
Abstract
Aminopeptidase A (APA) is one of the many renal hydrolases. In mouse kidney, APA is predominantly expressed on the brush borders and sparsely on the basolateral membranes of proximal tubular epithelial cells. However, when large amounts of monoclonal antibodies (MAbs) against APA were injected into mice, we observed strong binding of the MAbs to the basolateral membranes, whereas the MAbs bound only transiently to the brush borders of the proximal tubular epithelial cells. In parallel, APA itself disappeared from the brush borders by both endocytosis and shedding, whereas it was increasingly expressed on the basolateral sides. Using ultrastructural immunohistology, we found no evidence for transcellular transport of endocytosed APA to the basolateral side of the proximal tubular epithelial cells. The absence of transcellular transport was confirmed by experiments in which we used a low dose of the MAbs. Such a low dose did not result in binding of the MAbs to the brush borders and had no effect on the presence of APA in the brush borders of the proximal tubular epithelial cells. In these experiments we still could observe binding of the MAbs to the basolateral membranes in parallel with the local appearance of APA. In addition, treatment of mice with chlorpromazine, a calmodulin antagonist that interferes with cytoskeletal function, largely inhibited the MAb-induced modulation of APA. Our studies suggest that injection of MAbs to APA specifically interrupts the normal intracellular traffic of this enzyme in proximal tubular epithelial cells. This intracellular transport is dependent on the action of cytoskeletal proteins.
Keywords
A
In a previous study we found that a single injection of a relatively large amount of a monoclonal rat anti-mouse APA antibody (MAb; ASD-4) into mice resulted in direct binding of the MAb to the podocytes. Unexpectedly, we also observed binding of the MAb to the basolateral sides of PTECs, a site at which APA is only sparsely expressed (Assmann et al. 1992). Concomitantly, the expression of APA at the basolateral side of PTECs increased, whereas a reduction occurred at the apical side of the PTECs. As indicated above, APA may be involved in regulation of the Ang II activity. Therefore, changes in the localization of APA may be relevant for Ang II physiology. The aim of the present study was to examine the mechanisms of this antibody-mediated modulation of APA on the PTECs.
Materials and Methods
Antibodies
Three MAbs against mouse APA (coded ASD-3, ASD-37, and ASD-41), and one MAb against mouse dipeptidyl peptidase IV (DPP IV; coded ASD-31) were raised by immunizing rats with the detergent phase of a Triton X-114-solubilized mouse brush border preparation as described (Assmann et al. 1992; Mentzel et al. 1996a,b). The characteristics of the anti-APA MAbs have been previously described in detail (Mentzel et al. 1996a,b). ASD-3 is an IgG1, whereas ASD-37 and ASD-41 are IgG2a. The anti-APA and anti-DPP IV MAbs were produced as ascites in BALB/c nu/nu mice and were purified by ammonium sulfate precipitation (Assmann et al. 1992). A polyclonal rabbit anti-mouse glomerular basement membrane (GBM) antibody was produced as previously described (Assmann et al. 1985).
Animal Experiments
In a previous study we observed that a single IV injection of an MAb against APA (ASD-4) induced a reallocation of APA from the apical brush border to the basolateral side of PTECs (Assmann et al. 1992). To determine whether this so-called antigenic modulation, i.e., the removal of APA from the brush borders and the appearance of APA at the basolateral sides of PTECs, was specific for only one MAb, we examined the effects of three other anti-APA MAbs. BALB/c mice were injected IV with a relatively large amount of the anti-APA MAbs (8 mg). In control experiments, BALB/c mice were injected IV with 8 mg of the anti-DPP IV MAb and C57Bl6 mice were injected IV with 20 mg of the polyclonal rabbit anti-mouse GBM antibody. To examine the effect of a low dose (1 mg) of anti-APA antibodies, we selected one anti-APA MAb, i.e., ASD-41, which had a strong modulating effect on APA in the PTECs at a dose of 8 mg. To study the role of the cytoskeleton mice (n = 5) were treated with 5 mg chlorpromazine/100 g body weight every 12 hr by IP injection. A control group received saline in the same volume. To ensure its effectiveness, this treatment was started 1 day before the injection of 8 mg ASD-41 and was continued thereafter. At Day 1 after injection of the antibodies, mice were sacrificed by cervical dislocation and their kidneys processed for immunofluorescence (IF), enzyme histochemistry and, in some cases, for immunoelectron microscopy (IEM).
