
Abstract
After being synthesized in the cytoplasm of axon terminals, acetylcholine is packaged into synaptic vesicles by a proton-dependent transporter, vesicular acetylcholine transporter (VAChT). Localization of VAChT is restricted to cholinergic neurons, especially their terminals. We used an anti-VAChT antibody from INCSTAR to localize cholinergic terminals in the rat cochlear nucleus (CN), an important brainstem auditory center. VAChT immunoreactivity in the rat CN appears as labeled puncta and a few connecting fibers. In ventral CN (VCN), VAChT-labeled puncta are closely associated with somatic profiles of medium to large neurons. In and near the granular regions of VCN, VAChT-labeled puncta are more diffusely scattered. In the subpeduncular corner and the medial sheet, some VAChT-labeled fibers are seen in connection with especially prominent VAChT-labeled puncta. In dorsal CN (DCN), VAChT-labeled puncta show no clear association with somata and are found in all layers. Ultrastructurally, VAChT labeling is seen in the cytoplasm and is associated with synaptic vesicle membrane of terminals with small round vesicles. Such VAChT-labeled terminals synapse with cell bodies and dendrites in the CN.
Acetylcholine (ACh) metabolism in a cholinergic terminal and synapse involves several processes: synthesis, packaging, release, degradation, and recycling (Figure 1). Some of the key elements associated with these processes have been used as markers to identify cholinergic terminals. For example, choline acetyltransferase (ChAT, the synthetase for ACh) has long been studied as an indicator of cholinergic structures (Hebb 1957; McCaman and Dewhurst 1970; Fonnum et al. 1973; Wainer et al. 1984). On the other hand, because of its association with cholinergic synapses, including pre- and postsynaptic sites and synaptic clefts, acetylcholinesterase (AChE, the degradative enzyme for ACh) has also been widely studied (Koelle 1963; Shute and Lewis 1965; Silver 1967; Godfrey et al. 1977). Vesicular ACh transporter (VAChT), a proton-dependent transporter that facilitates packaging of ACh into synaptic vesicles after its synthesis in terminal cytoplasm, has recently emerged as a useful marker for identifying cholinergic terminals. Successful production of a specific antibody against VAChT (Gilmore et al. 1996) has provided a valuable histological tool for studying the distribution of cholinergic terminals in the central nervous system.
In the cochlear nucleus (CN), cholinergic innervation has been demonstrated by the presence of cholinergic/cholinoceptive markers such as choline acetyltransferase (ChAT), acetylcholinesterase (AChE), and ACh receptors (AChR). A lesion of virtually all centrifugal pathways to the CN depleted approximately 85% of ChAT activity in the rat CN (Godfrey et al. 1983), indicating that the CN is basically a cholinoceptive region rather than a cholinergic projection structure. This has been confirmed by the demonstration of ChAT-immunoreactive puncta, presumably cholinergic terminals, in the CN (Henderson and Sherriff 1991; Vetter et al. 1993; Godfrey 1993) and by the demonstration of muscarinic AChRs (mAChRs) in the CN with a distribution pattern similar to that of ChAT (Yao and Godfrey 1995). However, many questions remain about the exact character and function of cholinergic transmission in the CN (Yao and Godfrey 1996; Happe and Morley 1998). In this study, cholinergic terminals in the rat CN were localized by VAChT immunohistochemistry, using an antibody specific for VAChT, which has been found to be more sensitive than most ChAT antibodies for visualization of cholinergic terminals (Gilmore et al. 1996).
