
Abstract
Placental leucine aminopeptidase (P-LAP), also called oxytocinase, is an enzyme responsible for hydrolyzing oxytocin. This enzyme is identical to cystine aminopeptidase. We examined the tissue distribution of P-LAP in normal adult mice and also in mothers and fetuses during mouse pregnancy using immunohistochemical (IHC) analysis. P-LAP-immunoreactive protein was expressed in various organs in a cell- and gestational stage-dependent manner. In the kidney, P-LAP was located in distal and collecting tubules but not in proximal tubules. The islet of Langerhans in the maternal pancreas stained positively for P-LAP in the periphery in early gestation. This staining pattern changed so that both the periphery and inner cells were positive during mid-gestation and finally only inner cells were positive in late gestation. Among the hematopoietic cells in the fetal liver, only megakaryocytes showed strong expression of P-LAP. The staining intensity increased with gestation on the apical surface of trophoblasts in the placental labyrinth. These data demonstrate that P-LAP is present in a variety of tissues, and its presence is affected by pregnancy and fetal development. Therefore, P-LAP may play a significant role in various physiological processes in non-pregnant, pregnant, and fetal mice.
A
Placental leucine aminopeptidase (P-LAP) is an enzyme responsible for cleaving the N-terminal ring structure of oxytocin (OT), and is also called oxytocinase. Moreover this enzyme is identical to cystine aminopeptidase (CAP; EC 3.4.11.3) (Tsujimoto et al. 1992). The findings that serum P-LAP activity rises progressively with gestational age until near term (Carter et al. 1974; Mizutani et al. 1976; Yamahara et al. 2000) and decreases in patients with spontaneous preterm delivery (Kozaki et al. 2001) suggest that this enzyme is involved in fetal development and the regulation of the uterotonic effects of OT. Several groups, including ours, have isolated a P-LAP cDNA clone from a human placental cDNA library (Rogi et al. 1996; Laustsen et al. 1997). Structural analysis has revealed that P-LAP is a type II membrane-spanning protein belonging to the M1 family of zinc metal-lopeptidases, which also contains APN/CD13, NEP/CD10, and APA/BP-1. Contrary to the initial hypothesis that P-LAP is restricted to the placenta, a wide-spread distribution of P-LAP in human tissues was first demonstrated by Northern blotting analysis (Rogi et al. 1996) and was later confirmed at the protein level by IHC (Nagasaka et al. 1997). Interestingly, an enzyme homologous to P-LAP, with 87% identity at the amino acid level, has been cloned from a rat adipocyte cDNA library and named insulin-regulated membrane aminopeptidase (IRAP) (Keller et al. 1995). IRAP is present in glucose transporter isotype GLUT4 vesicles of rat adipocytes and is co-translocated from the cytosol to the cell membrane with GLUT4 by insulin stimulation in adipocytes and skeletal muscle cells (Kandror et al. 1994; Mastick et al. 1994). Therefore, IRAP has now also been examined in adipose tissue and muscles. The roles of P-LAP, especially in tissues other than placenta, remain unclear. To address the potential functional roles of P-LAP, it would be helpful to establish the precise location of P-LAP and to examine the changes in expression levels and localization during physiological processes, including pregnancy and development. Moreover, the evidence that P-LAP composes a family with APN, NEP, and APA, which are established as cell differentiation markers, suggests that P-LAP expression might be associated with cell growth and differentiation in some tissues.
The mouse is one of the best-studied mammalian experimental models and research is not restricted by the inherent ethical and practical limitations associated with humans. In this study we investigated the P-LAP expression pattern in mice to find clues to its physiological functions not only in placenta but also in other organs. To achieve this goal, we first examined the tissue distribution of P-LAP by Western blotting and IHC in normal adult mice aged 8-12 weeks. We then conducted a series of experiments using IHC procedures in maternal and fetal mouse tissues to determine the effects of pregnancy and development on P-LAP expression in a variety of tissues.
Materials and Methods
Animals and Tissue Preparation
C57/BL6J mice were purchased from Chubu Kagaku Shizai (Nagoya, Japan). Mice aged 8-12 weeks were bred and maintained in our animal facility under climate-controlled conditions with a 12-hr light/dark cycle and were allowed free access to standard chow and drinking water. The local council on animal care approved experimental protocols. The day after timed mating was designated as day 1 of gestation. Maternal and fetal mouse tissues were obtained at a particular day of pregnancy from mice (six maternal and six fetal mice at each gestational day) anesthetized by IP administration of ketamine (80 mg/kg) and xylazine (16 mg/kg). Six non-pregnant female mice of the same strain with similar ages to pregnant mice were used as controls. Tissue samples for IHC were fixed in 10% formalin and embedded in paraffin until use.
Western Blotting Analysis
Western blotting analysis was performed as described previously (Nomura et al. 1991) with slight modification. Fresh tissue samples were collected from three non-pregnant mice aged 10 weeks and were quickly washed to remove blood and secretions with cold PBS on ice. They were immediately frozen in liquid nitrogen, crushed, and homogenized in an extraction buffer containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 5 mM EDTA, 2 mM phenylmethyl sulfonyl fluoride, 5 μg/ml leupeptin, 2% Triton X-100, and 5% 2-mercaptoethanol, followed by centrifugation at 12,000 × g for 5 min. The supernatant was collected as protein extract from each individual sample. Protein concentration was determined by BCA protein assay reagent (Pierce; Rockford, IL). Samples diluted in 2 × SDS buffer [125 mM Tris-HCl (pH 6.8), 4% SDS, 10% glycerol, 0.2% bromphenol blue, and 4% 2-mercaptoethanol] were resolved on 7.5% SDS-polyacrylamide gels under reducing conditions and then transferred to a PVDF membrane (Immobilon; Millipore, Bedford, MA). After blocking the nonspecific binding with 5% nonfat skim milk in PBS with 0.05% Tween-20 (TPBS), the membrane was incubated with anti-P-LAP polyclonal antibody (1:1000 dilution in TPBS) (Matsumoto et al. 2000) or anti-GAPDH antibody at 4C overnight. It was then washed in TPBS four times and incubated with peroxidase-conjugated swine anti-rabbit IgG (Bio-Rad; Hercules, CA). Positive bands were visualized using the enhanced chemiluminescence (ECL) Western blotting detection kit (Amersham Pharmacia Biotech; Piscataway, NJ). Experiments were repeated three times using three individual samples.
