
Abstract
To elucidate whether the two different gap junction proteins connexin43 (Cx43) and connexin26 (Cx26) are expressed and localized in a similar manner in the adult rat cochlea, we performed three-dimensional confocal microscopy using cryosections and surface preparations. In the cochlear lateral wall, Cx43-positive spots were localized mainly in the stria vascularis and only a few spots were present in the spiral ligament, whereas Cx26-positive spots were detected in both the stria vascularis and the spiral ligament. In the spiral limbus, Cx43 was widely distributed, whereas Cx26 was more concentrated on the side facing the scala vestibuli and in the basal portion. In the organ of Corti, Cx43-positive spots were present between the supporting cells but they were fewer and much smaller than those of Cx26. These data demonstrated distinct differences between Cx43 and Cx26 in expression and localization in the cochlea. In addition, the area of overlap of zonula occludens-1 (ZO-1) immunolabeling with Cx43-positive spots was small, whereas it was fairly large with Cx26-positive spots in the cochlear lateral wall, suggesting that the differences are not associated with the structural difference between carboxyl terminals, i.e., those of Cx43 possess sequences for binding to ZO-1, whereas those of Cx26 lack these binding sequences.
Keywords
G
In the cochlea, gap junctions were detected by electron microscopy (Jahnke 1975; Reale et al. 1975; Nadol et al. 1976; Forge 1984; Spicer and Schulte 1996), and the immunolocalization of connexin26 (Cx26), connexin30 (Cx30), and connexin31 (Cx31) has been reported (Kikuchi et al. 1995; Lautermann et al. 1998; Xia et al. 2000; Suzuki et al. 2001). It is proposed that gap junctions are assembled into two independent cellular networks in the cochlea, i.e., the epithelial and connective tissue gap junction networks (Kikuchi et al. 1995). Moreover, the crucial role of gap junctions in auditory functions was confirmed by the discovery that mutation of the Cx26 gene is responsible for some cases of human hereditary non-syndromic sensorineural deafness (Kelsell et al. 1997). After this discovery, mutations in additional connexins such as Cx30 (Grifa et al. 1999) and Cx31 (Xia et al. 1998; Liu et al. 2000) were implicated in human hereditary non-syndromic sensorineural deafness. More recently, mutations in Cx43 were also demonstrated to be responsible for a recessive form of non-syndromic deafness (Liu et al. 2001). Although recent discoveries have led to a better understanding of the role of gap junctions in auditory function, many important questions about gap junctions in the cochlea remain unanswered: why is more than one connexin present in the cochlea; do different connexins perform different functions; by what molecular mechanisms do mutations of connexin genes cause deafness? To answer these questions, an accurate knowledge of the expression and localization of different connexins in the cochlea appears to be essential.
The purpose of this study was to determine whether the two different connexins, Cx43 and Cx26, are expressed and localized in a similar manner in the cochlea. For this purpose, we performed three-dimensional (3-D) analysis on the expression and localization of Cx43 and Cx26 in the adult rat cochlea using two types of prepared samples, i.e., cryosections and surface preparations, in combination with confocal microscopy. The results indicated that the expression and localization of Cx43 differed from those of Cx26. Therefore, we further investigated whether this difference was associated with the expression and localization of zonula occludens-1 (ZO-1), because the carboxyl terminals of Cx43 have been shown to be bound with ZO-1 (Giepmans and Moolenaar 1998; Toyofuku et al. 1998), whereas those of Cx26 lack the sequences that potentially interact with ZO-1 (Nielsen et al. 2001). To conduct this investigation, we carried out double immunolabeling for ZO-1 and either Cx43 or Cx26 in the cochlear lateral wall.
Materials and Methods
Animals
A total of 20 adult male Wistar rats free of middle ear infection were used. The animals were perfused transcardially with freshly prepared 2% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4, 4C) under deep pentobarbital anesthesia, and the temporal bones were removed bilaterally. The cochlear lateral walls, spiral limbus, and the organ of Corti were dissected out under a dissection microscope and placed in the same fixative at 4C for 1 hr. Two types of sample preparations, i.e., cryosections and surface preparations, were made for immunolabeling. For cryosections of the cochlear lateral wall and spiral limbus, the specimens were infused with Tissue-Tek OCT compound (Miles; Elkhart, IN), frozen in disposable vinyl specimen molds (Tissue-Tek Cryomold; Miles) by partial immersion in liquid nitrogen, and stored at −80C. Serial 14-μm frozen sections were cut with a cryostat microtome (CRYOCUT 1800; Reichert-Jung, Heidelberg, Germany) and placed on silane-coated slides (Matsunami; Osaka, Japan). For surface preparations, the stria vascularis was gently separated from the spiral ligament under a dissection microscope and placed on silane-coated slides so that the marginal cell layer or the basal cell layer was in contact with the surface of the slides. Surface preparations of the organ of Corti were placed on the slides so that the side of the basilar membrane was in contact with the surface of the slides. All animal experiments described in this study adhered to the standards detailed in “Principles of Laboratory Animal Care” (NIH publication no. 85 23, revised 1985) and were approved by the Committee for Animal Research, Kyoto Prefectural University of Medicine.