Immunofluorescence and Immunoelectron Microscopy
The binding of the injected anti-APA and anti-DPP IV MAbs was examined by direct IF with a fluorescein isothiocyanate (FITC)-labeled rabbit anti-rat IgG (DAKO, Copenhagen, Denmark), as described before (Mentzel et al. 1996a,b). The binding of the injected polyclonal rabbit anti-mouse GBM antibodies was also visualized by a direct IF with an FITC-labeled swine anti-rabbit IgG (DAKO; Copenhagen, Denmark). The localization of APA was examined by an indirect IF with a rat anti-mouse APA MAb of another suclass (ASD-38; IgG2b subclass) whose binding was not blocked by the injected MAb (Mentzel et al. 1999). The binding of the anti-APA MAb was then visualized by using a subclass-specific, FITC-labeled sheep anti-rat IgG (Serotec; Oxford, UK). For detection of the DPP IV antigen, similar methods were used. All sections were examined in a fluorescence microscope equiped with Ploemopak epiillumination (Leitz Wetzlar; Wetzlar, Germany) and the staining intensity was recorded semiquantitatively on a scale from 0 to 4+ as described previously (Assmann et al. 1983).
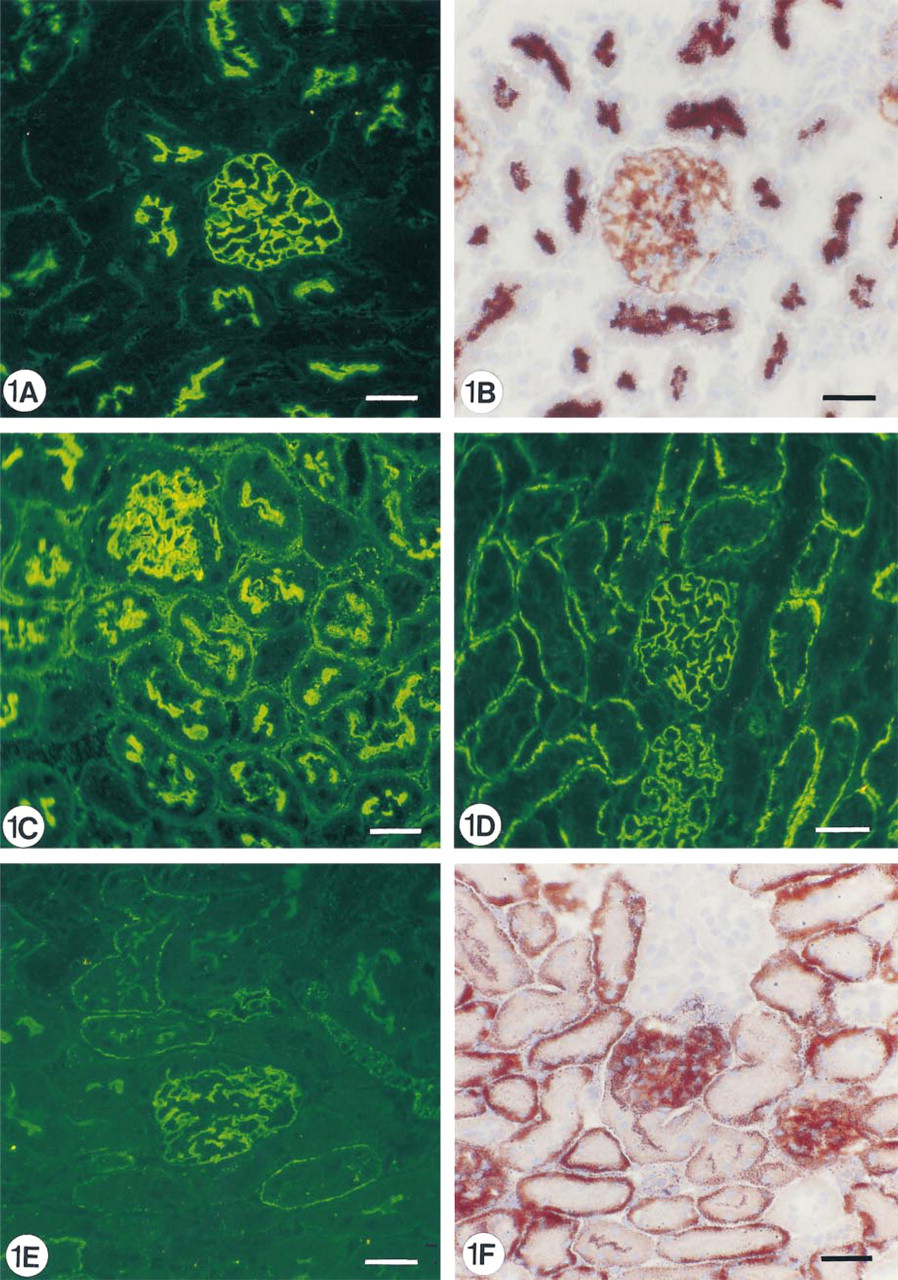
Immunofluorescence and enzyme histochemistry of a normal mouse kidney showing the location of aminopeptidase A (
The exact binding of the injected MAbs was in some selected cases also studied by IEM with a peroxidase-labeled rabbit anti-rat IgG (Harlan Seralab; Blackthorn, Bicester, UK) absorbed with 2% normal mouse serum and visualized with diaminobenzidine as described previously (Assmann et al. 1992). The