Immunohistochemical Staining
The tissues used were kidney, liver, lung, brain, heart, thymus, intestine, stomach, duodenum, spleen, pancreas, ovary, placenta, and uterus. Thin sections of 4-μm thickness were cut (at least three sections from each of the tissues in each mouse) and mounted on silane-coated slides (Matsunami; Tokyo, Japan). These sections were immunostained by the streptavidin-biotin-peroxidase method (Hsu et al. 1981). Sections were deparaffinized with xylene and microwaved (500 W) at 90C for 10 min in citric acid buffer (2 mM citric acid and 9 mM trisodium citrate dehydrate, pH 6.0) for antigen retrieval. These reacted sections were incubated with normal goat serum for 30 min. The slides were further incubated with primary antibody for rabbit anti-P-LAP polyclonal antibody (1:100 dilution) and reacted with methanol that contained 0.3% H2O2 for 30 min to block any endogenous peroxidase. The antigen-antibody complex was then visualized with 3.3′-diaminobenzidine (DAB) solution (1 mM DAB, 50 mM Tris-HCl buffer, pH 7.6, and 0.006% H2O2). Chromogenic development was performed as described by Graham et al. (1965). The slides were counterstained with Meyer's hematoxylin. Sections not exposed to primary antibody were always included as negative controls; none of them showed immunostaining. Slides were examined using a light microscope (Nikon E 600; Nikon, Tokyo, Japan) and photographed with Coolpix P700c (Nikon).
Results
P-LAP Western Blotting
Western blotting analysis against P-LAP showed a single band of approximately 170 kD, which is similar in size to human P-LAP (Figure 1, Lane 1), in all non-pregnant mouse tissues examined (Figure 1, Lanes 3-10). Negative control using non-pregnant woman serum showed no bands (Figure 1, Lane 2), indicating that the antibody specifically reacted with mouse P-LAP. Immunoreactive P-LAP proteins in the liver and forebrain ran faster, and those in the lung, kidney, uterus, and heart showed broad bands. The signal intensities obtained using anti-GAPDH antibody instead of anti-P-LAP antibody were comparable among the lanes, indicating that equivalent quantities of the samples were loaded in each lane (data not shown). This widespread tissue distribution of P-LAP at the protein level prompted us to investigate the distribution using IHC staining to find the precise cellular localization of P-LAP.
P-LAP Distribution in Adult Mouse Tissues
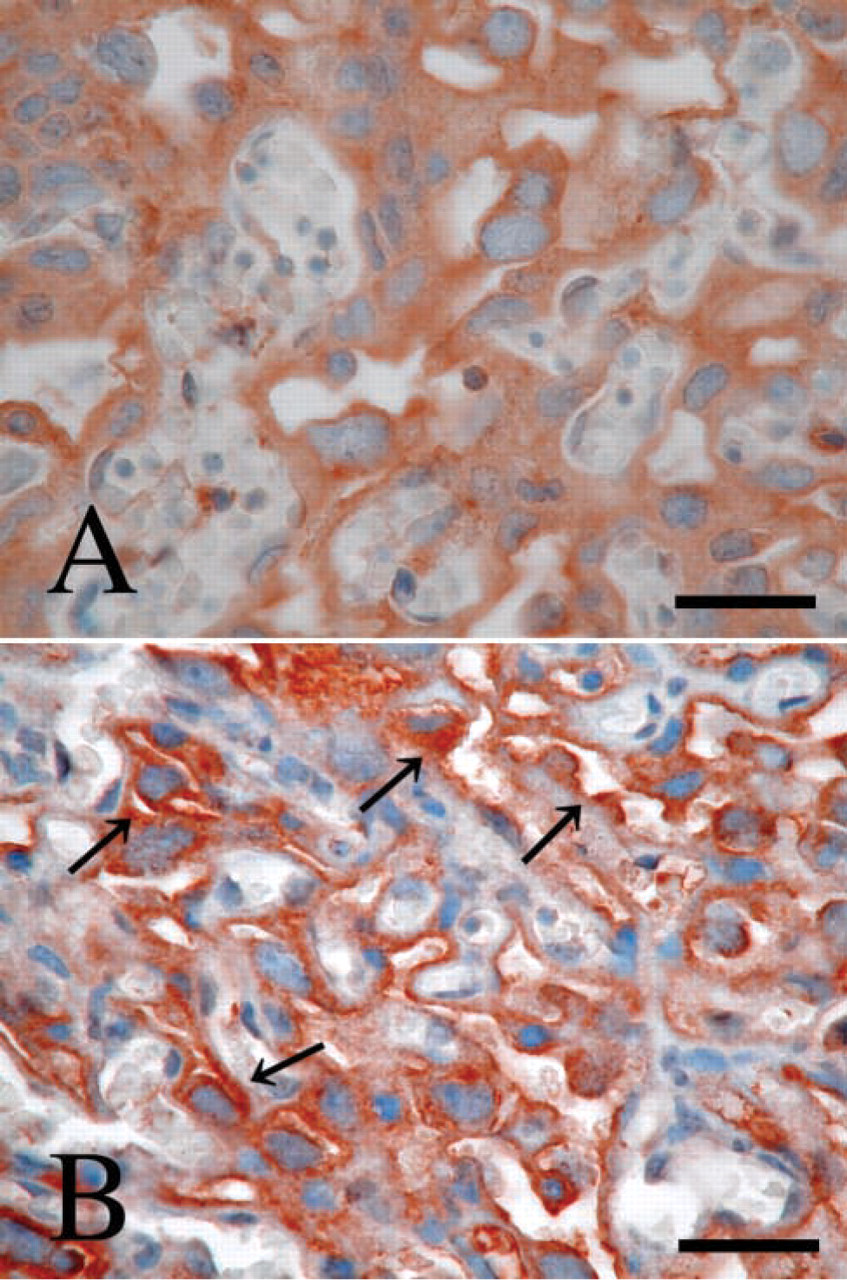
Next, we examined P-LAP localization in various organs of adult mice using IHC. We assessed the staining by examining 18 sections obtained from six mice for each tissue, the results of which are summarized in Table 1. Strong expression was noted in the pit epithelium of the stomach, cardiomyocytes of the heart, enterocytes in the intestine, bile canaliculi in the liver, islet of Langerhans, and distal and collecting tubules in the kidney. In the lung, we observed moderate immunoreactivity in bronchial epithelium but no immunoreactivity in alveolar epithelium. In the brain, including both cerebellum and cerebrum, neuronal cells showed positive immunoreactivity but glial cells did not. Other sites with moderate staining included follicle epithelium of the ovary and endometrial gland of uterus. Among hematopoietic cells in bone marrow, only megakaryocytes showed weakly positive staining. Not every tubule in the kidneys had equal P-LAP-positive staining. Under low magnification, immunoreactive P-LAP protein was clearly seen in the distal tubules in the cortex and collecting tubules in the medulla (Figure 2A). Under high magnification, moderate expression was observed in endothelial cells of the glomerulus (Figure 2B) and strong expression in the apical surface of the distal tubule (Figure 2C), but no staining was observed in the proximal tubule (Figure 2D).
