
Abstract
Hakata antigen was first reported as a serum protein that reacted with an autoantibody from patients with systemic lupus erythematosus. Recently, it has been found that Hakata antigen is a new member of the ficolin/opsonin p35 family, which is a distinct lectin family, on the basis of homology of structures and the common characteristic of possessing lectin activity. In this study we analyzed the tissue distribution of Hakata antigen. Hakata antigen mRNA and protein were generated in the lung and liver. In the lung, Hakata antigen was produced by both ciliated bronchial epithelial cells and Type II alveolar epithelial cells and was secreted into the bronchus and alveolus. In the liver, Hakata antigen was produced by bile duct epithelial cells and hepatocytes and was also secreted into the bile duct. These results demonstrate that Hakata antigen is a unique lectin protein that exists not only in serum but also in bronchus/alveolus and bile, and indicate that Hakata antigen plays a role in bronchus/alveolus and bile under physiological conditions.
H
We have recently identified the cDNA encoding Hakata antigen, revealing that Hakata antigen consists of a collagen-like domain in the middle section and a fibrinogen-like domain in the C terminus, both of which are homologous to two lectin activity-possessing proteins, human ficolin-1 and opsonin p35 (Sugimoto et al. 1998). Homologies between Hakata antigen and either human ficolin-1 or opsonin P35 were both 48% overall, higher in the collagen-like domain (48%, 54%) and in the fibrinogen-like domain (52%, 53%), respectively (Sugimoto et al. 1998). We have further demonstrated that purified Hakata antigen sustains lectin activity, showing affinity with GalNac, GlcNac, and
Our present analysis of the tissue distribution of Hakata antigen reveals that it is produced by ciliated bronchial epithelial cells and Type II alveolar epithelial cells in the lung and by bile duct epithelial cells and hepatocytes in the liver, which are secreted into bronchus/alveolus and bile duct, respectively.
Materials and Methods
Northern Blotting Analysis
A membrane filter blotted with 2 μg of poly(A)+ mRNA from human tissues (Clontech Labs; Palo Alto, CA) was hybridized with 32 P-labeled Hakata antigen cDNA fragment (nucleotides 448-864) overnight at 42C in a solution containing 50% formamide, 5 × Denhardt's solution, 5 × SSPE, 0.5% sodium dodecyl sulfate (SDS), and 200 μg/ml salmon sperm DNA. After two extensive washings at room temperature (RT) for 10 min in a 0.1 × standard saline citrate (SSC)/0.1% SDS solution, the filter was exposed to an autoradiographic imaging system and the image was then read using a bioimaging analyzing system (BAS 1000; Fuji Photo Film, Minamiashikaga, Japan).
Preparation of Polyclonal and Monoclonal Antibodies Against Hakata Antigen
A polyclonal antibody against Hakata antigen was produced as described previously (Sugimoto et al. 1998). A monoclonal antibody against Hakata antigen was produced using Hakata antigen denatured by pyridylethylation as antigen (Cavins and Friedman 1970). Each 200 μ g of denatured Hakata antigen was injected along with complete Freund's adjuvant (Difco Laboratories; Detroit, MI) into Balb/c mice. After 4 weeks the mice were boosted with 200 μ g of the antigen without adjuvant, and 3 days later spleens were removed to fuse with P3U1 myeloma cells. Hybridoma cells were screened by using an ELISA with Hakata antigen. One of the positive clones was designated Pr19. Antibodies were then collected from ascites of mice bearing the Pr19 hybridoma cells. Pr19 belonged to the IgG1 isotype.
Western Blotting Analysis
Fresh human lung and liver tissues were taken after informed consent from donors undergoing surgery, and the tissues were immediately frozen. Each frozen tissue was homogenized with 50 mM Tris-HCl (pH 8.0) solution containing 8 M urea, 4% SDS, and 5 mM EDTA, and the supernatant was collected after centrifugation. Hakata antigen in serum was purified according to the method used by Yae et al. (1991). Bronchoalveolar lavage (BAL) fluids from inpatients were provided by the Department of Pathology, Kyushu University Hospital, and were used after removal of cells. Bile samples from inpatients, collected by percutaneous transhepatic cholangio drainage tubes, were provided by the First Department of Surgery, Kyushu University Hospital.
Total homogenates of the human lung and liver tissues, purified Hakata antigen, BAL fluids, and bile were applied to SDS-PAGE and transferred electrophoretically to PVDF membranes (Millipore; Bedford, MA). The membranes were probed with either a polyclonal or a monoclonal antibody against Hakata antigen and were visualized by enhanced chemiluminescence (Tropix; Bedford, MA).
Immunohistochemical Analysis
Autopsy specimens of normal human lung and liver were fixed with 10% formalin solution for about 2 weeks and then embedded in paraffin. The sections were pretreated with 0.1% trypsin for 30 min at 37C and then probed with Pr19 by the biotin-streptavidin method as described (Hitotsumatsu et al. 1997). Specificity of the signal was confirmed by the observations that the signal was not elicited by treatment without the primary antibody and that adding an excess amount of pyridylethylated, but not native, Hakata antigen diminished the signal. Anti-Sp-A antibody and anti-CC10 antibody were kindly provided by Dr. S. Hashimoto (Kyushu University).