expression of APA was studied by indirect IEM using a rat anti-mouse APA MAb as described above. This antibody was then detected by a peroxidase-labeled rabbit anti-rat IgG as described previously (Assmann et al. 1992; Mentzel et al. 1996a,b). In addition, the binding of the injected MAbs was studied by incubating 20-μm-thick frozen sections for 18 hr at 4C with 1-nm gold-labeled goat anti-rat IgG Fab fragments (Nanoprobes; Stony Brook, NY) absorbed with 2% normal mouse serum. After rinsing in PBS, the sections were postfixed in 1% glutaraldehyde for 10 min, rinsed in distilled water, and silver-enhanced for 10 min at room temperature (RT) with HQ silver, according to the instructions of the manufacturer. The sections were rinsed in distilled water, postfixed in cacodylate-buffered 1% OsO4 for 30 min at RT, dehydrated, and embedded in Epon 812. Ultrathin sections were prepared on an ultratome (LKB Instruments; Bromma, Sweden), counterstained with 3% uranyl acetate, and examined in an electron microscope (JEOL; Tokyo, Japan) as described (Assmann et al. 1992; Mentzel et al. 1996a,b).
Enzyme Histochemistry
The enzyme activity of APA in the kidneys was visualized by enzyme histochemistry according to Lojda and Gossrau as described before (Lojda and Gossrau 1980; Mentzel et al. 1996a,b).
Results
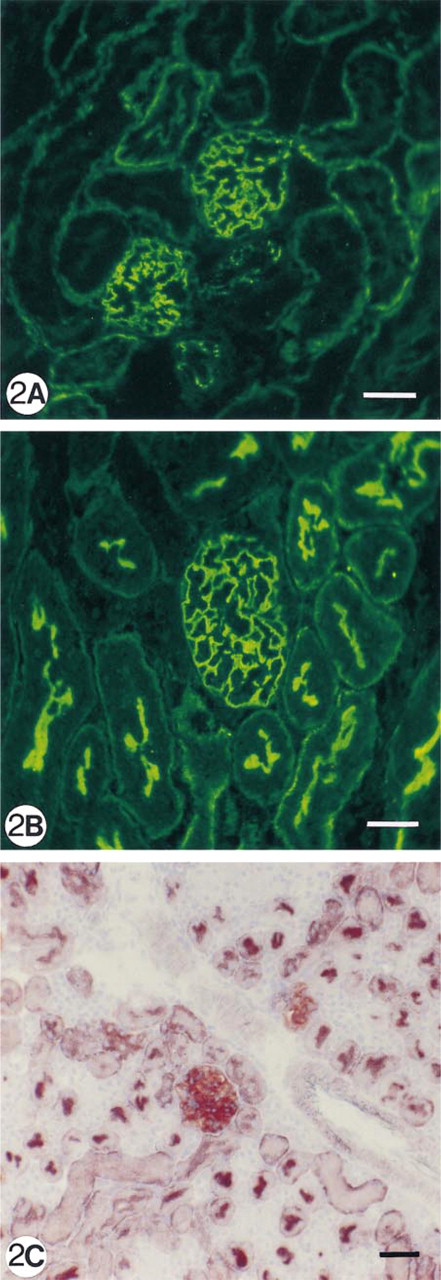
Immunofluorescence (