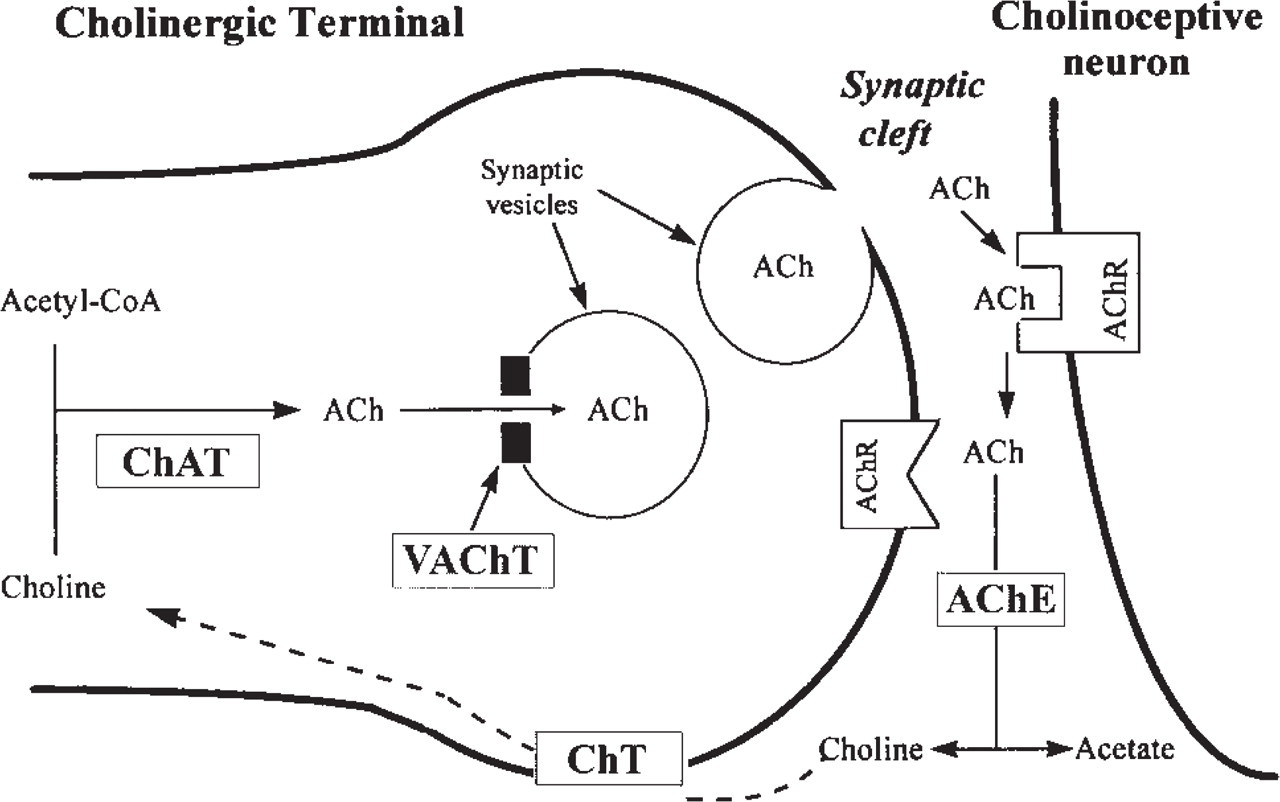
Schematic representation of acetylcholine (ACh) metabolism in a cholinergic terminal and synaptic cleft. ACh synthesis is catalyzed by choline acetyltransferase (ChAT). Packaging of ACh into synaptic vesicles is mediated by vesicular ACh transporter (VAChT). ACh in the synaptic vesicles is released, in response to depolarization of the presynaptic membrane, into the synaptic cleft, where it binds to post- and/or presynaptic ACh receptors (AChR), eliciting cholinergic effects. ACh is degraded by acetylcholinesterase (AChE) to acetate and choline. Choline is taken up into the cholinergic terminal, via a choline transporter (ChT), for recycling into ACh synthesis.
Materials and Methods
Our immunohistochemical procedure was similar to that employed in our previous studies and those of others (Gilmore et al. 1996; Yao and Godfrey 1998). In brief, young (250-350 g) male Sprague-Dawley rats were deeply anesthetized with sodium pentobarbital IP and perfusion-fixed with 3% paraformaldehyde in PBS, pH 7.4, for light microscopic (LM) immunohistochemistry. Glutaraldehyde at 0.2% was added to the fixative for electron microscopic (EM) immunocytochemistry. For LM, brain blocks were cryoprotected with 30% sucrose overnight for 2 days before 30-μm-thick transverse sections were cut in a cryostat. For EM, 50-μm-thick sections were cut with a vibratome and collected in PBS. Free-floating sections were blocked with 5% normal rabbit serum followed by incubation with anti-VAChT (INCSTAR; Stillwater, MN) at 1:1000 dilution at 4C for 20 hr. Chromogenic detection used the Vectastain ABC/DAB system (Vector; Burlingame, CA). For LM, sections were dehydrated and mounted on precoated slides. For EM, sections were dehydrated with graded ethanols and absolute acetone, osmicated with 1% OsO4, infiltrated, and flat-embedded with Spurr's resin. Thin sections (80-100 nm thick) were cut through the CN, collected/mounted on copper grids, and poststained with 1% uranyl acetate and 1% lead citrate.