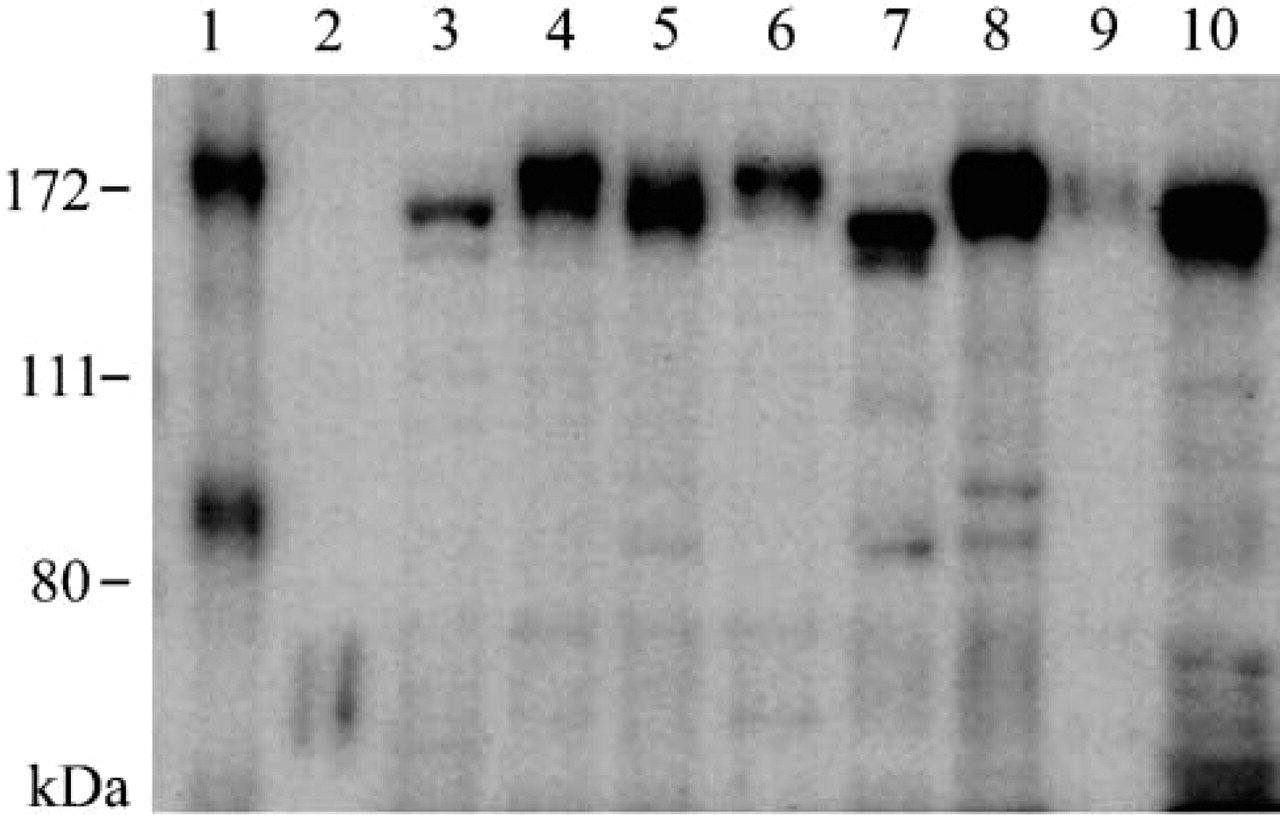
Western blotting analysis of P-LAP in various mouse tissues. Proteins (7 μg) prepared from normal human endometrium (Lane 1), non-pregnant woman serum (Lane 2), adult mouse liver (Lane 3), lung (Lane 4), kidney (Lane 5), uterus (Lane 6), fore-brain (Lane 7), heart (Lane 8), spleen (Lane 9), and pancreas (Lane 10) were analyzed using the anti-P-LAP antibody. Molecular weight markers are indicated at left.