Immunohistochemistry
The specimens on the slides were air-dried and fixed in absolute ethanol at −20C for 15 min. After incubation in PBS with 5% skim milk for 15 min at room temperature to block nonspecific labeling, primary antibodies, including a mouse monoclonal antibody against Cx26 (1:200 dilution; Zymed Laboratories, South San Francisco, CA), a rabbit polyclonal antibody against a Cx26-specific peptide (amino acid residues 101–119; 1:2000 dilution) (Kuraoka et al. 1993), a rabbit polyclonal antibody against Cx26 (1:200 dilution; Zymed), a mouse monoclonal antibody against Cx43 (1:500 dilution; Chemicon, Temecula, CA), a rabbit antibody against Cx43 (1:500 dilution; Zymed), and a rabbit polyclonal antibody against ZO-1 (1:200 dilution; Zymed), were applied overnight at 4C. After three 5-min washes with PBS, incubation with Alexa 488-conjugated goat anti-rabbit or goat anti-mouse IgG (1:500 dilution; Molecular Probes, Eugene, OR) was performed for 1 hr at 37C. After the final rinses in PBS, sections were mounted in Vectorshield (Vector Laboratories; Burlingame, CA). All antibody dilutions were made in PBS containing 1% bovine serum albumin (Sigma; St Louis, MO) and 0.1% sodium azide with 0.1% Triton X-100. Some sections were double-labeled with the combined pairs Cx26 and Cx43, Cx26 and ZO-1, or Cx43 and ZO-1 using Alexa 488-conjugated and Alexa 594-conjugated secondary antibodies (1:500 dilution; Molecular Probes). On some slides, Alexa 488-conjugated or Texas Red-X-conjugated phalloidin (1:200 dilution; Molecular Probes) was used to visualize filamentous actin (F-actin). As negative controls, non-immune normal rabbit (Zymed) and mouse (Zymed) sera were used in place of the first antibodies under otherwise similar conditions for immunofluorescence.
Confocal Laser Scanning Microscopy
The immunostained slides of cryosections and surface preparations were observed with a confocal laser scanning microscope (Olympus FLUOVIEW system; Olympus, Tokyo, Japan) equipped with an oil-immersion objective (Plan Apo × 60, NA = 1.4; Olympus). An argon–krypton laser produced excitation bands at 488 nm for Alexa 488 and 568 nm for Alexa 594 and Texas Red-X. Fluorescence images were collected with emission filters for 510–550 nm for Alexa 488 and 585–610 nm for Alexa 594 and Texas Red-X. Simultaneous images (800 × 600 pixels, 12 bits each) of the Alexa 488 label and either Alexa 594 or Texas Red-X were acquired and stored. Three-dimensional analysis was done by acquiring a 2-D image at each of more than 10 adjacent focus planes that were 1, 1.5, or 2 μm apart within the specimen by driving a focus motor. This was ultimately viewed as a stereo pair made by projecting the data along two axes about 10° apart. Digital images were transferred to a Macintosh computer (Apple Computer; Cuperino, CA) equipped with the Photoshop Program (Adobe Systems; Tucson, AZ) and printed on an ink-jet printer (PM-3500C; Epson, Tokyo, Japan). A total of 40 cochleas were analyzed.
Results
Cx43 and Cx26 in Cochlear Lateral Wall
The localization of Cx43 was notably different from that of Cx26 in the cochlear lateral wall of adult rats. In cryostat sections under low magnification, immunolabeling for Cx43 using the rabbit anti-Cx43 antibody (Zymed) showed that Cx43-immunolabeled spots were localized mainly in the stria vascularis, but a few spots were also observed on the lateral side of the spiral ligament (Figures 1A and 1C). In contrast, Cx26 immunolabeling using the mouse monoclonal anti-Cx26 antibody (Zymed) was detected not only in the stria vascularis but also throughout the spiral ligament (Figures 1B and 1C), consistent with the result of our previous study (Suzuki et al. 2001).
To ascertain the precise localization of Cx43 and Cx26 in the stria vascularis, we performed double labeling using rabbit anti-Cx43 (Zymed) or rabbit anti-Cx26 antibody (Kuraoka et al. 1993) and F-actin using Texas Red-X-labeled phalloidin. The apical surface of the marginal cells and the basal cell layer were stained with Texas Red-X-labeled phalloidin (Figures 1D and 1E). Most Cx43-positive spots were localized between the Texas Red-X-phalloidin-positive marginal cells and the basal cells, indicating preferential localization of Cx43 within the stria vascularis (Figure 1D). On the other hand, Cx26-positive spots were distributed not only within the stria vascularis but also in the spiral ligament (Figure 1E).
To examine the relationship between Cx43 and Cx26 in the cochlear lateral wall, we carried out double immunolabeling using rabbit anti-Cx43 (Zymed) and mouse anti-Cx26 (Zymed) antibodies in the cryostat sections and 3-D analysis using serial confocal images. Stereo pairs reconstructed from serial confocal images demonstrated that Cx43 was localized within the stria vascularis, whereas Cx26 was present in the stria vascularis and spiral ligament (Figure 1F). In addition, yellow spots were found in the stria vascularis, indicating co-localization of Cx43 and Cx26.
