Cx43 appears to be differentially regulated according to growth status. Cx43 levels become elevated during growth but decline at confluency, whereas Cx37 shows the reverse pattern (Larson et al. 1997). In in vitro expression systems, distinctive molecular permeability, ionic selectivity, unitary conductance, and voltage gating properties are associated with gap junction channels composed of different connexin types (Moreno et al. 1994; Elfgang et al. 1995; Mazzoleni et al. 1996;Veenstra 1996; Cao et al. 1998). For example, of the three connexins expressed in the endothelium, Cx37 channels are reported to have very high unitary conductance values (300 pS), Cx43 channels relatively low conductance values (60 pS), and Cx40 channels intermediate values (150-200 pS) (Traub et al. 1994; Veenstra et al. 1994; Beblo et al. 1995; Bukauskas et al. 1995). Furthermore, the properties of endogenous channels can be altered by transfecting cells to express an additional connexin (Koval et al. 1995), and cotransfection to express two connexins gives channel properties different from those observed with single transfection (Brink et al. 1997). Differential expression of multiple connexins in endothelium in vivo would potentially thus offer scope for regulation of intercellular communication in response to diverse physiological stimuli in health and disease.
A question raised by our findings concerns the precise organization of the three connexins at the level of the gap junction channel. Our observation that the three sizes of gold markers were visualized with equal frequency on each side of the junction membrane raises the possibility of molecular arrangements involving heteromeric connexons (containing mixtures of connexins within the connexon) (Stauffer 1995; Jiang and Goodenough 1996; Brink et al. 1997; Diez et al. 1998), and heterotypic gap junction channels (i.e., channels in which one connexon made from one connexin type abuts a partner connexon made from a second connexin type) (Wolosin et al. 1997; Cao et al. 1998). In in vitro expression systems, connexons composed of Cx43 do not form functional channels with those composed of Cx40 (Bruzzone et al. 1993; Elf-gang et al. 1995) owing to inability to dock (Haubrich et al. 1996), but Cx37 channels are compatible with both Cx43 channels and Cx40 channels (Elfgang et al. 1995; Brink et al. 1997). Expression of Cx37 in endothelial cells might thus facilitate continuity of functional linkage in gap junctions containing the otherwise incompatible Cx43 and Cx40.
In conclusion, the present findings indicate that the connexin make-up of main pulmonary artery endothelial gap junctions, involving three connexin types capable of conferring different properties in vitro, provides inherent potential for complex regulation, functional differentiation, and versatility of endothelial intercellular communication properties in vivo. A clearer idea of the molecular structure of individual channels in terms of connexin composition, and their functional correlates, remains a challenge for future work.
Footnotes
Acknowledgements
Supported in part by project grants from the British Heart Foundation (grant nos. PG 97175 and 93136) and the Wellcome Trust (grant no. 046218/Z/95). Dr Yu-Shien Ko is a cardiologist from Chang Gung Memorial Hospital, Taipei, Taiwan, and gratefully acknowledges personal support from its Overseas Biomedical Scholarship Award.
We wish to thank Dr Robert Gourdie (Medical University of South Carolina, Charleston, SC) for providing the crude Cx40 antiserum and Dr J-P. Briand (IBMC, Strasbourg, France) for the gift of the Cx37 peptide.