Immunolocalization of P-LAP in normal mouse tissues a
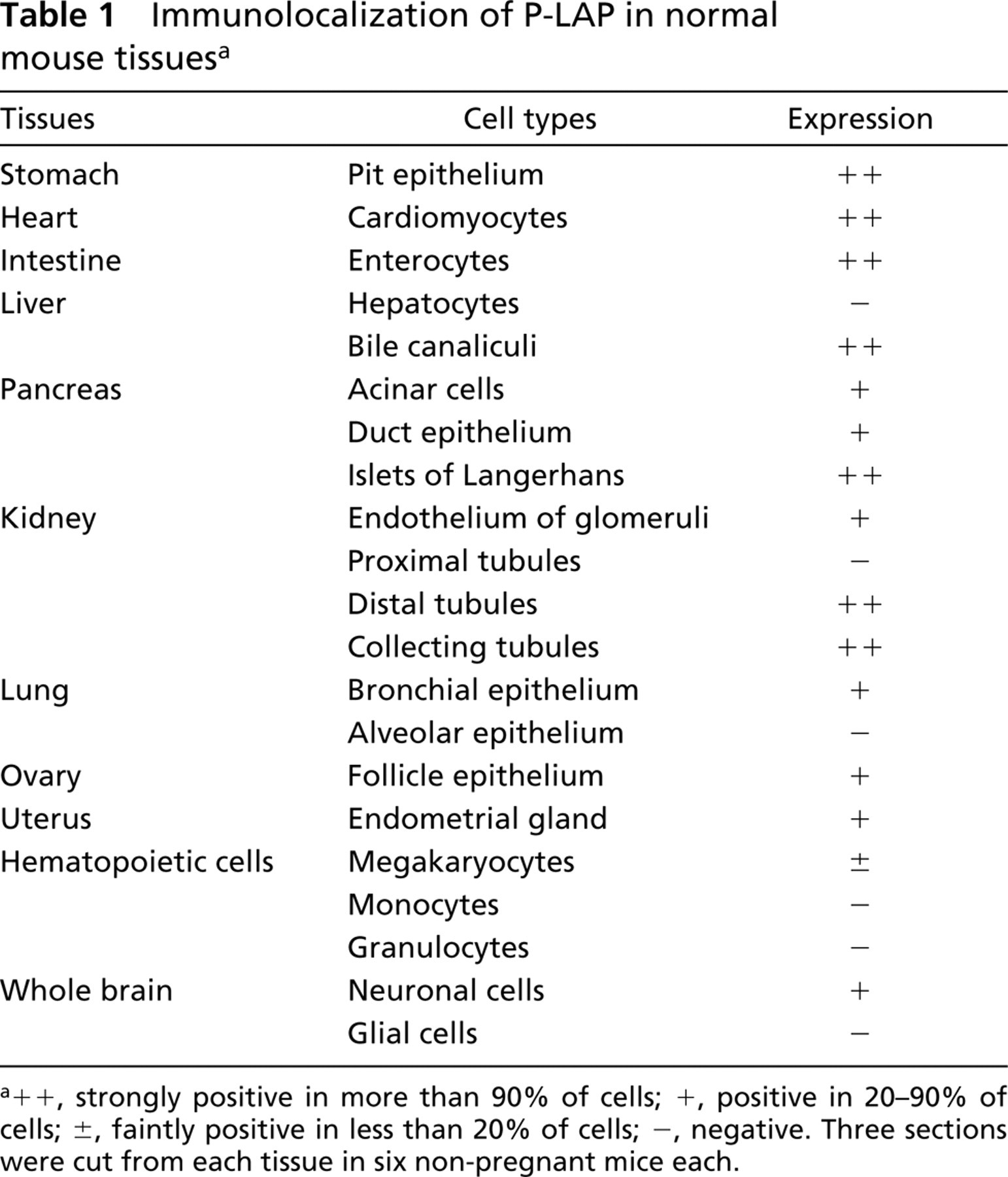
a++, strongly positive in more than 90% of cells; +, positive in 20–90% of cells; ±, faintly positive in less than 20% of cells; –, negative. Three sections were cut from each tissue in six non-pregnant mice each.
P-LAP Distribution in Maternal and Placental Tissue
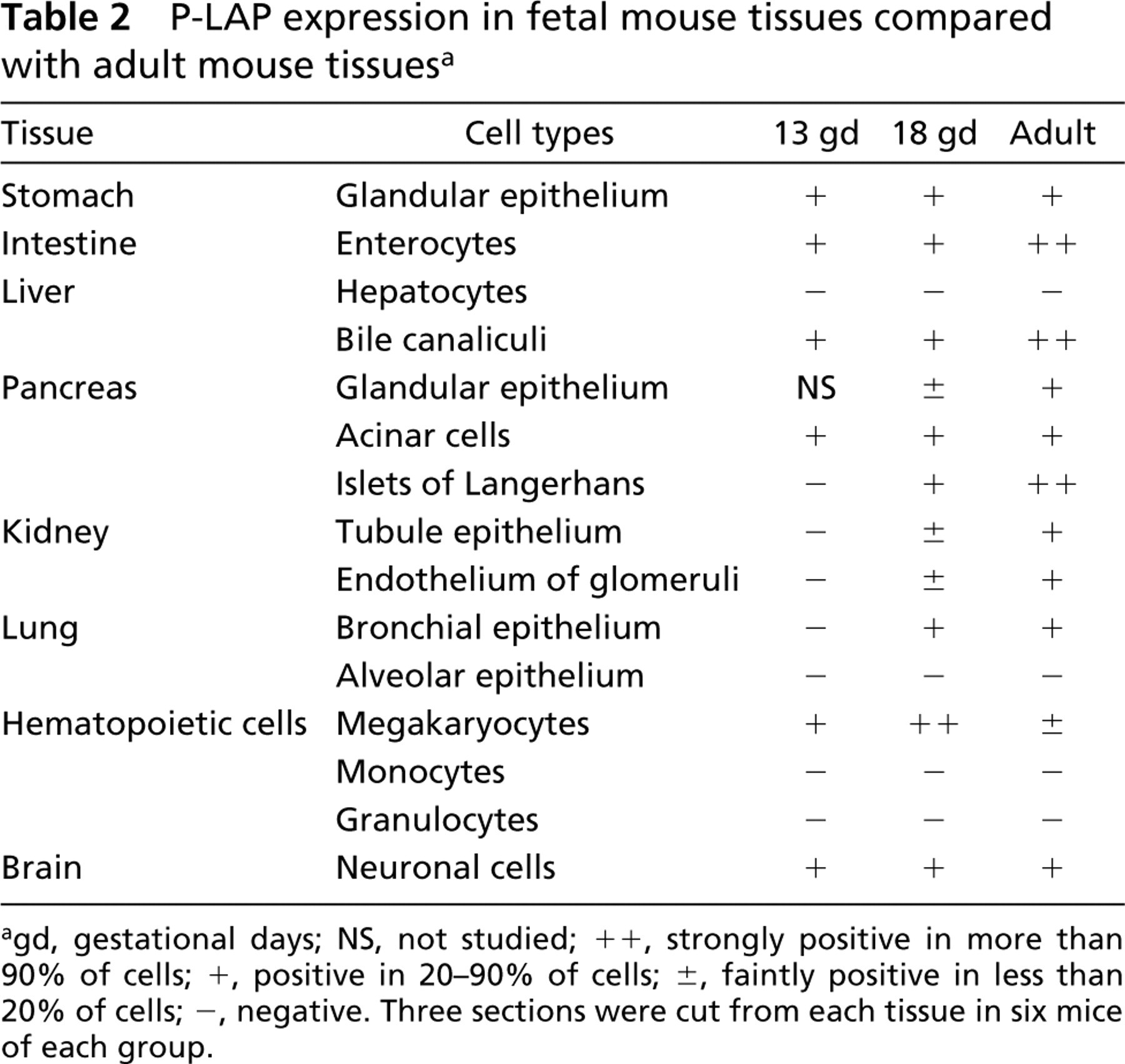
Next we investigated the P-LAP immunoreactivity in pregnant mouse tissues to examine whether or not P-LAP staining patterns change during pregnancy. In contrast to tissues such as stomach, intestine, liver, kidney, lung, ovary, and brain, which showed no clear alterations (data not shown), the pancreatic islet of Langerhans revealed apparent changes in the P-LAP staining pattern during pregnancy. In non-pregnant adult mice, strong immunoreactivity was prominent only in the periphery of the islet (Figure 3A). A similar staining pattern was observed on gestational day 6 (Figure 3B). However, on gestational day 13, P-LAP-reactive cells were observed both in the periphery and in the inner area of the islet (Figure 3C). On gestational day 18, P-LAP expression in the periphery decreased and only the cells in the inner area showed mosaic-like staining patterns (Figure 3D). At this time we also could observe enlargement of the islet volume. In the placenta, cells with weakly positive staining of P-LAP were observed throughout the tissue on gestational day 6. It could not be determined whether these were decidual or trophoblastic cells because the labyrinth was still immature at this stage. On gestational days 13 (Figure 4A) and 18 (Figure 4B), the trophoblast cells of the placental labyrinth facing the maternal blood space demonstrated strong P-LAP expression, the staining intensity of which was stronger at gestational day 18 than at gestational day 13.