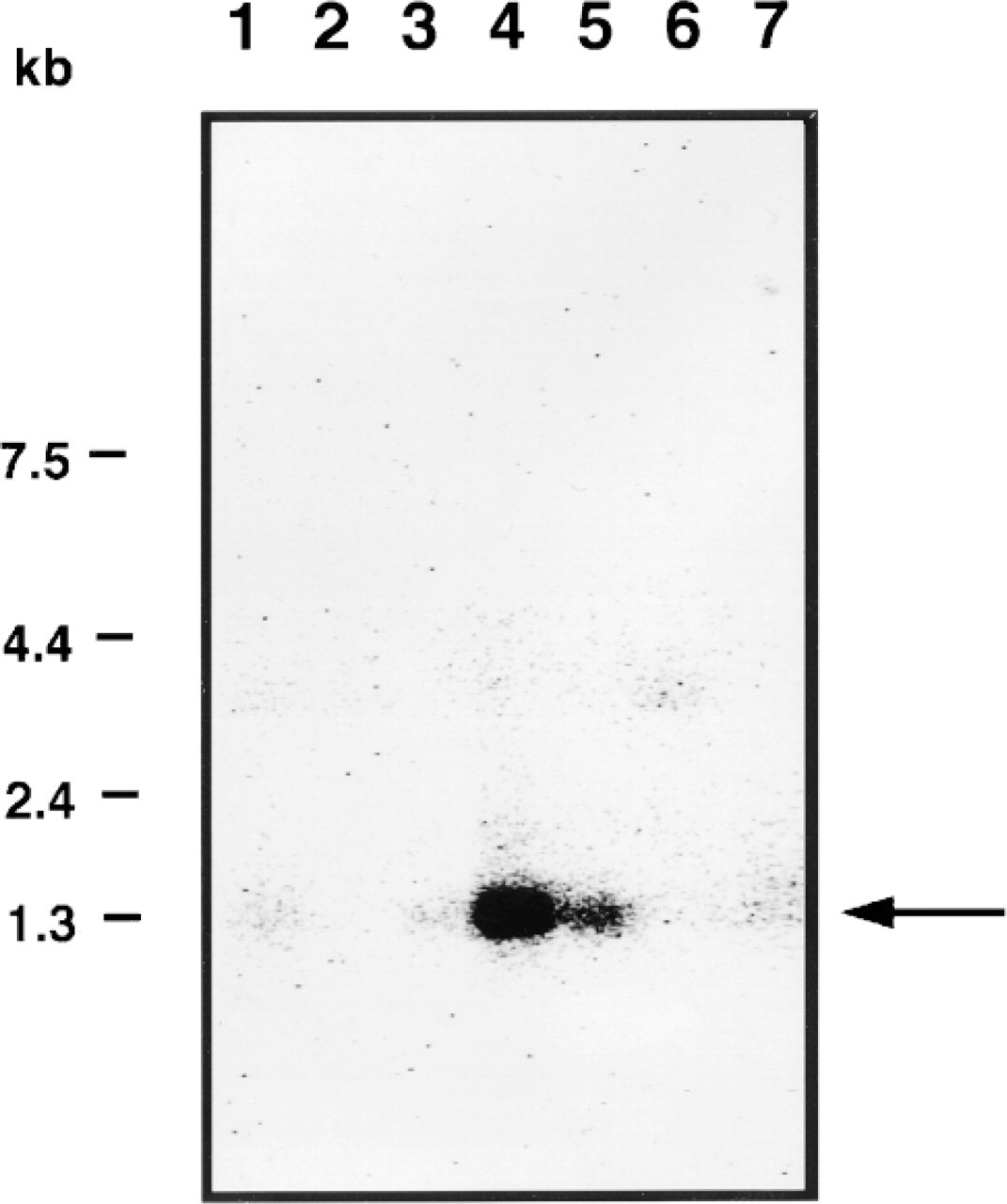
Northern blotting of Hakata antigen. A membrane filter containing mRNA from human tissues was hybridized with 32 P-labeled Hakata antigen cDNA fragment (nucleotides 448-864). mRNAs are derived from 1, heart; 2, brain; 3, placenta; 4, lung; 5, liver; 6, skeletal muscle; 7, kidney. Size markers are indicated at left. Arrow indicates the band of Hakata antigen.
Results
Expression of Hakata Antigen mRNA
To determine which tissues express Hakata antigen, we first performed Northern blotting using a complementary oligonucleotide to Hakata antigen (nucleic acids 448-864 from the translation starting site) as a probe. This oligonucleotide hybridized mRNAs derived from the lung and liver (Figure 1). No band was detected in mRNAs of spleen, thymus, prostate, testis, ovary, small intestine, colon, peripheral blood leukocytes, heart, brain, placenta, skeletal muscle, or kidney (Figure 1; data not shown). The sizes of the bands were about 1.3
Western Blotting of Hakata Antigen in Lung and Liver
To confirm that the lung and liver express Hakata antigen protein as well as mRNA, we next carried out Western blotting of Hakata antigen using total homogenates of lung and liver. Using the polyclonal antibody against Hakata antigen, the homogenates of both lung and liver gave rise to sharp bands whose sizes were identical to that of Hakata antigen purified from serum, although some bands of higher molecular weight were also detected in the case of the liver (Figure 2). It is unlikely that these bands are polymeric forms because Hakata antigen exists as a monomer under reducing conditions. These results suggest that the lung and liver are rich in Hakata antigen protein, whose molecular size is almost the same as that in serum.