As can be demonstrated by IF and enzyme histochemistry, the APA molecule and its enzyme activity are mainly localized on podocytes and brush borders of PTECs, but not or sparsely on the basolateral invaginations of these proximal cells (Figures 1A and 1B). After injection of 8 mg of anti-APA MAbs, all MAbs bound extensively to the brush borders of PTECs within the first hours after injection (Figure 1C. After 1 day, we observed that all three MAbs were bound to approximately the same extent to the basolateral sides of the PTECs, and not or only weakly and most often focally to the brush borders (Figure 1D. As seen by IF and enzyme histochemistry, APA expression was also predominantly observed at the basolateral sides of PTECs, and only in brush borders of some proximal tubules (Figures 1E and 1F). Injection of a rat anti-mouse DPP IV MAb (8 mg) or rabbit anti-mouse GBM antiserum (20 mg) did not affect the APA expression in the mouse kidneys (not shown). DPP IV was redistributed from the apical to the basolateral sides of the PTECs on injection of the rat anti-mouse DPP IV MAbs, as reported earlier by others (Yamaguchi et al. 1996). The appearance of APA at the basolateral side of PTECs and the removal of APA from the brush borders suggested that this might be caused by transcellular transport of apical anti-APA MAb/APA immune complexes to the basolateral side of PTECs. Therefore, we also examined the effects of a low dose (1 mg) of one of the anti-APA MAbs (ASD-41), on the basis of the assumption that the injection of such a low amount of MAb should not gain access to the lumina of PTECs. Indeed, after injection of 1 mg ASD-41 we did not observe binding of the MAb to the brush border of the PTECs, and the expression of APA at this side