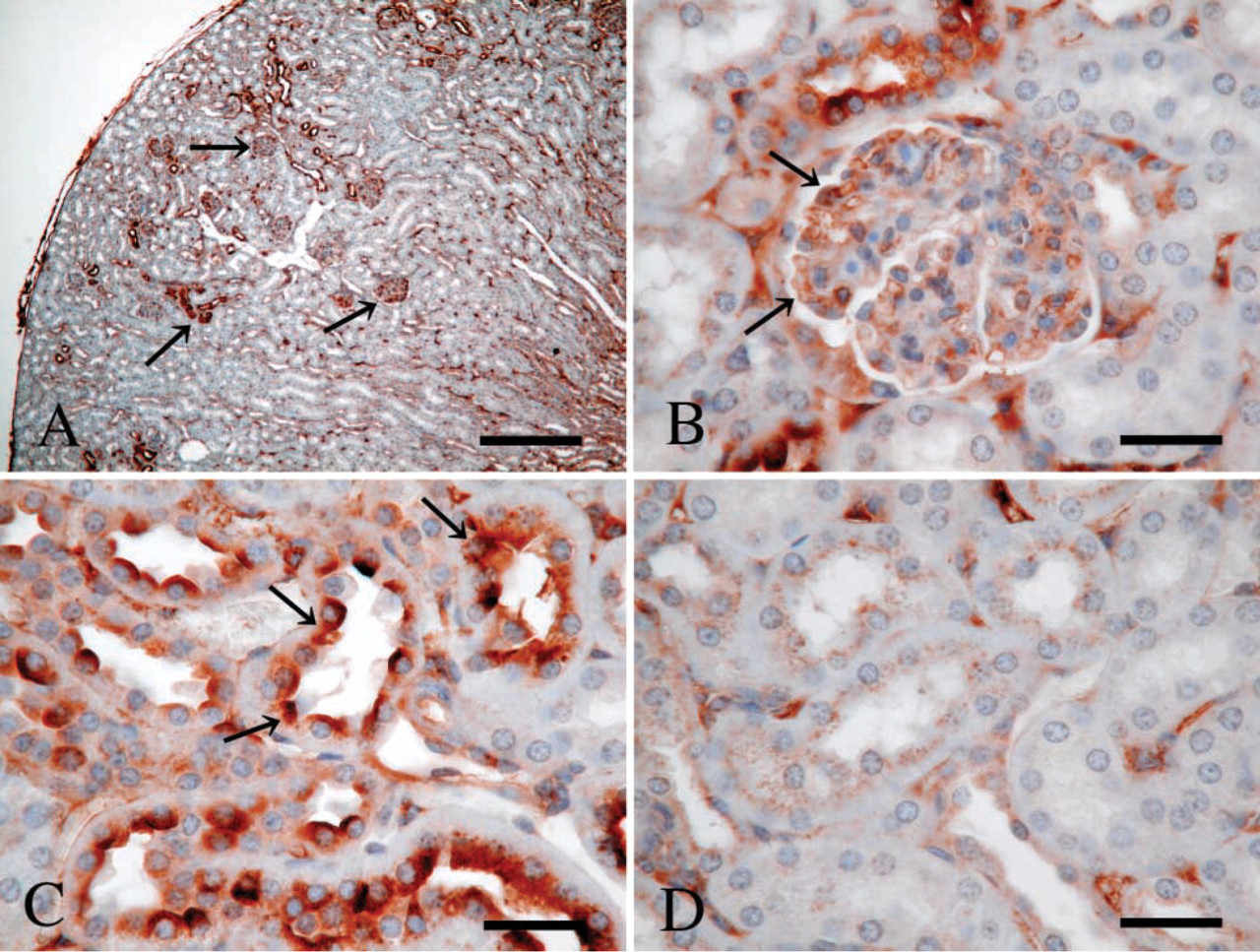
Immunohistochemistry of P-LAP in normal mouse kidney. (
Fetal Tissue Distribution of P-LAP
We also examined the P-LAP localization in fetal mouse tissues of gestational days 13 and 18. The results are summarized in Table 2. During this period of gestation, glandular epithelium of the stomach, bronchial epithelium of the lung, neuronal cells of the brain, and endothelium of bile canaliculi in the liver showed positive P-LAP expression, which was similar to the results in adult mice. However, in contrast with the faint staining in adults, megakaryocytes in fetal liver showed strong P-LAP staining on gestational day 13 (data not shown) and more intense staining at gestational day 18 (Figure 5A) in the cytoplasm. Interestingly, the megakaryocytes at this time had small and non-lobulated nuclei, suggesting that they were still immature. We also looked closely at the kidneys because renal function remains immature in the fetus. A moderate staining pattern was detected in the cortex at gestational day 18 (Figure 5B). Mild to moderate staining was found in the medulla, but it was difficult to determine in detail the localization of P-LAP in the different types of renal tubules, possibly because of the structural immaturity. The P-LAP staining pattern in the pancreas also differed between fetus and adult. Fetal islet of Langerhans had weaker immunoreactivity and a smaller number of positive cells in the periphery even at gestational day 18 compared with that in the adult (data not shown).