Light microscopic examinations and photographic demonstrations of VAChT immunolabeling within the brainstem and the CN were conducted with a microscope (Leitz Aristoplan 2; Oberkochen, Germany). An imaging program, Neurolucida 2.1 (MicroBrightField; Colchester, VT), was used to measure the relative density of VAChT labeling for comparison among different CN regions, using the same procedures as described previously (Yao and Godfrey 1995). Such measurements have been found to correlate with the amount of antigen (Yao and Godfrey 1995; Huang et al. 1996). In addition, the perimeters of labeled puncta were measured using Neurolucida with a X100 objective lens (oil immersion). For each region, measurements were made for two sections of each CN of three rats with optimal contrast between labeling and background (12 sections total), in sections with large representations of the subdivisions (AVCN, PVCN, IN, and DCN). Comparisons among regions were evaluated statistically using analysis of variance combined with t-tests.
Subcellular localization of VAChT labeling was observed and photographed using an MC 10 electron microscope (courtesy of the Department of Pathology, Medical College of Ohio).
Results
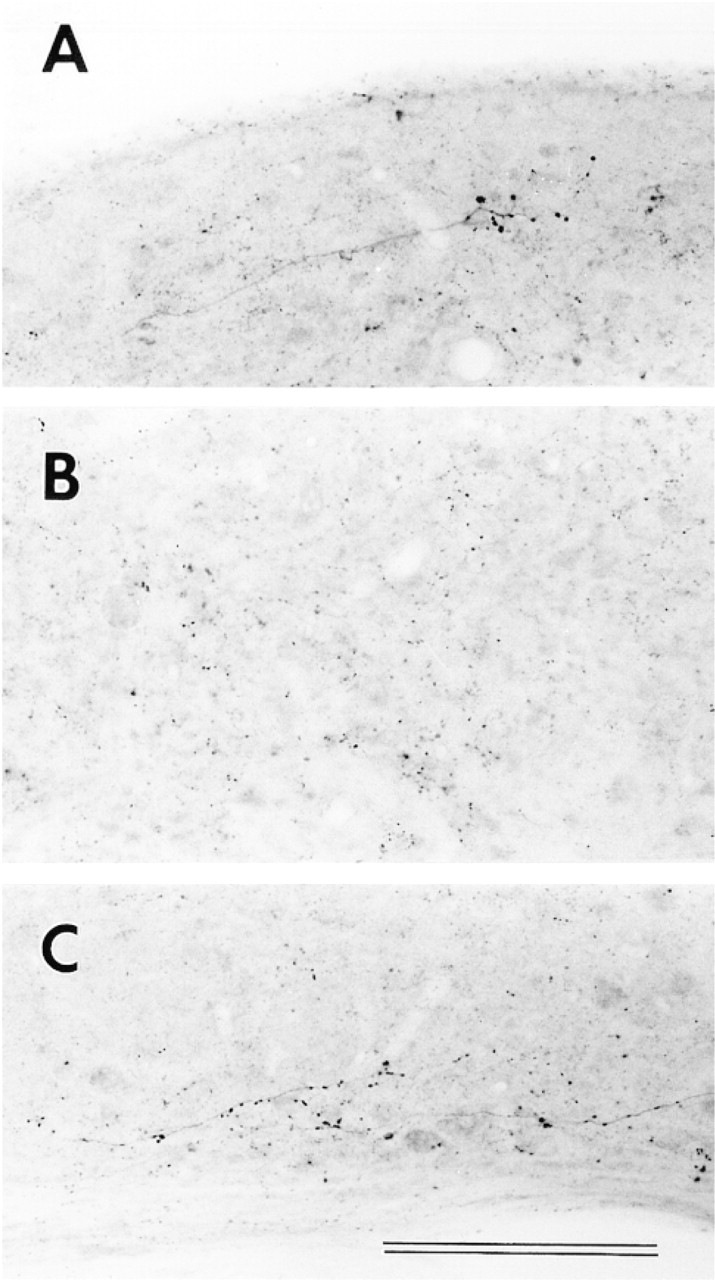