Generation of a Monoclonal Antibody Against Hakata Antigen (Pr19)
We have previously shown that Hakata antigen exists as a polymer consisting of disulfide bonds in serum and that polyclonal rabbit antiserum against Hakata antigen recognizes both monomeric and polymeric Hakata antigens (Yae et al. 1991). Because the polyclonal antibody caused nonspecific background staining in immunohistochemistry (not shown), we attempted to generate a monoclonal antibody against Hakata antigen for immunohistochemistry. One hybridoma clone (Pr19) produced an antibody that specifically recognized monomeric Hakata antigen but not polymeric Hakata antigen (Figure 3). We applied this antibody in the following immunohistochemical analyses.
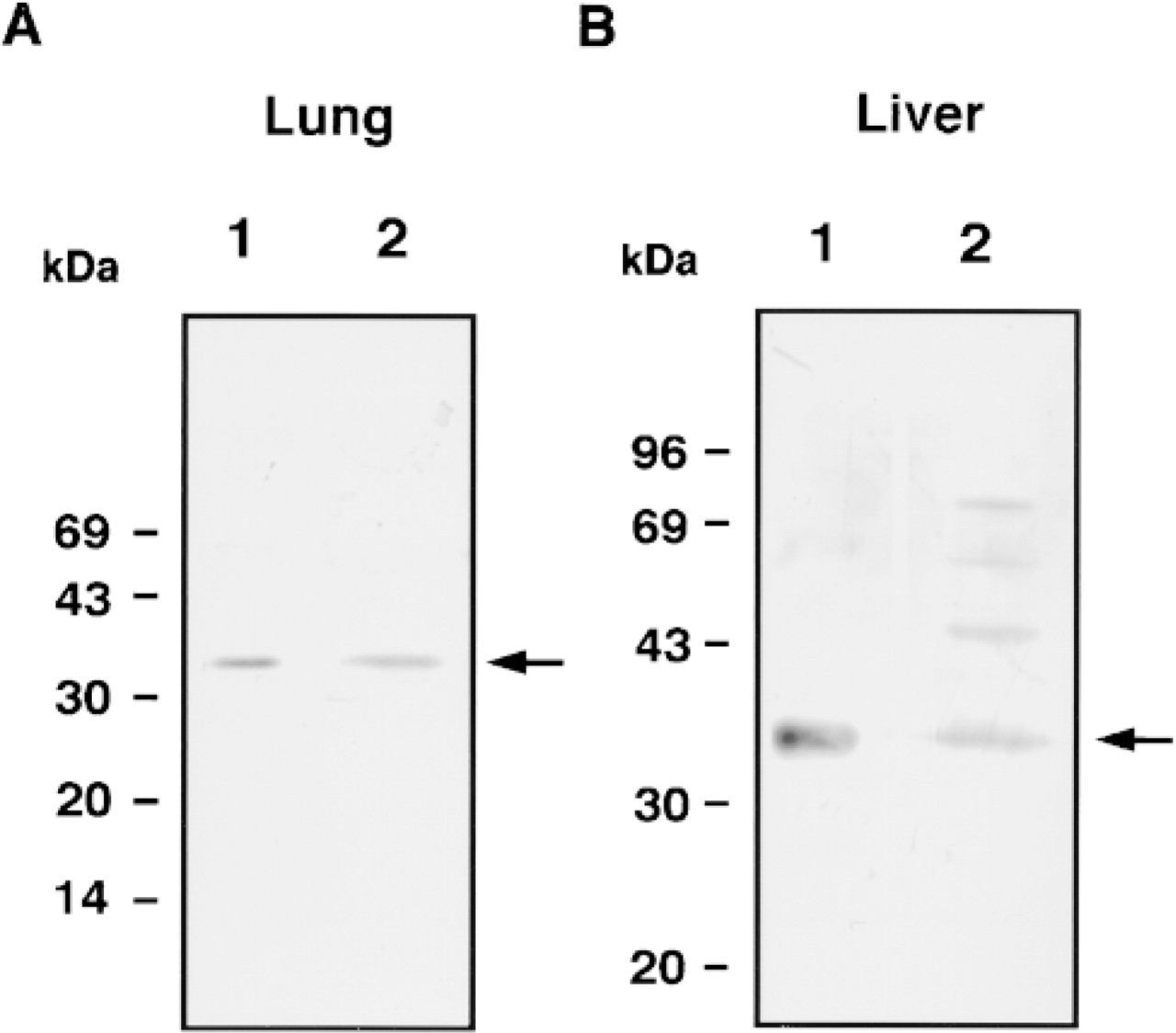
Western blotting of Hakata antigen in lung and liver. Ten ng (
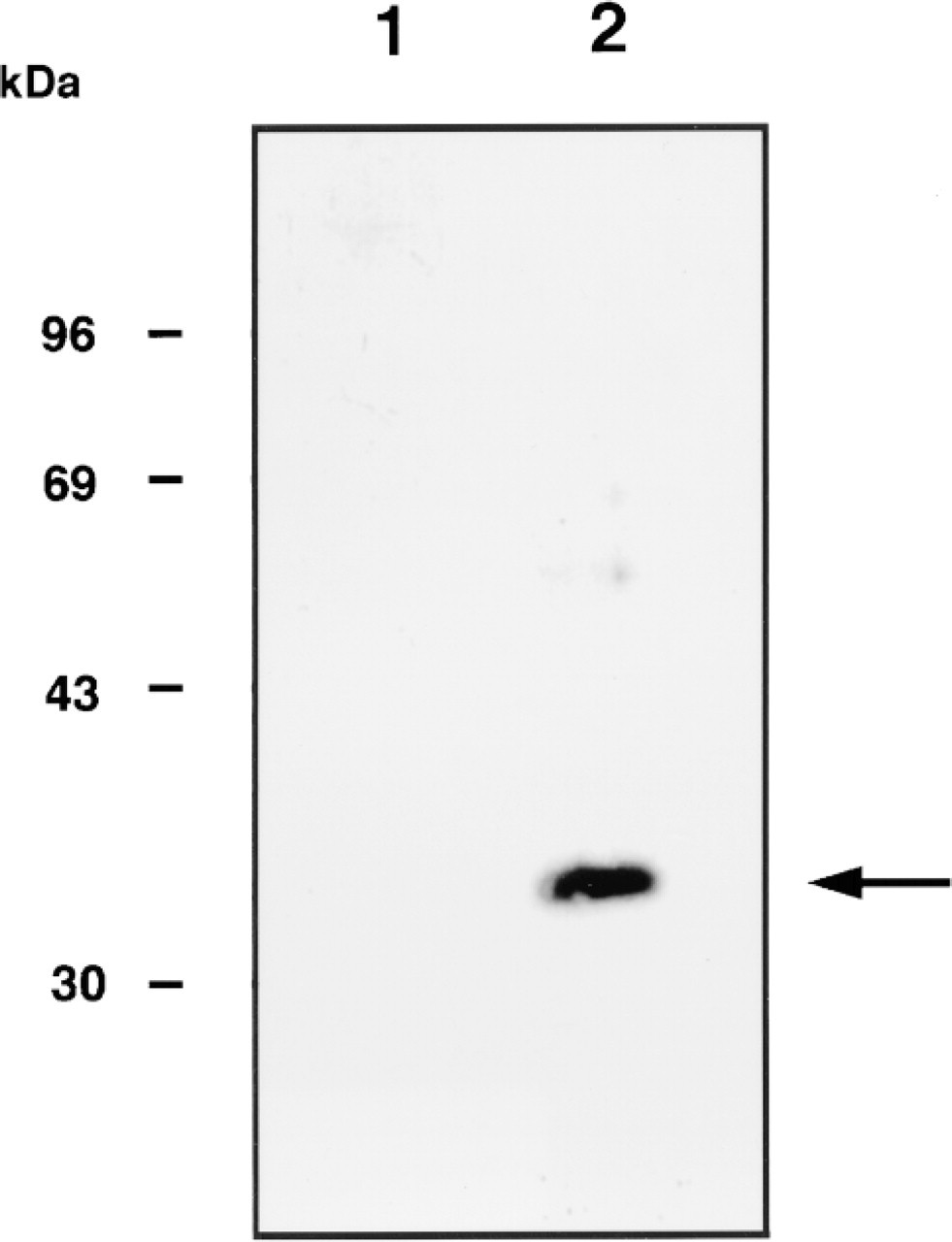
Western blotting of Hakata antigen by Pr19. One μg of purified Hakata antigen was probed with a monoclonal antibody against Hakata antigen (Pr19) in either nonreducing (Lane 1) or reducing (Lane 2) conditions. Molecular size markers are indicated at left. Arrow indicates the band of monomeric Hakata antigen (35 kD).