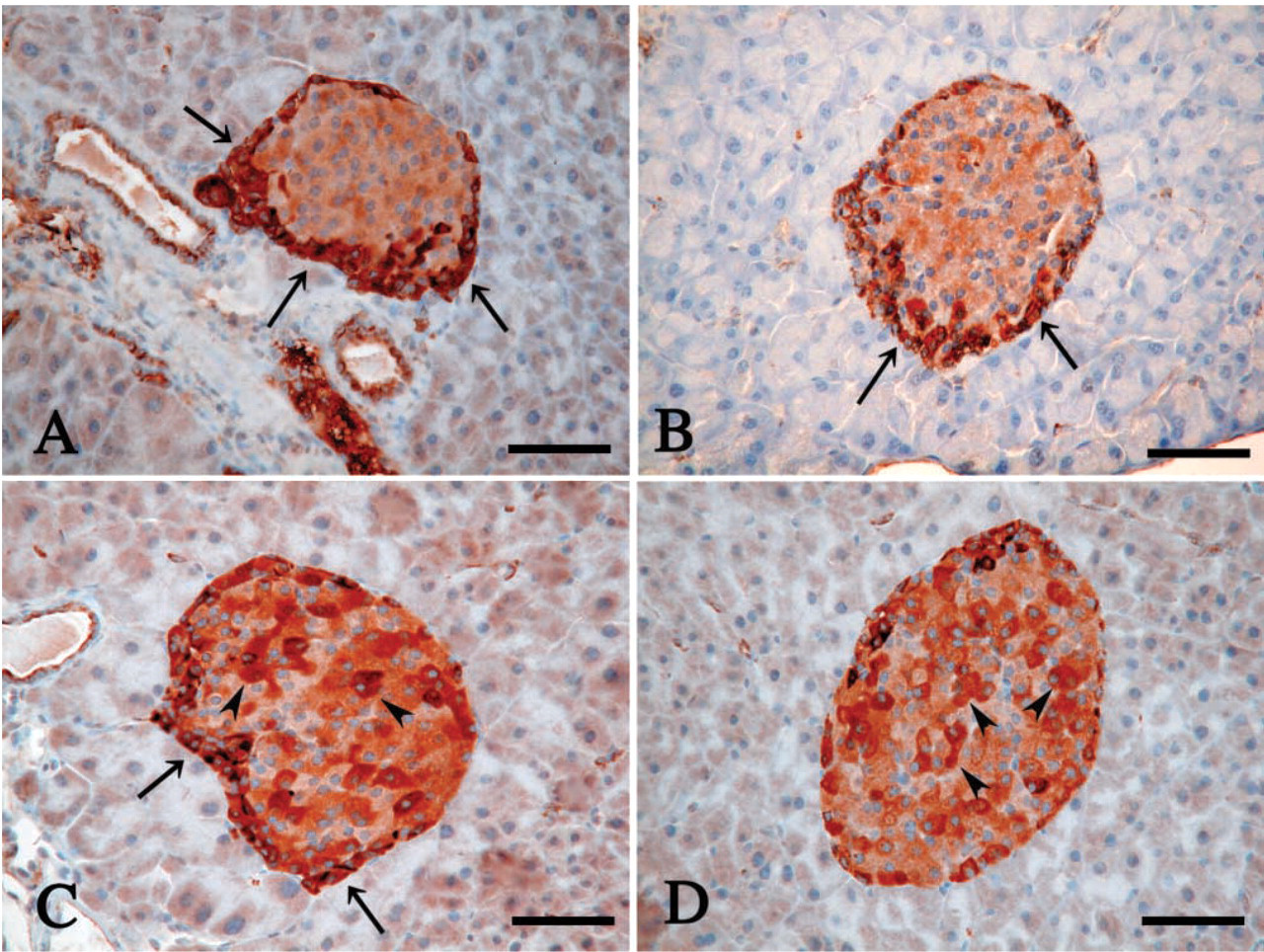
Immunohistochemistry of P-LAP expression in the islet of Langerhans. Mice used were non-pregnant (
Discussion
P-LAP was previously regarded as a placenta-specific enzyme, but widespread distribution of P-LAP in human tissues has now been established (Rogi et al. 1996; Nagasaka et al. 1997). In this study we examined the P-LAP staining pattern in non-pregnant mice and compared it with that of pregnant mice and their fetuses. We observed immunoreactivity against P-LAP in various endothelial and epithelial cells in non-pregnant mice, which was consistent with the results in humans (Nagasaka et al. 1997). In addition, we also observed changes in the P-LAP staining patterns in several organs in mother and fetus during pregnancy. These results strongly suggest the involvement of P-LAP in many physiological processes, including functions related to maintaining homeostasis during pregnancy and regulating fetal development.