Our results for VAChT in the brainstem appear similar to those of Gilmore et al. (1996), with intense labeling in the FN and moderate labeling in the CN (Figure 2). Measurements were made of the densities of VAChT immunoreactivity across different CN sub-regions and the FN (Figure 3). Such measurements represent relative amounts of labeling, regardless of the nature of labeled structures (e.g., somata, axons, dendrites, or terminals), and allow objective quantitative comparisons. The VAChT labeling in the CN is about one third of that in the FN. Inside the CN, there is a tendency, although statistically not significant, towards denser labeling in the medial sheet and lesser labeling in the granular regions superficial to AVCN and PVCN. In DCN, labeling density is similar to that in VCN. Although differences across the layers are small, that between the fusiform soma and deep layers is statistically significant (p<0.02).
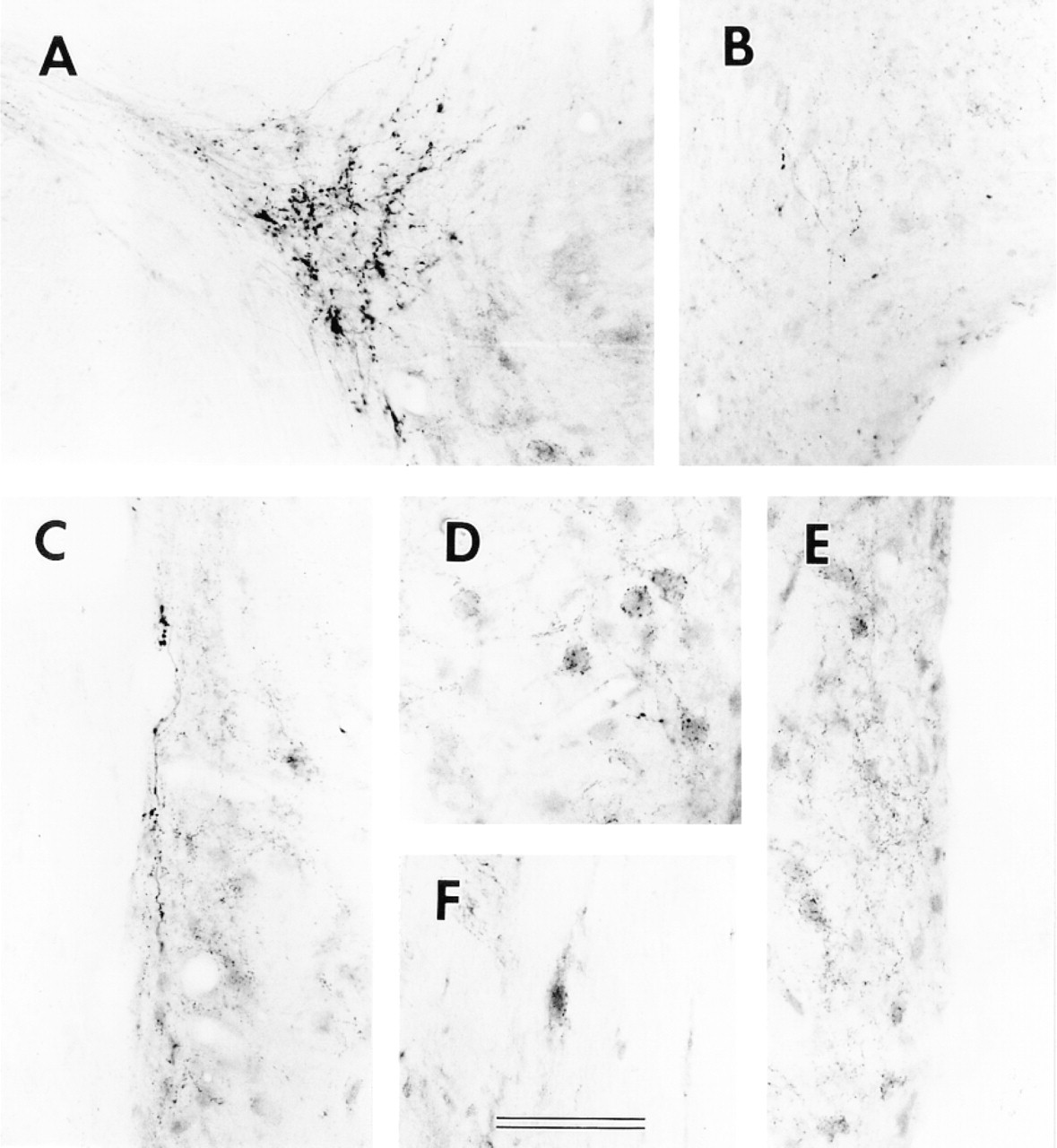
At higher magnification, VAChT labeling in all CN regions is in fibers and puncta but not in cell bodies. In both AVCN and PVCN (Figures 4 and 5), the VAChT in the subpeduncular corner (SPC) and medial sheet (MS) is associated with passing fibers having many prominent varicosities and puncta. Inside the core portions (magnocellular