remained unchanged. Still, the antibodies were bound to the basolateral sides of PTECs (Figure 2A. In addition, APA expression was present at the basolateral sides of PTECs (Figures 2B and 2C). The above-mentioned changes in the localization of APA were confirmed by indirect IEM (Figures 3A-3C). By IEM we observed shedding of the APA/anti-APA complexes (Figures 4A and 4B). In addition, endocytotic vesicles containing APA or anti-APA MAb were predominantly seen in the upper part of the PTECs (Figures 5A and 5B). We never observed fusion of vesicles with the basolateral membranes and therefore have found no firm evidence for transcellular vesicle transport of APA. Together, these data strongly suggest that the modulation of APA in the PTECs is not a consequence of transcellular transport of immune complexes formed at the brush borders.
The Role of the Cytoskeleton in APA Modulation in the PTECs
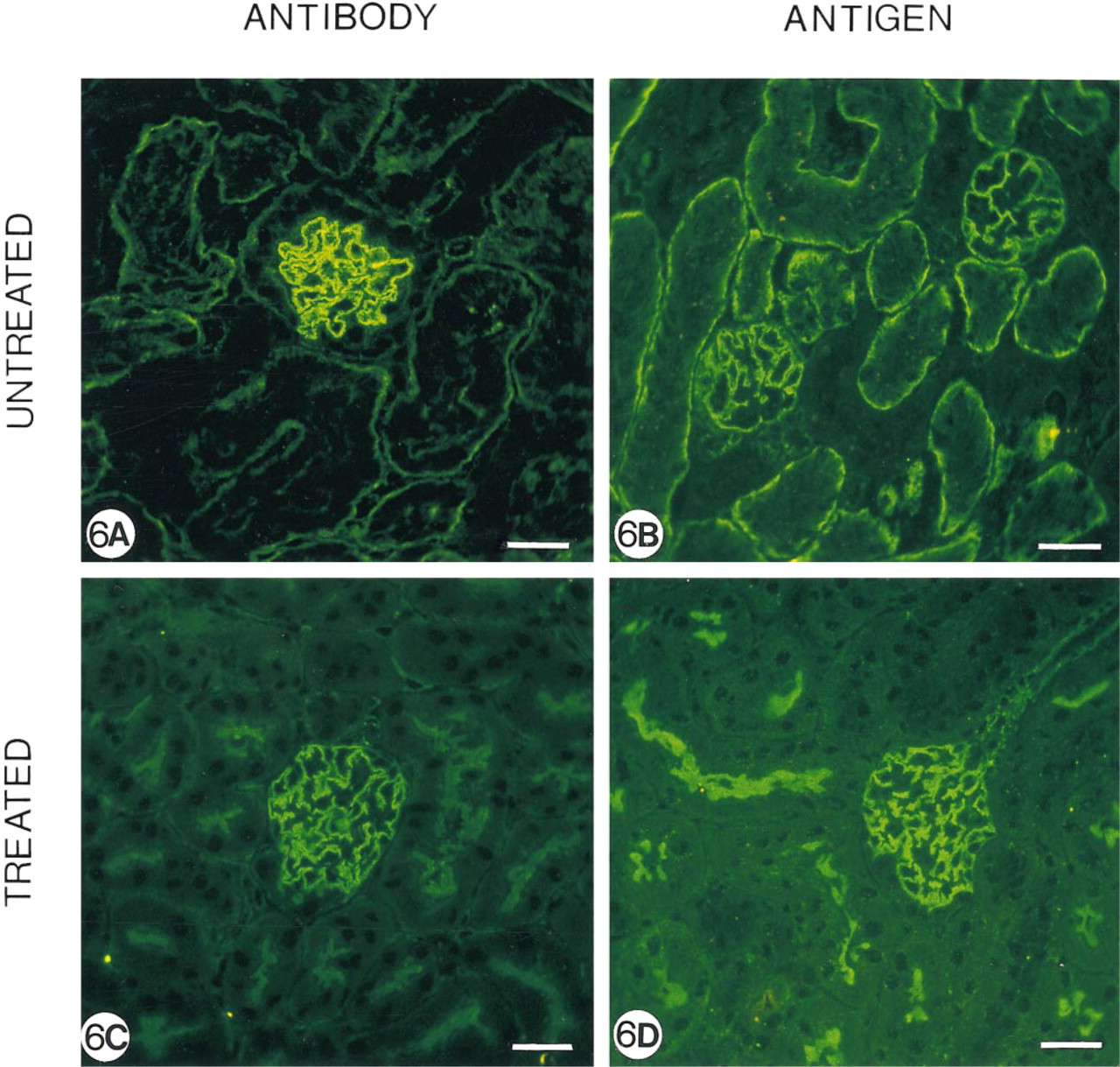
Because we have previously demonstrated that APA in mouse PTEC is covalently coupled to the γ-isoform of actin (Mentzel et al. 1994), we wondered what role the cytoskeleton plays in this anti-APA MAb-induced APA modulation. Therefore, mice injected with 8 mg ASD-41 were treated with the calmodulin antagonist chlorpromazine. As expected, in untreated control animals we observed binding of the MAb and expression of APA, particularly at the basolateral side of PTECs (Figures 6A and 6B). In contrast, mice treated with chlorpromazine demonstrated prominent binding of the MAb and APA expression at the brush borders (Figures 6C and 6D), and only minor localization of the injected MAb and APA expression at the basolateral side. These data suggest that the expression of APA at the basolateral sides and brush borders depends on cytoskeletal activity.