To further investigate the co-localization of Cx43 and Cx26 in the stria vascularis, we carried out 3-D analysis of double immunolabeling using the rabbit anti-Cx43 (Zymed) and mouse anti-Cx26 (Zymed) antibodies and surface preparations, in which the surfaces of the marginal cells were viewed from the side facing the endolymphatic space. Serial confocal images, in which the planes were 1 μm apart, showed that a substantial proportion of the immunolabeled spots were yellow, confirming co-localization of Cx43 and Cx26 (Figure 2). However, many immunolabeled spots were not yellow, but were either green or red, indicating that separate localization of Cx43 and Cx26 predominated over the co-localization, especially on the basal side.
Identical results for the localization of Cx43 and Cx26 in the cochlear lateral wall were obtained with different antibodies against Cx43 and Cx26, including mouse anti-Cx43 (Chemicon) and rabbit anti-Cx26 (Kuraoka et al. 1993) antibodies, in different combinations (data not shown). With non-immune rabbit and mouse sera, nonspecific fluorescence was barely detected in the adult rat cochlear lateral wall (Figures 1G and 2M).
Cx43 and Cx26 in the Spiral Limbus
Differences between the localization of Cx43 and that of Cx26 were also found in the spiral limbus. Using the rabbit anti-Cx43 and mouse anti-Cx26 antibodies, we found that Cx43-positive spots were widely distributed in the spiral limbus (Figure 3A). On the other hand, Cx26-positive spots were more concentrated on the side facing the scala vestibuli and in the basal portion of the spiral limbus, although some Cx26-positive spots were detected in the apical portion of the spiral limbus (Figure 3B). Merged images showed that Cx43 and Cx26 were sometimes co-localized in the spiral limbus (Figure 3C). With non-immune rabbit and mouse sera, nonspecific fluorescence was barely detected in the spiral limbus (Figure 3D).
Cx43 and Cx26 in the Organ of Corti
Cx43-positive spots were clearly detected between several types of supporting cells. Although precise identification of the supporting cells was not possible in this study, Cx43-positive cells appeared to include at least the inner and outer phalangeal cells and Hensen's cells. The Cx43-positive spots were fewer and smaller than the Cx26-positive spots. Double labeling using Alexa 488-conjugated phalloidin and the mouse anti-Cx26 antibody revealed that inner and outer pillar cells, as well as the cell boundaries between outer hair cells and outer phalangeal cells, were strongly stained with Alexa 488-conjugated phalloidin and that many Cx26-positive spots were localized among supporting cells (Figure 4A). Stereo paired images of immunolabeling using the rabbit anti-Cx43 and mouse anti-Cx26 antibodies showed that Cx43-positive spots were distributed adjacent to more abundant Cx26-positive spots and that some Cx43-positive spots were co-localized with Cx26-positive spots (Figures 4B and 4C). With non-immune rabbit and mouse sera, nonspecific fluorescence was barely detected in the organ of Corti (Figure 4D).
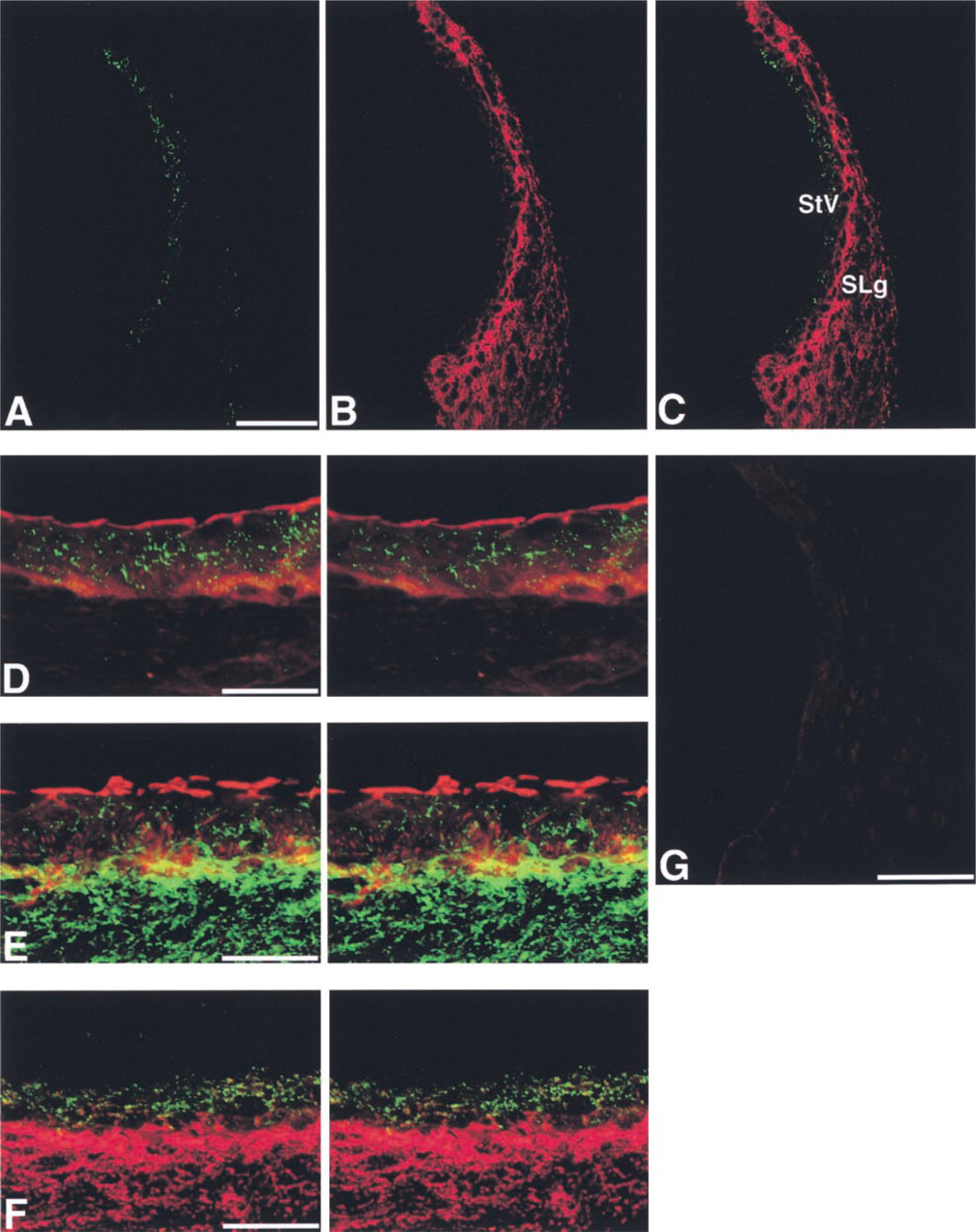
Immunohistochemical localization of Cx43 and Cx26 in the cochlear lateral wall. (
ZO-1, Cx43, and Cx26 in the Cochlear Lateral Wall