regions) of VCN, VAChT-labeled puncta appear to be associated with somata and their proximal dendrites. The somata apposed by VAChT puncta are almost all medium to large in size. In the superficial granular regions of VCN and the granular region between VCN and DCN, the VAChT-labeled puncta are sparsely scattered and less prominent (Figures 4C, 5B, and 5E). Although a few labeled fibers are present, no prominent fiber tract or bundle is labeled. There is also VAChT labeling, in the form of fibers and strings of puncta, in the neuropil of the VCN.
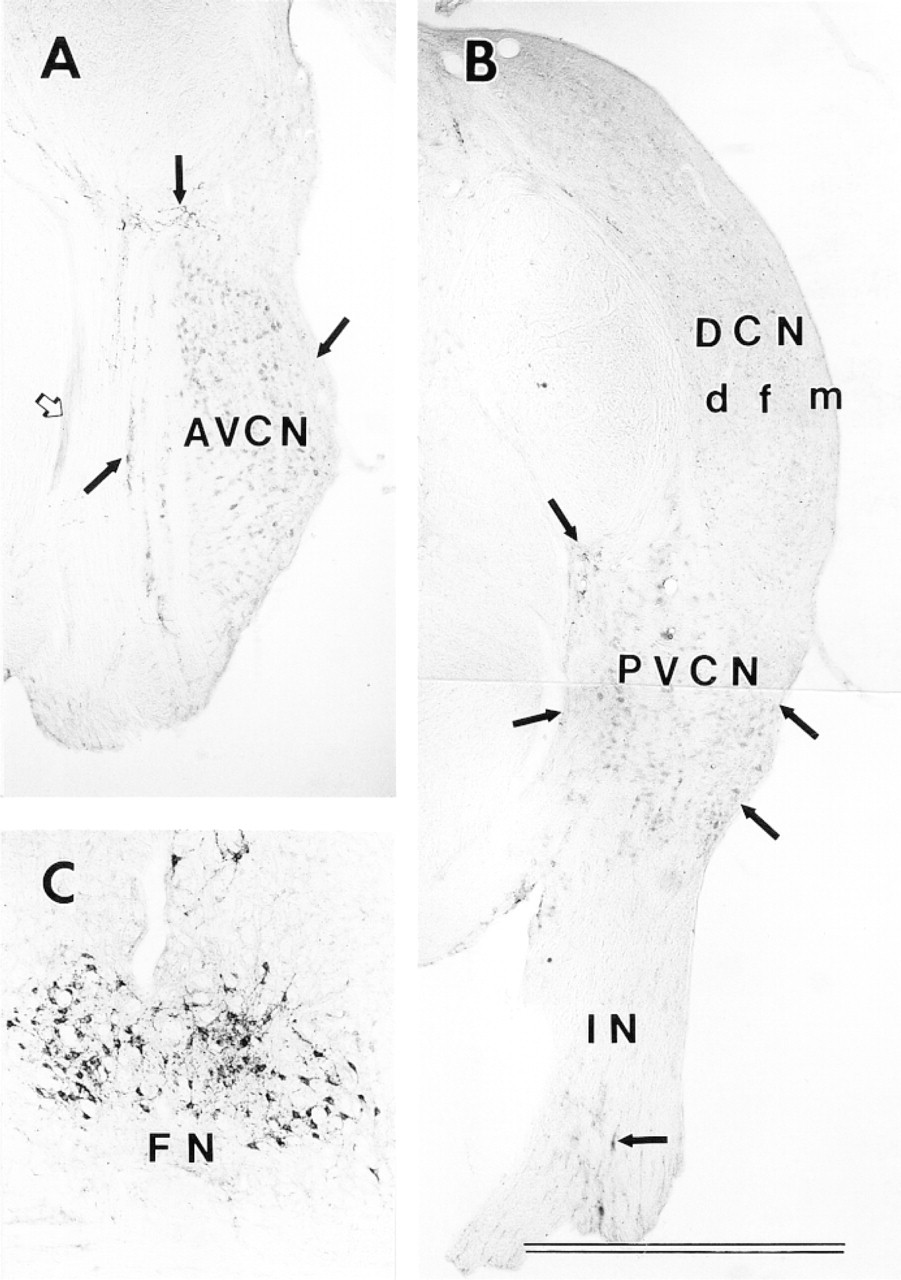
Immunolabeling for vesicular acetylcholine transporter (VAChT) in the cochlear nucleus (CN) and facial nucleus (FN). (
In the DCN, as in the VCN, VAChT immunoreactivity is associated with fibers and puncta (Figure 6). There are some prominent labeled fibers in the molecular and deep layers oriented parallel to the surface of the DCN (Figures 6A and 6C). In the fusiform soma layer, there are fewer labeled fibers but many labeled puncta which, unlike those in the VCN, are scattered without any clear association with somatic profiles (Figure 6B). There appears to be more labeling in the neuropil or background in the DCN than in the VCN.