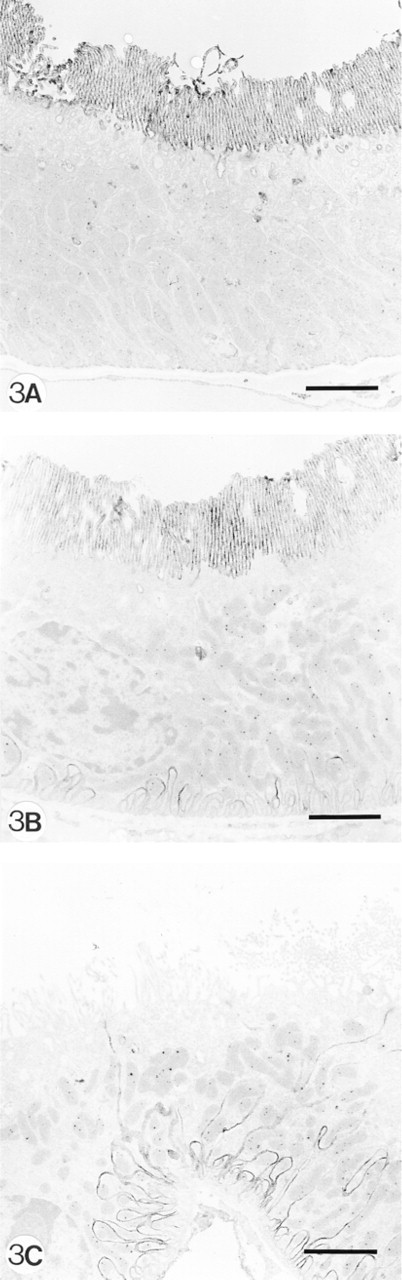
Indirect immunoelectron microscopy (immunoperoxidase technique) showing aminopeptidase A expression in PTECs of a normal mouse (
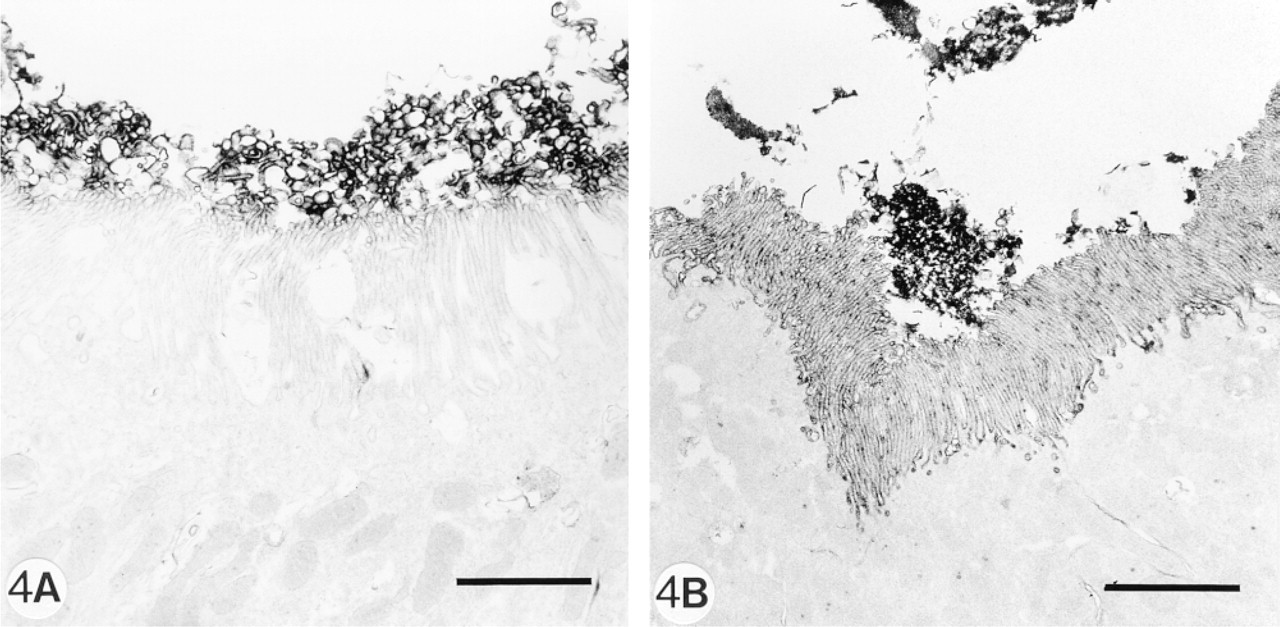
Immunoelectron microscopy staining (immunoperoxidase technique) demonstrating the presence of MAb ASD-41 (
Discussion
In this study we demonstrate that injection of anti-APA MAbs alters the localization of APA on mouse PTECs. Only anti-APA MAbs were able to cause this redistribution of APA expression. Therefore, the modulation is dependent on the interaction of an antibody with its specific antigen. However, because all anti-APA MAbs that we have used had a modulating effect, this modulation appears to be independent of the particular characteristics of the MAbs, such as the binding domain, the ability to block the enzyme activity, or the capacity to induce albuminuria (Assmann et al. 1992; Mentzel et al. 1996a,b). Such an antibody-induced modulation of an antigen is not unique for APA, because it has also been described for other renal tubular proteins such as megalin (Heymann antigen), DPP IV, and angiotensin-converting enzyme (Noble et al. 1981; Camussi et al. 1985; Fukatsu et al. 1988; Yamaguchi et al. 1996). The exact mechanisms of antigenic modulation in the PTEC are not very well understood at present.