ZO-1 has been shown to be bound with the carboxyl terminals of Cx43 (Giepmans and Moolenaar 1998; Toyofuku et al. 1998) and Cx45 (Laing et al. 2001), and it is suggested to play a role in determining the localization of several connexins, including Cx43. In contrast, the carboxyl terminal sequence of Cx26 lacks the final three residues (V/L)-X-(V/I) that potentially interact with ZO-1 (Nielsen et al. 2001). To assess whether the difference in localization between Cx43 and Cx26 is associated with the localization of ZO-1, i.e., whether the localization of ZO-1 accords with that of Cx43 more closely than with that of Cx26, we carried out double immunolabeling for ZO-1 and either Cx43 or Cx26 in the cochlear lateral wall. In the cryosections, immunolabeling using rabbit anti-ZO-1 (Zymed) and mouse anti-Cx43 (Chemicon) antibodies showed that most Cx43-positive spots were present in the stria vascularis and that only a small ZO-1-immunolabeled area overlapped with Cx43-positive spots, although some Cx43-positive spots were co-localized with ZO-1 immunolabeling in the basal cell layer (Figures 5A and 5C). Conversely, immunolabeling using rabbit anti-ZO-1 (Zymed) and mouse anti-Cx26 (Zymed) antibodies demonstrated that a fairly large ZO-1-immunolabeled area overlapped with Cx26-positive spots in the basal cell layer of the stria vascularis and spiral ligament (Figures 5B and 5D).
Discussion
In this study we provide evidence that the expression and localization of Cx43 are different from those of Cx26 in the adult rat cochlea, although there is some overlap between them, and that Cx43 and Cx26 are occasionally co-localized. The difference was demonstrated in three distinct regions of the cochlea, i.e., the cochlear lateral wall, spiral limbus, and organ of Corti. In the cochlear lateral wall, we found that Cx43-positive spots were localized mainly in the stria vascularis, but a few spots were present in the spiral ligament, whereas Cx26-positive spots were detected in both the stria vascularis and spiral ligament. In the spiral limbus, Cx43-positive spots were widely distributed, whereas Cx26-positive spots were more concentrated on the side facing the scala vestibuli and in the basal portion of the spiral limbus. In the organ of Corti, Cx43-positive spots were present between the supporting cells (the inner and outer phalangeal cells and Hensen's cells), similarly to Cx26-positive spots. However, the Cx43-positive spots in the supporting cells of the organ of Corti were much smaller and fewer than the Cx26-positive spots.
Our results also indicate that the differences between the expression and localization of Cx43 and those of Cx26 in the cochlea are not associated with their difference in structure, i.e., the carboxyl terminals of Cx43 possess sequences for binding to ZO-1, whereas those of Cx26 do not. Our results for double immunolabeling for ZO-1 and either Cx43 and Cx26 in the cochlear lateral wall demonstrated only a small area of overlap of ZO-1-immunolabeling with Cx43-positive spots, whereas there was a fairly large area of overlap with Cx26-positive spots in the basal cell layer of the stria vascularis and the spiral ligament, in discord with our hypothesis that the localization of ZO-1 accords with that of Cx43 more closely than with that of Cx26. In turn, the overlap of ZO-1 and Cx26 does not necessary imply functional association of these proteins because the abundance of the two proteins would also increase the probability of their overlap. It therefore remains to be determined whether the overlap of ZO-1 and Cx26 has any physiological or functional significance in the cochlear lateral wall.