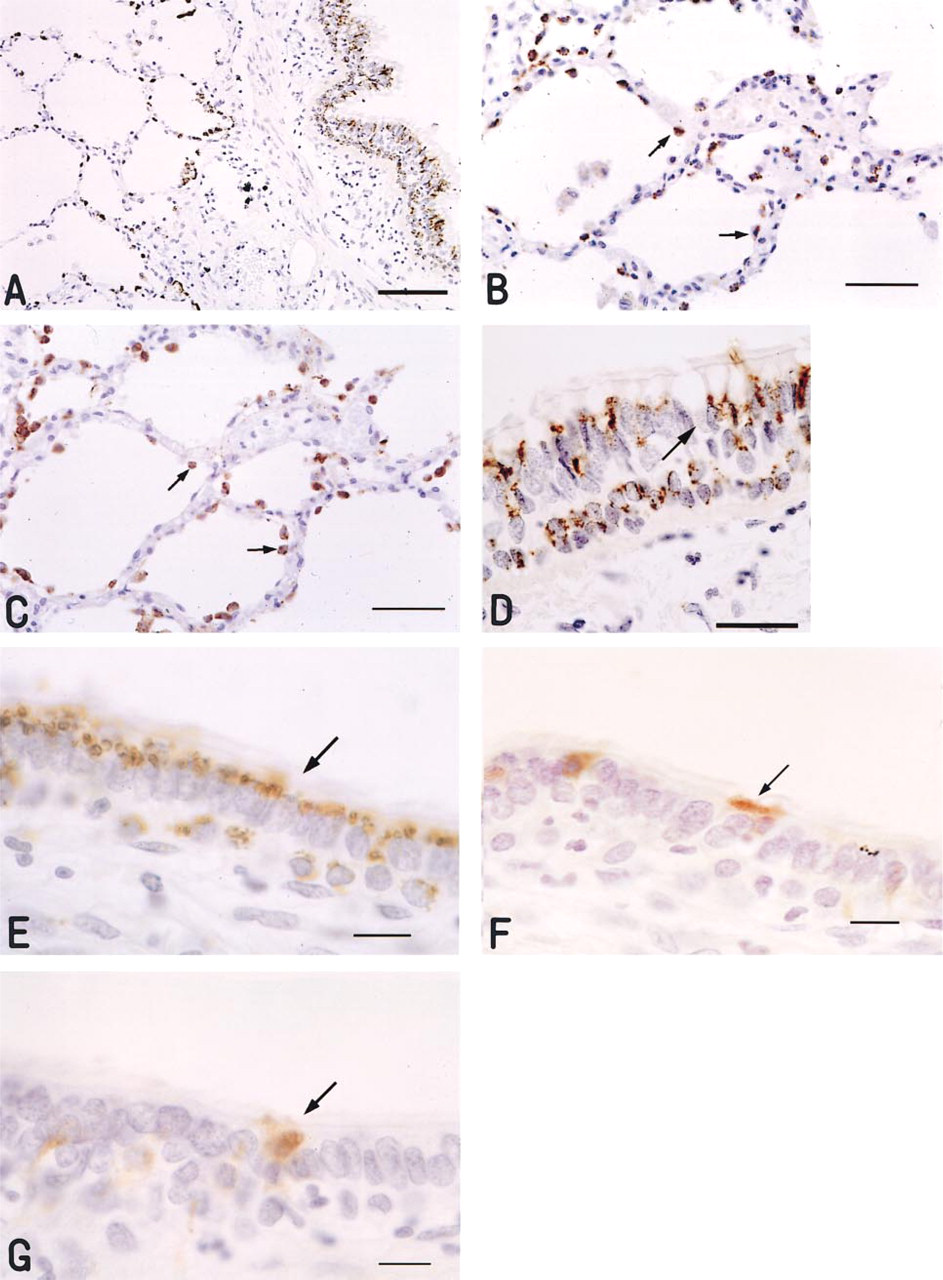
Immunoperoxidase staining of human lung probed with Pr19. (
Expression of Hakata Antigen in Lung
To localize Hakata antigen protein expressed in human lung, we carried out immunohistochemical analyses using Pr19. In the alveolar region, Type II alveolar epithelial cells, characterized by their round shape and plump cytoplasm, showed strong cytoplasmic staining with Pr19 (Figures 4A and 4B), whereas Type I alveolar epithelial cells, alveolar macrophages, and endothelial cells were not stained. To confirm that Type II alveolar epithelial cells were stained by Pr19, we stained two sequential sections with either Pr19 (Figure 4B) or anti-SP-A antibody (Figure 4C) because SP-A is known to be secreted from Type II alveolar epithelial cells (Hawgood and Shiffer 1991; Wright and Dobbs 1991). In these sections, both antibodies recognized Type II alveolar epithelial cells. In the bronchus and bronchioles, ciliated columnar epithelial cells were mainly stained by Pr19 (Figures 4A and 4D). Reserve cells of the epithelium located in the basal membrane were also strongly stained. These reaction products showed fine granular deposits in their cytoplasm; however, goblet cells were negative for Pr19 staining. To confirm that ciliated epithelial cells were mainly stained by Pr19, we stained sequential sections with either Pr19 (Figure 4E) or anti-SP-A antibody (Figure 4 F) or with anti-CC10 antibody (Figure 4G), the latter two of which are known to recognize Clara cells (Xu et al. 1998). Consequently, Pr19 gave rise to positive staining mainly in ciliated epithelial cells, whereas anti-SP-A antibody and anti-CC10 antibody did so in nonciliated epithelial cells (Clara cells). The staining pattern of Pr19 was the same in lung tissue derived from six different individuals (not shown). The results suggest that the monomeric Hakata antigen recognized by Pr19 was produced specifically by Type II alveolar epithelial cells and ciliated bronchial epithelial cells in human lung.
To clarify whether Hakata antigen is secreted into bronchus/alveolus, we subjected BAL fluids to Western blotting of Hakata antigen. In nonreducing conditions, a polymeric Hakata antigen with the same mobility as that in serum was detected in all BAL fluids from eight donors (Figure 5A), whereas in reducing conditions these polymeric Hakata antigens in BAL fluid dissociated to monomeric ones (35 kD; Figure 5B). These results demonstrated that the Hakata antigen generated in bronchial epithelial cells and Type II alveolar epithelial cells in human lung is secreted as a polymer into bronchus/alveolus.