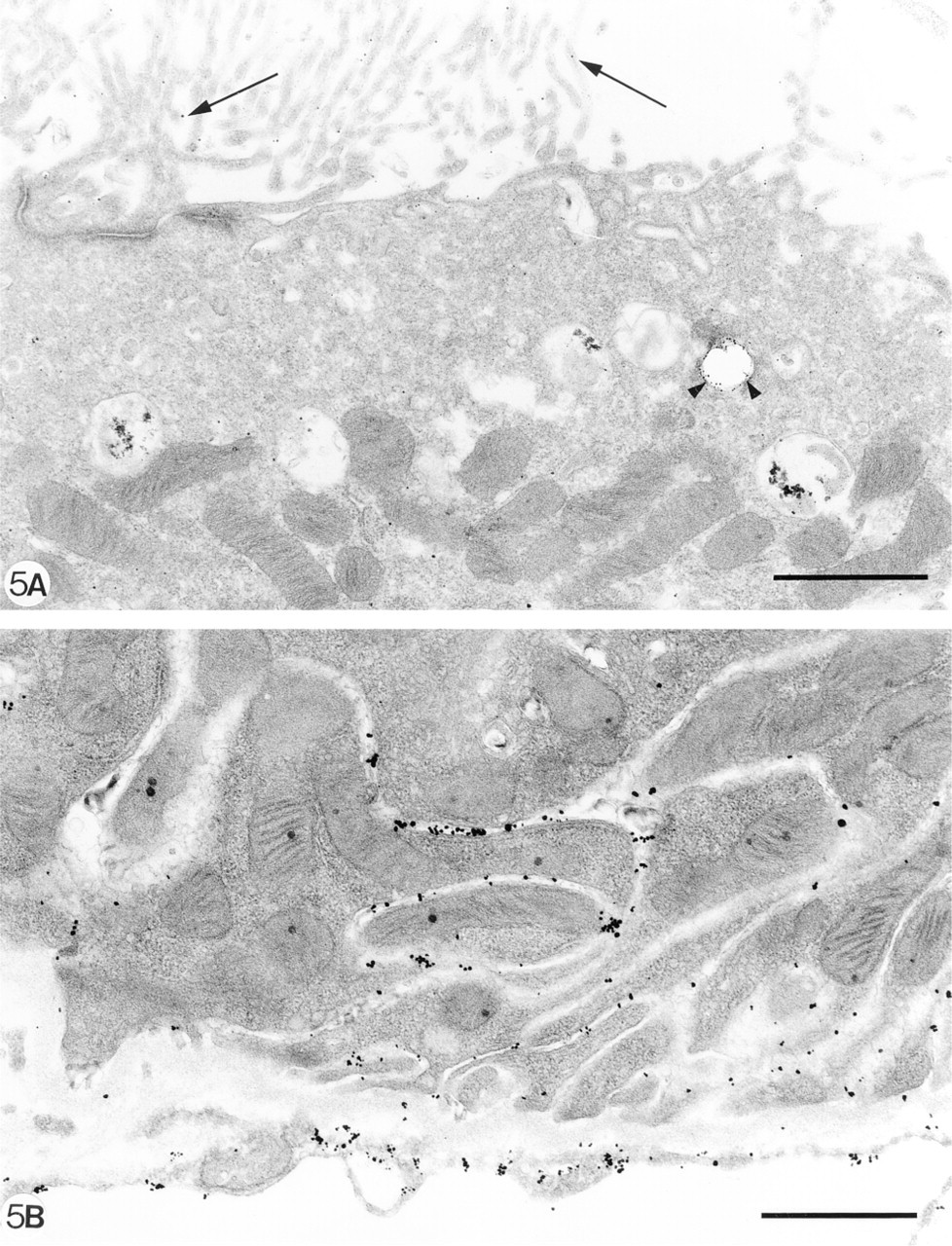
Direct immunoelectron microscopy (silver-enhanced immunogold technique) showing apical (
The data from our initial studies, in which we used high antibody doses, suggested that the reallocation of APA in the PTECs might be the result of a transcellular transfer of APA from the brush borders to the basolateral membrane. However, we could not substantiate such a routing because IEM failed to demonstrate transcellular transport. In addition, injection of a low dose of one of the anti-APA MAbs (ASD-41) resulted in similar binding to the basolateral side of the PTECs with the appearance of APA at that site, despite the absence of binding to the brush border membranes. This preferential rapid binding of IgG to the basolateral membranes can be explained by the fact that large molecular weight proteins more readily cross the endothelium of peritubular capillaries and the tubular basement membrane than glomerular capillaries (Venkatachalam and Karnovsky 1972). This suggests that binding of the injected antibodies to the brush borders is not a requirement for the appearance of APA at the basolateral membranes of the PTECs and that the formation of basolateral complexes of APA/anti-APA MAbs occurs independently from the formation of these complexes at the apical side (Fukatsu et al. 1988; Yamaguchi et al. 1996). We believe that the disappearance of APA from the brush borders and the appearance of APA at the basolateral membranes are separate phenomena, induced by two different mechanisms.
Immunofluorescence localization of the injected MAb ASD-41 (
Both endocytosis and shedding are involved in the disappearance of APA from the brush borders after injection of high doses of anti-APA MAbs. By IEM, we could demonstrate both APA and anti-APA MAbs in endocytotic vesicles just beneath the brush borders or in the upper part of the PTECs. These endocytosed immune complexes are believed to end up in lysosomes, where they are degraded (Helenius et al. 1983). In addition, we could demonstrate that APA and MAbs were directed to the tip of the brush borders and shed into the tubular lumen. In some brush borders of PTECs, APA was still present without bound anti-APA MAbs, suggesting that these particular PTECs were not exposed to the injected MAbs.