The present study elucidated Cx43 localization in the rat cochlea. We demonstrated here that Cx43 was localized in the stria vascularis, spiral limbus, and supporting cells in the organ of Corti. Formerly, no reliable information was available about the expression and localization of Cx43 in the cochlea. Liu et al. (2001) noted that the issue of whether Cx43 is present in a highly restricted group of supporting cells or throughout non-sensory epithelial cells and in type I fibrocytes would have to be resolved, because these authors found that the pattern of staining for Cx43 was dependent on tissue fixation conditions in their study. Similarly, Lautermann et al. (1998) detected only very weak staining for Cx43 in the stria vascularis and the spiral ligament, and between supporting cells of the organ of Corti, and they failed to find Cx43 in the spiral limbus. In contrast, the use of more than one antibody for each connexin and different preparation makes our present study the most thorough and compelling one about the distribution of Cx43 in the cochlea. Concerning Cx43 and Cx26, identical results of almost equal quality were obtained with different antibodies, including mouse monoclonal and rabbit polyclonal antibodies, and with samples prepared with different fixatives, such as ethanol/acetic acid (data not shown). Furthermore, the use of confocal laser scanning microscopy and two different sample preparation methods enabled us to acquire simultaneous high-resolution stereo paired images of immunolabeling for the two different connexins, or for one connexin combined with either ZO-1 or F-actin.
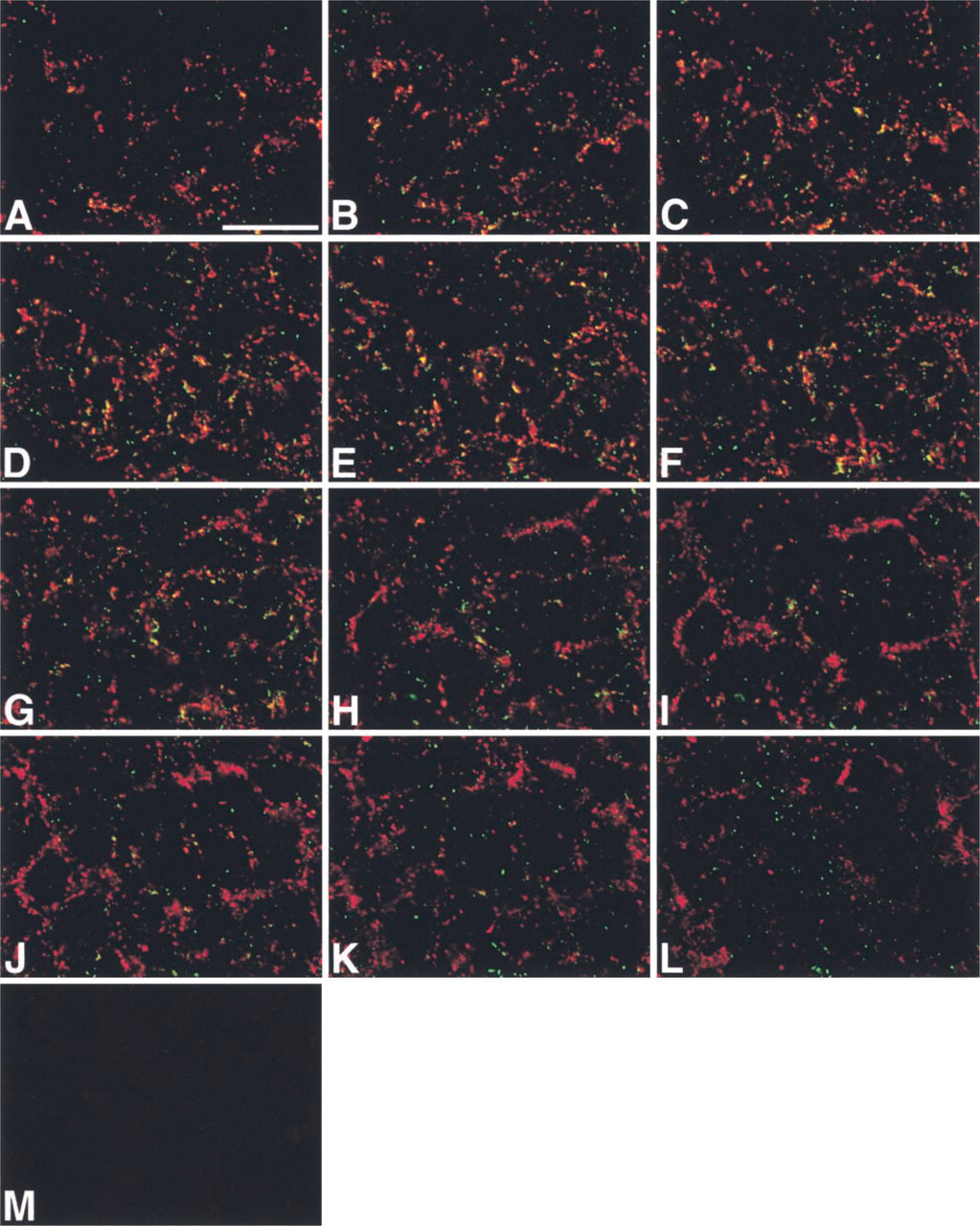
IHC localization of Cx43 (green) and Cx26 (red) in the stria vascularis using a surface preparation. (
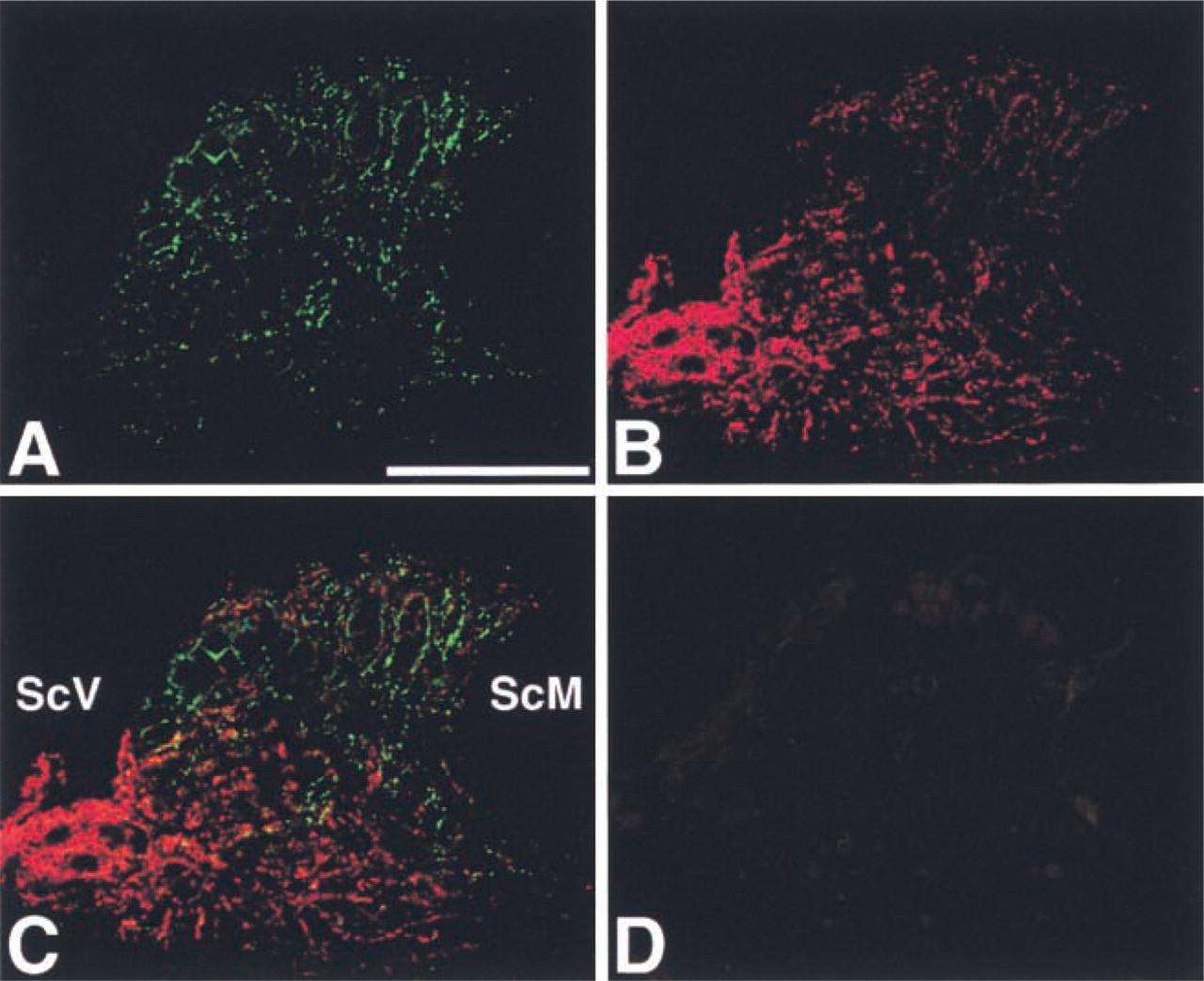
IHC localization of Cx43 (green) and Cx26 (red) in the spiral limbus. (