The roles of P-LAP in vivo have not yet been clearly elucidated. Human P-LAP hydrolyzes bioactive peptides, including OT as well as vasopressin and angiotensin III (Tsujimoto et al. 1992; Matsumoto et al. 2000), which corresponds to the substrate specificity of rat IRAP, a homologue of P-LAP (Herbst et al. 1997). Therefore, it is highly likely that P-LAP regulates peptide functions through the metabolism of peptide. Furthermore, the co-localization and insulin-stimulated co-translocation of P-LAP with GLUT4 may hint at a role of P-LAP in glucose homeostasis. This possibility appears likely because the amino terminus of IRAP/P-LAP causes GLUT4 translocation, which results in increased glucose uptake in insulin-sensitive cells (Waters et al. 1997). Conversely, however, recently developed IRAP/P-LAP-deficient mice maintain normal glucose homeostasis even though the mice have reduced levels of GLUT4 in skeletal muscle and adipose tissues (Keller et al. 2002). Further detailed characterization of the mice would help to elucidate the physiological roles of P-LAP, as was the case with APA-deficient mice. APA-deficient mice have shown increased baseline blood pressure and enhanced the hypertensive response to AngII infusion, indicating that APA functionally regulates a circulatory system via metabolization of AngII (Mitsui et al. 2003).
P-LAP expression in fetal mouse tissues compared with adult mouse tissues a
agd, gestational days; NS, not studied; ++, strongly positive in more than 90% of cells; +, positive in 20–90% of cells; ±, faintly positive in less than 20% of cells; –, negative. Three sections were cut from each tissue in six mice of each group.
Immunohistochemistry of P-LAP expression in the placenta. (
Among the tissues investigated, Western blotting showed high expression levels of P-LAP in the lung, kidney, heart, and pancreas. Because serum from non-pregnant women is known to contain no P-LAP proteins (Yamahara et al. 2000), we used it as a negative control. A dominant single band of the approximately expected size indicated that the antibody used in this study, which was raised against human P-LAP, reacted to mouse P-LAP specifically. The apparent differences in molecular weights of P-LAP among the tissues might depend on differences in the glycosylation in each tissue. The expression levels of P-LAP were further evaluated by IHC, which could also determine the precise localization. For example, in the lung, which showed an intense signal in Western blotting, remarkable staining was noted in the vascular endothelium compared with moderate staining in the bronchial epithelium and negative staining in the alveolar epithelium.
Because vasopressin, a key peptide for regulating water balance, is a representative target of P-LAP (Tsujimoto et al. 1992; Matsumoto et al. 2000), it would be reasonable to assume that P-LAP plays a critical role in regulating vasopressin action in the kidneys. Vasopressin functions in water reabsorption in principal cells of collecting ducts by inducing the translocation of water-channel protein aquaporin-2 from the cytosol to the plasma membrane (Nielsen et al. 1995). We detected immunoreactive P-LAP protein in adult mice in the distal tubules in the cortex and collecting tubules in the medulla, but not in the proximal tubules, which overlaps with the distribution of the vasopressin V2 receptor (Ostrowski et al. 1993). In addition, we have very recently found that P-LAP is translocated to the cell surface by vasopressin stimulation in renal cells (Masuda et al. 2003), which is similar to the finding that OT translocates P-LAP to the cell surface in vascular endothelial cells (Nakamura et al. 2000). Therefore, we can speculate that P-LAP functions in regulating excess vasopressin levels with a negative feedback system to maintain water homeostasis.
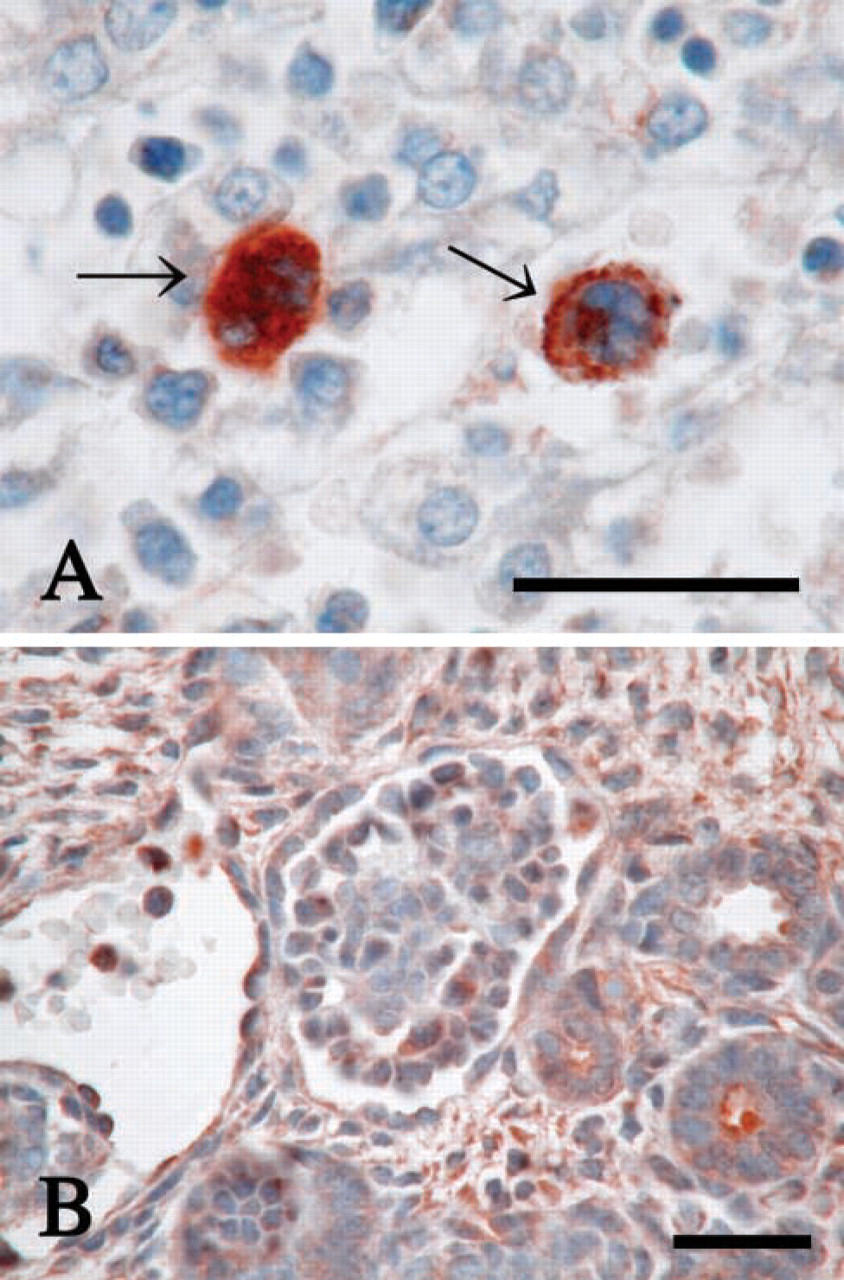
Immunohistochemical staining of P-LAP in the fetal tissues at gestational day 18. (
Western blotting demonstrated the high expression of P-LAP in the heart, which has been also reported in humans (Rogi et al. 1996). In this study, P-LAP was assigned immunohistochemically to cardiomyocytes in the heart, in which GLUT4 is co-translocated with IRAP/P-LAP by insulin stimulation (Slot et al. 1991), as is observed in adipocytes and skeletal muscle cells. High expression of P-LAP in the heart might be associated with GLUT4 gene expression. The finding that IRAP/P-LAP expression in the heart is reduced in GLUT4-deficient mice (Jiang et al. 2001) supports this postulation.