Expression of Hakata Antigen in Liver
In liver, hepatocytes showed diffuse and weak staining of the cytoplasm by Pr19 (Figures 6A and 6B), and bile duct epithelial cells were strongly stained (Figures 6A and 6C). Kupffer cells, sinusoidal capillaries around hepatocytes, and Glisson's sheath around bile ducts were not stained. These results demonstrated that monomeric Hakata antigen protein is produced in both hepatocytes and bile duct epithelial cells, but to a much greater extent in the latter. The staining pattern was the same in liver tissue derived from seven different individuals (not shown).
To clarify whether Hakata antigen is secreted into bile ducts, we subjected bile samples to Western blotting. In nonreducing conditions, polymeric Hakata antigen protein, which has the same mobility as that in serum, was detected in all samples, whereas in reducing conditions only monomeric Hakata antigen was detected in bile as in BAL fluid (Figure 7). Although the intensity of the bands was very weak in Lanes 3 and 7, longer exposure of the film gave rise to definite bands. Therefore, Hakata antigen, which is formed in bile duct epithelial cells, is secreted as a polymer into bile ducts.
Discussion
It has recently been shown that Hakata antigen is a new member of the ficolin/opsonin p35 family by cloning of its cDNA and by functional analyses (Sugimoto et al. 1998). Hakata antigen shows overall homologies of 48% with both human ficolin-1 and opsonin p35, higher in the collagen-like domain (48%, 54%) and in the fibrinogen-like domain (52%, 53%), respectively (Sugimoto et al. 1998). Although animal lectins are categorized into several groups (Gabius 1997), it is believed that the ficolin/opsonin p35 family is distinct from the others. However, the physiological roles of these members await elucidation. In this study we examined the tissue distribution of Hakata antigen to obtain clues about its physiological role. Taking together the results of Northern blotting, Western blotting, and immunohistochemical analyses of lung and liver, we have concluded that Hakata antigen is a novel lectin protein secreted into bronchus/ alveolus and bile duct, extending previous reports that it is a novel serum protein (Yae et al. 1991; Sugimoto et al. 1998). These results indicate that Hakata antigen may play a role not only in serum but also in bronchus/alveolus and bile duct under physiological conditions. Among the other members of the ficolin/ opsonin p35 family, it has been shown that mRNA of human ficolin-1 is expressed in peripheral blood leukocytes, spleen, lung, placenta, and thymus (Harumiya et al. 1996; Lu et al. 1996), whereas that of opsonin p35 is expressed in liver, lung, and placenta (Matsushita et al. 1996), although it is not known which cells in these