Our finding that the expression and localization of Cx43 differ from those of Cx26 in the adult rat cochlea indicates that gap junctions in the different regions of the cochlea are composed of multiple connexins in different combinations and/or proportions. The diversity of connexins in the cochlea that we found here is consistent with that reported by Xia et al. (2000), who found that the distributions of Cx31 and Cx26 were obviously different in the developing mouse cochlea. This diversity of connexins in the cochlea offers several important clues that might elucidate the role of gap junctions in auditory function under physiological and pathological conditions. For example, the region-specific distribution of different connexins implies differences in the function and regulation of the gap junctional intercellular channels in the cochlea. Distinct electrophysiological and ion-selective properties have been shown for homotypic and heterotypic gap junctions containing different connexins (Verselis and Veenstra 2000). Furthermore, there are phosphorylation sites in Cx43 but not in Cx26, indicating a difference in regulation between gap junctions containing Cx43 and those containing Cx26. Our finding that gap junctions in the different regions of the cochlea contain multiple connexins in different combinations and/or proportions suggests that the low molecular weight molecules that pass through gap junctions and play important roles in the auditory function vary with different regions in the cochlea.
A recent study on mutant mice with hearing loss caused by target ablation of Cx26 in the cochlear epithelial gap junction network has shown that cell death initially affected only the supporting cells of the inner hair cells (Cohen-Salmon et al. 2002). Combined with our present finding that the number and size of Cx43-positive spots in the supporting cells of the organ of Corti are much smaller than those of Cx26, this suggests that the gap junctional intercellular communication mediated by Cx43, which is present in such a small amount in the organ of Corti, is not sufficient to support cell survival in mutant mice and that Cx26, which is expressed in larger amounts, is essential for cell survival. Alternatively, the changes in the properties of gap junctional channels caused by target ablation of Cx26 might be more closely related to supporting cell death than is the large loss of total connexins in the organ of Corti caused by Cx26 ablation.