There are noticeable variations in the sizes of VAChT-labeled puncta in different parts of the CN. The measured puncta sizes are larger in the medial sheets, the subpeduncular corners at the dorsal aspects of the VCN, and the DCN molecular and deep layers than in the core portions of the VCN, granular regions, and the DCN fusiform soma layer (Figure 7) (p<0.005 in all cases except for the comparison between DCN molecular layer and PVCN granular region, for which p<0.02). In addition, the shapes of labeled puncta vary among different CN locations. The VAChT-labeled puncta associated with somata are, on average, flatter, those associated with the subpeduncular corner and medial sheet fibers larger and round to oval, and those in the neuropil smaller and round.
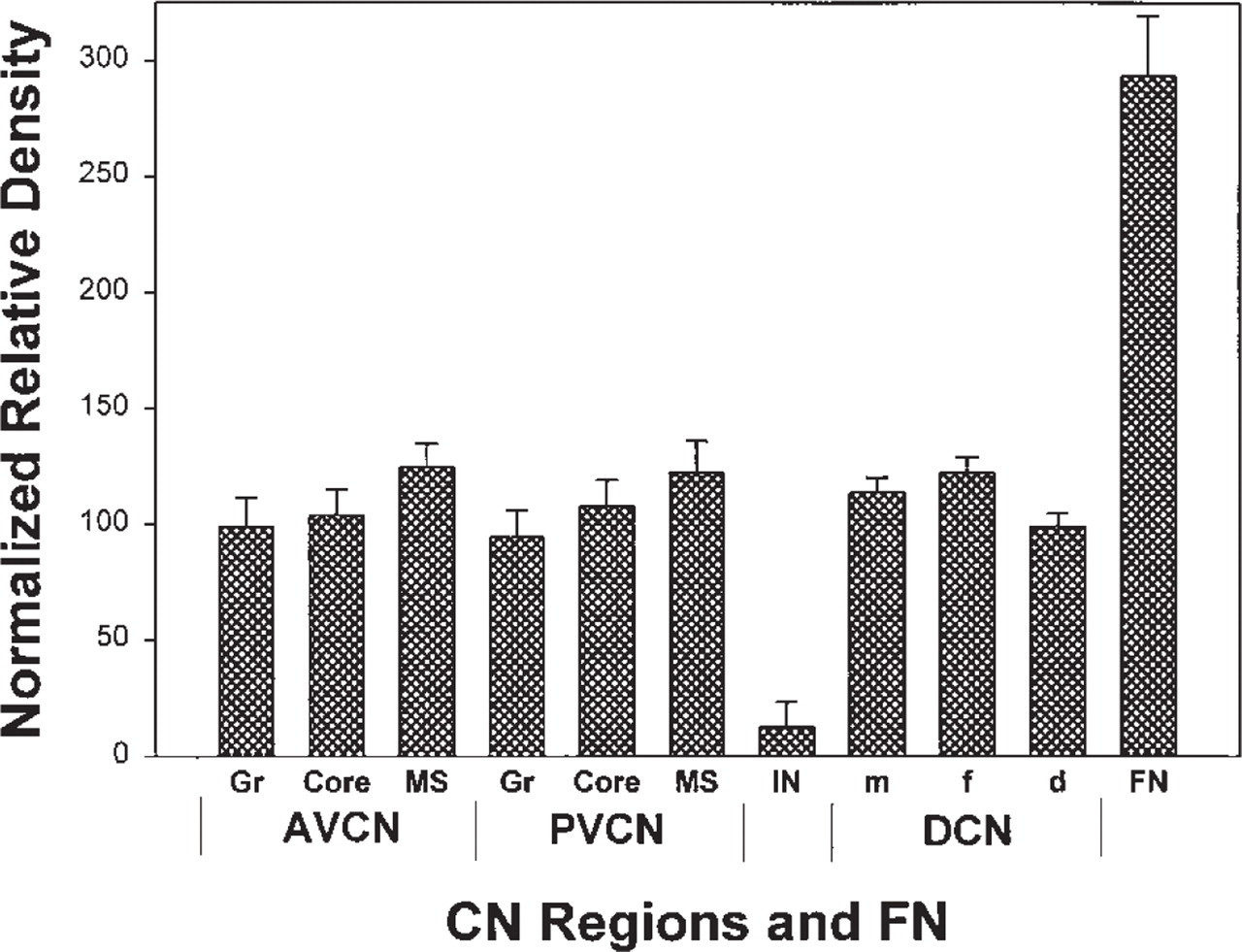
Densitometry of VAChT immunoreactivity in the rat CN. Labeling densities for identified CN regions were measured using Neurolucida 2.1 and corrected by subtracting measurements from the spinal trigeminal tract, a nearby noncholinergic structure, as an internal background control. A ratio (%) of each corrected density to the value for the whole CN was used to normalize for variations across different preparations.
The subcellular localization of VAChT immunoreactivity was examined by EM. Examples of VAChT-labeled puncta are shown in Figure 8. The VAChT-immunoreactive materials are closely associated with the synaptic vesicular membrane in the terminals. These VAChT-labeled terminals were found in all CN regions, forming axosomatic and axodendritic synapses. Compared to those of nonimmunoreactive terminals, all VAChT-labeled synaptic vesicles are of the small round type, corresponding to previous descriptions of those in cholinergic terminals of the CN (McDonald and Rasmussen 1971; Altschuler et al. 1993). Perimeters of VAChT-labeled synaptic vesicles are significantly smaller than those in terminals with large pleomorphic vesicles: mean ± SEM = 0.124 ± 0.001 μm for 132 measurements vs 0.165 ± 0.001 μm for 86 measurements, respectively (p<0.001).
Discussion
The VAChT immunohistochemical results in the brainstem in our preparations resemble those of Gilmore et al. (1996) at both light and electron microscopic levels. The close association of immunoreactive materials with the synaptic vesicle membrane, as seen in the EM preparations, is indicative of the specificity of the labeling. Therefore, the results for labeled puncta and fibers in this report can be considered representative of cholinergic terminals and axons, and they provide a map of cholinergic terminals in the rat CN.