organs produce these molecules. The finding that mRNAs of both molecules are expressed in either lung or liver, like Hakata antigen, raises the possibility that all members of the ficolin/ opsonin p35 family are secreted into either bronchus/ alveolus or bile and may play a similar role in these areas. On the other hand, the notion that peripheral blood leukocytes are a major source of human ficolin-1 (Harumiya et al. 1996; Lu et al. 1996), whereas mRNA of Hakata antigen was not detected in peripheral blood leukocytes, suggests that these three molecules play distinct roles in different areas of the body.
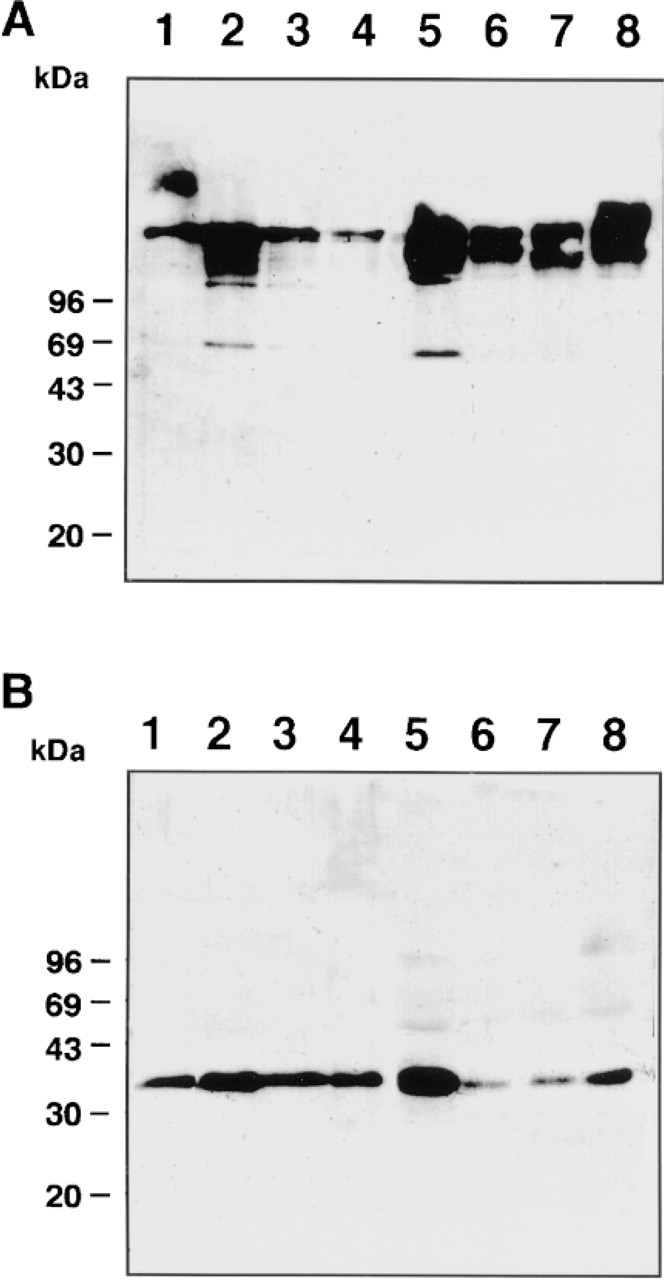
Western blotting of Hakata antigen in BAL fluid. Thirty ng of purified Hakata antigen (Lane 1,
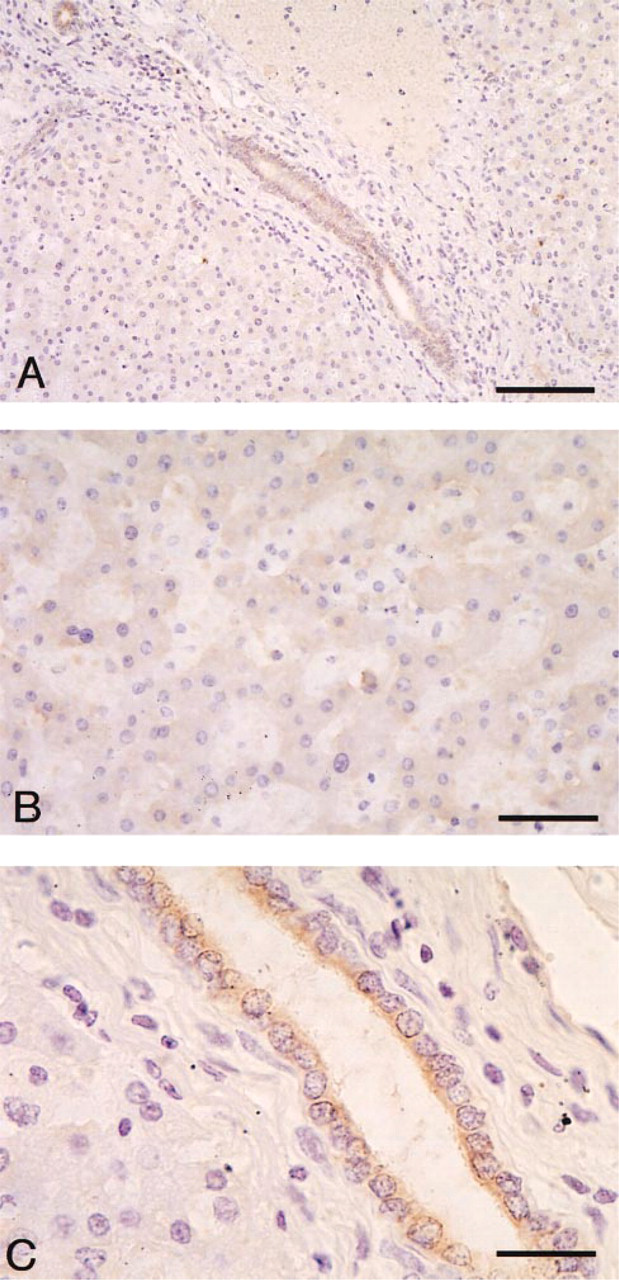
Immunoperoxidase staining of human liver probed with Pr19. (
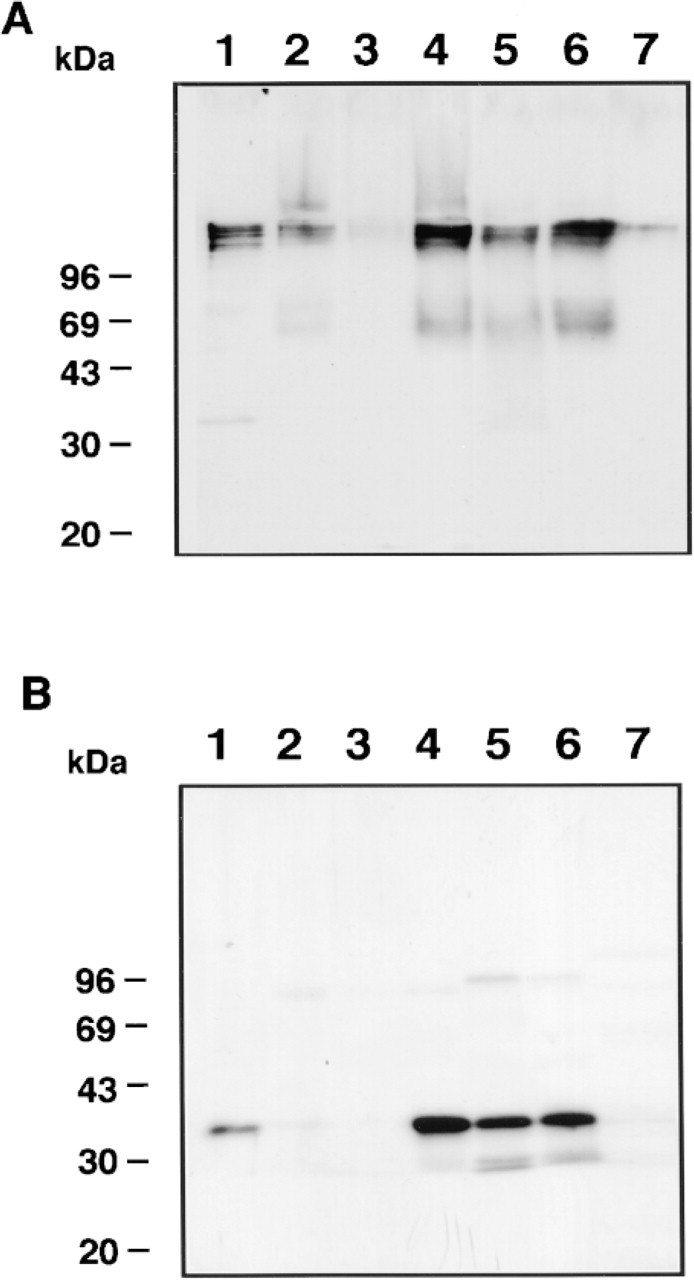
Western blotting of Hakata antigen in bile. Twenty-five ng of purified Hakata antigen (Lane 1,
We have demonstrated in this study that Hakata antigen as well as SPs was secreted into bronchus and alveolus. SP-A and SP-D are known to be water-soluble and to sustain lectin activity (Kuroki and Voelker 1994). Although these two SPs belong to the C-type lectin superfamily, whereas Hakata antigen does not, all of these molecules share common characteristics: (a) they are secreted by Type II alveolar epithelial cells; (b) collagen-like domains exist in the middle of the molecules; (c) these proteins form polymers consisting of octodecamers (SP-A, Hakata antigen) or a dodecamer (SP-D); and (d) these proteins can bind to carbohydrates (Kuroki and Voelker 1994). The difference between Hakata antigen and SP-A is that, in the bronchus and bronchioles, Hakata antigen is mainly produced by ciliated epithelial cells, whereas SP-A is produced by Clara cells. It has been shown that SP-A and SP-D play two important roles in bronchus and alveolus. One is the pulmonary surfactant system; these SPs bind to phospholipid to generate the surfactant monolayer, lowering surface tension at the air-liquid interface (Hawgood and Shiffer 1991; Weaver and Whitsett 1991; Wright and Dobbs 1991). The other is that these molecules modulate host defense functions in the lung by stimulating phagocytosis and chemotaxis of macrophages and by binding to a variety of microorganisms (Tenner et al. 1989; Kuan et al. 1992; van Iwaarden et al. 1991,1992; Zimmerman et al. 1992; Wright and Youmans 1993). On the basis of analogies in the distribution, structure, and function of these SPs and Hakata antigen, it is assumed that Hakata antigen plays roles similar to these SPs. Further studies aimed at this point are needed.