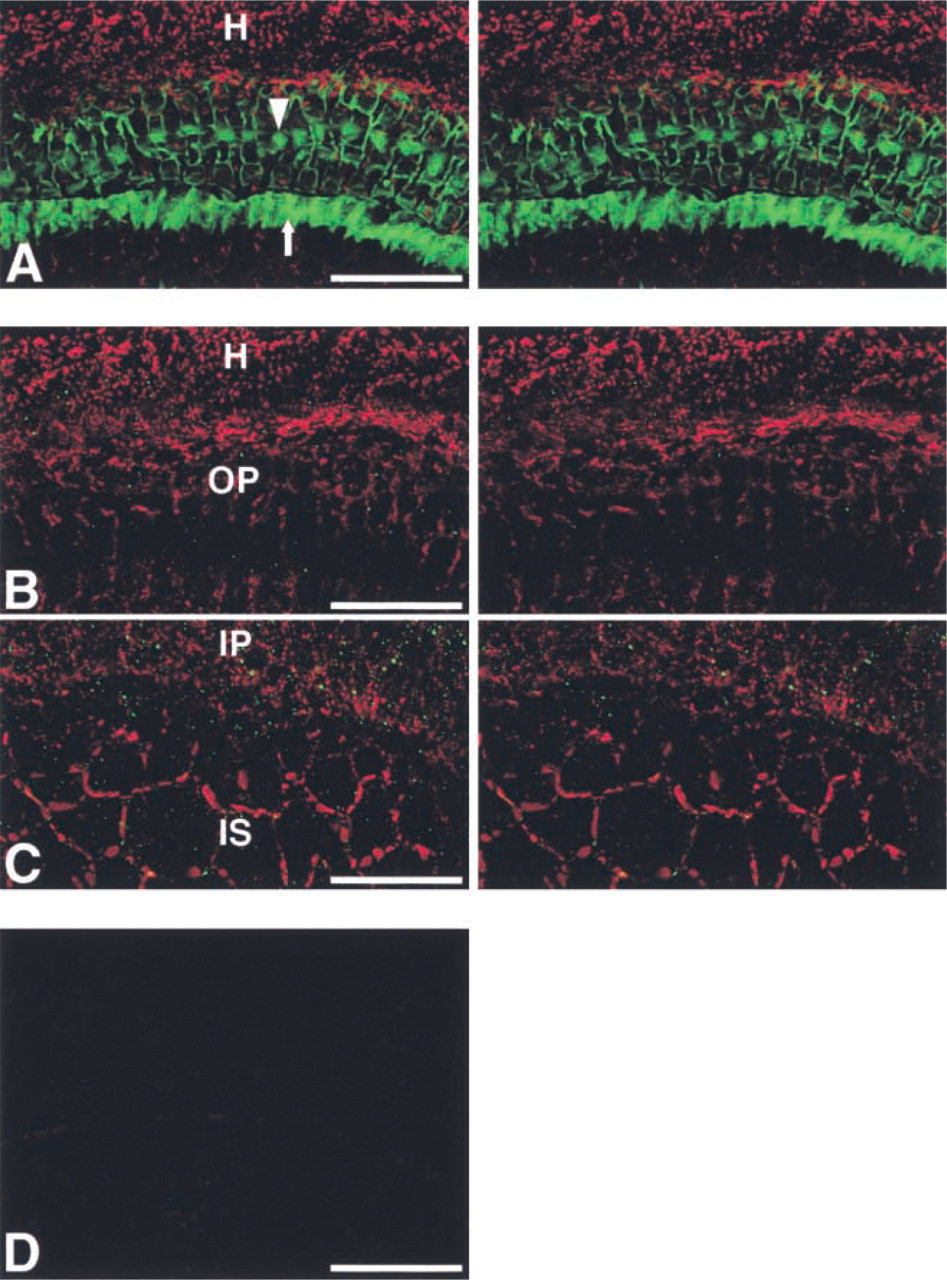
IHC localization of Cx43 and Cx26 in the organ of Corti using a surface preparation. (
The difference in expression and localization between Cx43 and Cx26 also suggests that the target cells for hearing loss caused by mutations of different connexins would not be the same. On the assumption that the expression and localization of Cx43 in the rat are similar to those in the human cochlea, the small number and size of Cx43-positive spots in the organ of Corti, combined with the substantial expression of Cx43 in the stria vascularis and spiral limbus, might indicate that the target cells for human non-syndromic deafness associated with mutations in Cx43 (Liu et al. 2001) are not supporting cells of the organ of Corti. Target ablation of Cx43 in the cochlear epithelial gap junction network, similar to that already carried out for Cx26 (Cohen-Salmon et al. 2002), will surely give us valuable information on this matter.