Shedding of immune complexes can be caused by two different mechanisms. First, cleavage of the antigen proximal to the plasma membrane can lead to removal of the immune complexes (Ehlers and Riordan 1991; Bazil and Horejsi 1993). APA indeed contains a cleavage site seven amino acids away from the membrane-spanning region (Wu et al. 1990; Hesp and Hooper 1995). Second, the immune complexes can be removed from the apical side of PTECs by pinching off small portions of the brush border membranes on which immune complexes have been formed, a process regulated by the cytoskeleton (Jacobsen et al. 1995). Several findings are in favor of this latter mechanism. First, by IEM both APA and MAbs are particularly located in the brush border vesicles shed into the tubular lumen. Second, we have previously demonstrated that APA in the mouse PTEC is directly coupled to γ-actin, an important cytoskeletal component (Mentzel et al. 1994). Third, treatment of mice with chlorpromazine, an anesthetic and calmodulin antagonist that is known to interfere with cytoskeletal activity and movement (Camussi et al. 1986; Andres et al. 1996), resulted in lack of modulation of APA in the brush borders of the PTECs.
The formation of basolateral immune complexes is caused by a different mechanism that acts independently from the endocytosis and shedding of APA at the apical side of the PTECs. Although little is known about the specific sorting of APA in the PTECs, the sorting of other brush border enzymes such as DPP IV and aminopeptidase N, type II glycoproteins that resemble APA both structurally and functionally (Wu et al. 1990; Shipp and Look 1993; Mentzel et al. 1996a,b), has been thoroughly investigated. For both enzymes it has been demonstrated that approximately 80% of newly synthesized protein is transported directly from the trans-Golgi network to the apical membranes. The other 20% is transported from the trans-Golgi network to the basolateral membranes and is subsequently endocytosed and transported to the apical brush border membranes (Casanova et al. 1991; Vogel et al. 1992, 1995; Weisz et al. 1992). This process is rapid (t1/2 $ 60 min) (Casanova et al. 1991), which may account for the almost absent expression of DPP IV and aminopeptidase N at the basolateral membranes of the PTECs (Mentzel et al. 1996a,b). Most likely, the intracellular routing of APA is identical to the routing of DPP IV and aminopeptidase N. One can now imagine that the injected anti-APA antibodies interfere with this routing by literally trapping this small pool of basolaterally sorted APA. This latter process also is dependent on cytoskeletal activity, because treatment with chlorpromazine prevented or considerably reduced the appearance of APA at the basolateral membranes. However, because chlorpromazine affects diverse biological processes, we cannot exclude the possibility that other mechanisms are involved.
Because APA plays such an important regulatory role in the renin-angiotensin system, the effects of this antigenic modulation of APA in the PTECs on the activity of Ang II must be taken into consideration. The concentrations of Ang II in the lumen of PTECs exceed the circulatory concentrations by 100-1000-fold (Braam et al. 1993), and the disappearance of APA from the brush borders by antigenic modulation will most likely result in reduced degradation of Ang II and even higher concentrations in the tubular lumen, which may have a profound effect on the proximal tubular fluid reabsorption (Edwards and Aiyar 1993). Furthermore, the proximal tubules express Ang II receptors at both the basolateral and brush border membranes (Harrison Bernard et al. 1997), which contribute to the removal of Ang II from the proximal tubular lumen or interstitial space by endocytosis (Hunyady et al. 1994a,b). The effects of the formation of APA/anti-APA MAb immune complexes on the regulation of Ang II receptors in PTEC are not known but will very likely result in physiological changes in the tubular activities of Ang II.
In conclusion, injection of anti-APA MAbs in mice results in modulation of APA in the PTECs. At the basolateral sides of the PTECs the injected antibodies trap a small pool of basolaterally sorted APA, whereas in the brush border immune complexes are removed by either endocytosis and degradation and/or by shedding of brush borders into the tubular lumen. These two separate processes are dependent on the activity of the cytoskeleton. The effect of the removal of APA from the apical side of the PTECs and the appearance at the basolateral side for Ang II activity and tubular cell function is not known but most likely results in increased activity of Ang II at the apical membranes and decreased activity at the basolateral membranes. Therefore, the antibody-induced modulation of APA in the PTEC might provide one additional, although theoretical, pathway of regulating the Ang II effects in the kidney.
Footnotes
Acknowledgements
Supported by grants from the Dutch Kidney Foundation (C91.1169 and C96.1534).