In previous studies, it has been verified that Hakata antigen is a novel serum protein with a range of 7-23 μg/ml (Yae et al. 1991). This value means that Hakata antigen is a relatively abundant protein, whose concentration is equivalent to those of some complement proteins, such as C1q, C2, C5-9 (20-80 μ/ml). The serum concentration of human ficolin-1 has been reported to be at the same level as that of Hakata antigen (1-7 μ/ml, average 4.13 μ/ml; Le et al. 1998). It has been unclear which tissue is the main source of Hakata antigen in serum in the body. However, our present findings suggest that Hakata antigen in serum is produced in hepatocytes. This notion is compatible with the finding that the serum level of Hakata antigen declines in patients with liver cirrhosis (Fukutomi et al. 1996).
Bile duct epithelial cells are known to modify hepatic bile by both secretory and absorptive processes, producing as much as 40% of the daily output of bile (Boyer 1996). We have demonstrated here that Hakata antigen was produced by bile duct epithelial cells and secreted into the bile duct. To our knowledge, no lectin protein, other than Hakata antigen, has been shown to exist in bile. Its function remains unclear. It is well known that bile contains various kinds of glycoproteins that bind to lectins. These glycoproteins include mucin glycoproteins, which are the most abundant, and nonmucin glycoproteins, such as aminopeptidase N, α2-macroglobulin, hemopexin, immunoglobulin heavy chains, and the β-chain of haptoglobulin (Klinkspoor et al. 1994; Lipsett et al. 1994; Offner et al. 1994; van Wijland et al. 1994; Busch et al. 1995). These glycoproteins play important roles in protection against the detergent action of bile salts under physiological conditions, and in cholesterol crystallization under pathological conditions (Klinkspoor et al. 1994; Lipsett et al. 1994; Offner et al. 1994; van Wijland et al. 1994; Busch et al. 1995). Likewise, Hakata antigen may associate with these glycoproteins in bile and may affect the activities of these glycoproteins. Although the present limited analyses of inpatient bile did not show any significant difference in the amount of Hakata antigen contained (Figure 7), further studies focused on the association between Hakata antigen and these glycoproteins under either physiological or pathological conditions would be useful.
Footnotes
Acknowledgments
Supported in part by grants-in-aid for Scientific Research from the Ministry of Education, Science, Sports and Culture of Japan.
Dr Yae passed away in August of 1996. This work is dedicated to Dr Yae's collaborative efforts in scientific research in a career spanning nearly a quarter of a century.
We thank Ms K. Hatanaka for skillful help and Dr S. Hashimoto for providing antibodies against SP-A and CC10. We also thank Dr Dovie R. Wylie for critical review of this manuscript.