Our present study revealed that Cx43 and Cx26 are occasionally co-localized in the cochlea, especially in the stria vascularis. Rouan et al. (2001) reported that Cx43 and Cx26 were focally co-localized in suprabasal epidermal keratinocytes in carriers of hearing impairment and palmoplantar keratoderma with a dominant Cx26 mutation (ΔE42). They also demonstrated that the mutant Cx26 significantly inhibits not only intercellular conductance of co-expressed wild-type Cx26 but also that of wild-type Cx43, i.e., transdominant inhibition of Cx43 by the mutant Cx26. Our data showing co-localization of Cx43 and Cx26 in the stria vascularis suggest that such a trans-dominant inhibition of Cx43 by mutant Cx26 also occurs in the stria vascularis and plays some role in the pathogenesis of hearing loss. On the other hand, in the organ of Corti, it appears that most Cx43 and Cx26 is not incorporated into the same gap junctions, although some Cx43-positive spots were co-localized with Cx26-positive spots. This is because the number, size, and shape of the immunolabeled puncta were dramatically different between Cx43 and Cx26, i.e., Cx43 immmunolabeling showed a small number of tiny, round, particulate, and separate structures, whereas Cx26 immunolabeling demonstrated a great number of large and extended structures.
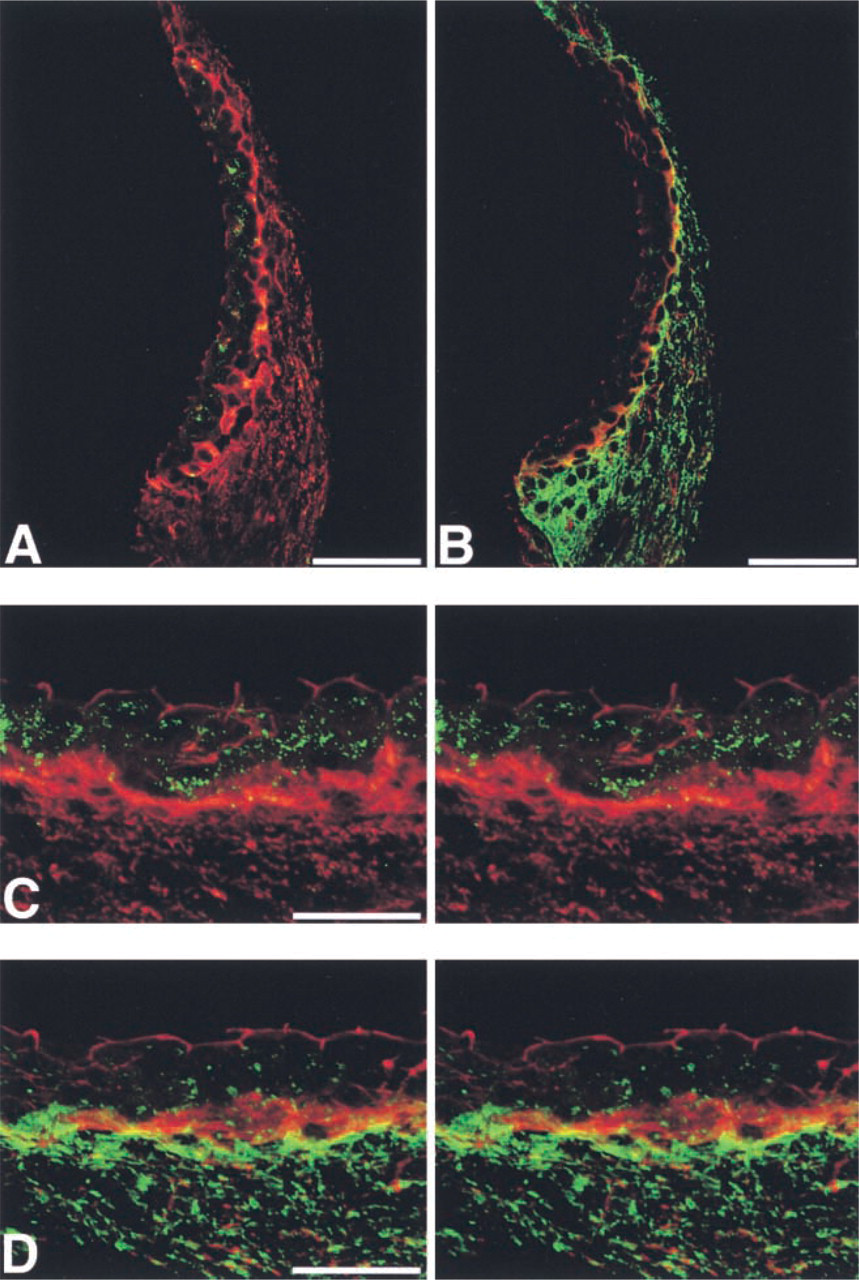
IHC localization of ZO-1, Cx43, and Cx26 in the cochlear lateral wall. (
In conclusion, we successfully demonstrated the expression and localization of Cx43 in the adult rat cochlea using confocal microscopy and two types of sample preparations. The expression and localization of Cx43 differed from those of Cx26 in the adult rat cochlea, although there was some overlap between them and co-localization of Cx43 and Cx26 was occasionally found.
Footnotes
Acknowledgements
Supported by grants-in-aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology, Japan.
We gratefully acknowledge the generous gift of the polyclonal anti-connexin26 antibody from Prof Y. Shibata (Kyushu University School of Medicine; Fukuoka, Japan).
We are also grateful to Dr Y. Oyamada for helpful discussion and critical reading and to Ms Ramona Ratliff for advice on English usage.