We observed a dramatic change in the P-LAP staining pattern in maternal islet of Langerhans, with positive immunoreactivity in the peripheral cells of the islet in early gestation, in both peripheral and inner cells in mid-gestation, and in inner cells only in late gestation. The peripheral pancreatic islets represent predominantly glucagon-producing cells (A-cells), while inner cells most likely express insulin (B-cells). Although further investigation is required to identify the cell types expressing P-LAP in the islet of Langerhans, we believe that this change in P-LAP localization may be associated with pancreatic functions. The normal endocrine pancreas adapts to the metabolic changes of pregnancy by islet hypertrophy, accompanied by increases in the number and granulation of B-cells, which leads to the elevation of insulin secretion (Aerts and Van Assche 1977; Ordorica and Frieden 1991). The findings that OT and vasopressin are present in the pancreas (Amico et al. 1988; Johansson et al. 1991) and that they directly increase the release of insulin (Dunning et al. 1984a; Gao et al. 1990; Richardson et al. 1990) and glucagon (Dunning et al. 1984b; Gao et al. 1991) indicate the possible roles of those peptides in regulating the endocrine function of the pancreas. In non-pregnant mice and in pregnant mice at an early gestational stage, P-LAP may downregulate glucagon production through metabolizing OT and vasopressin. As gestation progresses, increased P-LAP, probably in B-cells, may suppress insulin secretion to counteract the excessive OT and vasopressin. Although speculative, such regulatory mechanisms would contribute to maintaining glucose homeostasis during pregnancy.
Of greatest interest in the fetus is the P-LAP expression in liver megakaryocytes. In the fetal liver, hepatocytes, monocytes, and granulocytes showed no P-LAP immunoreactivity throughout development; only megakaryocytes expressed P-LAP immunoreactivity. This immunoreactivity appeared to be enhanced in late gestation compared with mid-gestation, suggesting the possible involvement of P-LAP in mega-karyocytopoiesis. No studies to date have addressed the role of P-LAP in hematology. However, several lines of evidence indicate that the involvement of P-LAP in megakaryocytopoiesis and hematopoietic cell differentiation is very likely. Ikaros, a transcription factor first identified as a regulator of lymphocyte differentiation (Georgopoulos et al. 1992), also regulates P-LAP gene transcription in trophoblasts (Ito et al. 2002). Therefore, P-LAP expression may be regulated by some transcription factors including Ikaros along with megakaryocyte differentiation. In addition, various types of hematopoietic cells have been shown to express their own cell surface aminopeptidases at unique stages of cell differentiation lineage (Kenny et al. 1989; Shipp and Look 1993). Therefore, P-LAP may serve as a cell-specific marker for megakaryocytes. In addition to the increased concentrations of megakaryocytes, their DNA content and size are increased in late gestation (Jackson et al. 1992), coinciding with the stage when P-LAP expression in mega-karyocytes increases. Several factors are required in megakaryocyte growth and differentiation, including thrombopoietin (de Sauvage et al. 1994; Kaushansky et al. 1994; Lok et al. 1994; Wendling et al. 1994) and the recently described placenta-specific prolactin-like proteins (Lin and Linzer 1999; Zhou et al. 2002). P-LAP is unlikely to hydrolyze these molecules directly because of their large size, but small peptides, including oxytocin and angiotensin III, which megakaryocytes actively take up (Daimon 1991), may regulate the activities of these growth factors in megakaryocytes. We can speculate that P-LAP might participate in megakaryocytopoiesis by metabolizing small effector peptides.
The presence of P-LAP on apical membranes of trophoblasts of the placental labyrinth in mice and the increase of staining intensity during pregnancy are similar to the findings in humans (Nagasaka et al. 1997; Yamahara et al. 2000). In mouse placenta, a cytoplasmic wall of trophoblasts is reduced in thickness as gestation progresses (Suzuki et al. 1997), which would seem to be beneficial to meet the increased demand for the exchange of nutrients and wastes. Increased P-LAP activity on the cell surface of trophoblasts may be cooperatively associated with the exchange through degrading the proteins.
In the present study, we determined the P-LAP location in non-pregnant, pregnant, and fetal mice. Our results may open the way to examine the roles of P-LAP in vivo, which will require further experiments investigating its natural substrates in each tissue.
Footnotes
Acknowledgements
Supported in part by Grants-in-Aid from the Ministry of Education, Culture, Sports, Science and Technology of Japan, and from the Ministry of Public Management, Home Affairs, Posts and Telecommunications of Japan for specific medical research (in collaboration with Nagoya Teishin Hospital).
We thank Hitomi Sato and Kazuko Matsuba for expert technical help.