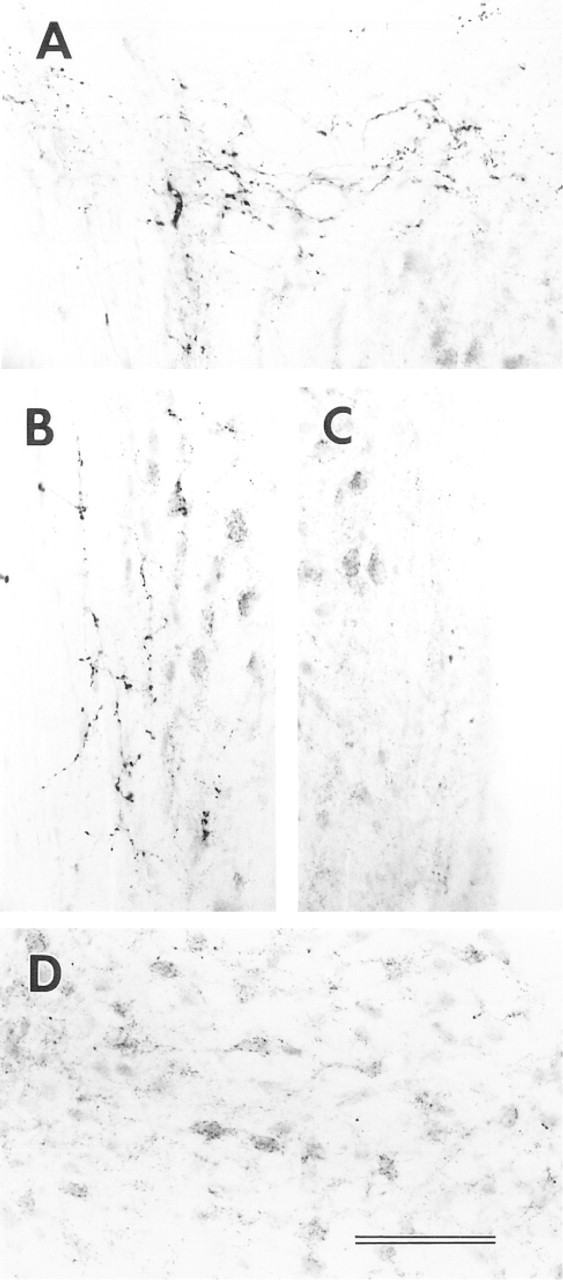
High magnification of VAChT immunolabeling in the AVCN. (
High magnification of VAChT immunolabeling in the PVCN and IN. (
The differences among VAChT-labeled puncta in different CN regions suggest a variety of configurations of cholinergic innervation in the CN. (a) In the core portions of the VCN, the association of most VAChT-labeled terminals with somata of medium to large neurons and their proximal dendrites suggests that most cholinergic transmission here is axosomatic.
In addition, apposition of many labeled puncta to each soma may suggest convergence of cholinergic input to these VCN neurons. There is evidence for both muscarinic and α7 nicotinic receptors on these VCN somata (Yao and Godfrey 1995; Happe and Morley 1998). (b) In the DCN fusiform soma layer and the granular regions superficial to the VCN, axodendritic cholinergic transmission may be predominant because most VAChT-labeled puncta in these locations are not closely associated with somata. (c) In the medial sheet, subpeduncular corner, and parts of the DCN molecular and deep layers, branching of individual fibers to provide multiple terminals, combined with relatively large terminal sizes, suggests that each cholinergic fiber may have a stronger influence here than elsewhere in the CN. The different patterns of cholinergic innervation in different CN regions may be associated with different origins (Godfrey et al. 1987), including intrinsic neurons (Godfrey 1993; Vetter et al. 1993), collaterals of the medial OCB (Benson and Brown 1990), and a population of smaller neurons of the ventral nucleus of the trapezoid body (VNTB) (Sherriff and Henderson 1994; Yao and Godfrey 1998). Alternatively, they may suggest subgroups within the VNTB population of smaller neurons, which appears to be the predominant source (Sherriff and Henderson 1994; Yao and Godfrey 1998).
High magnification of VAChT immunolabeling in DCN. (
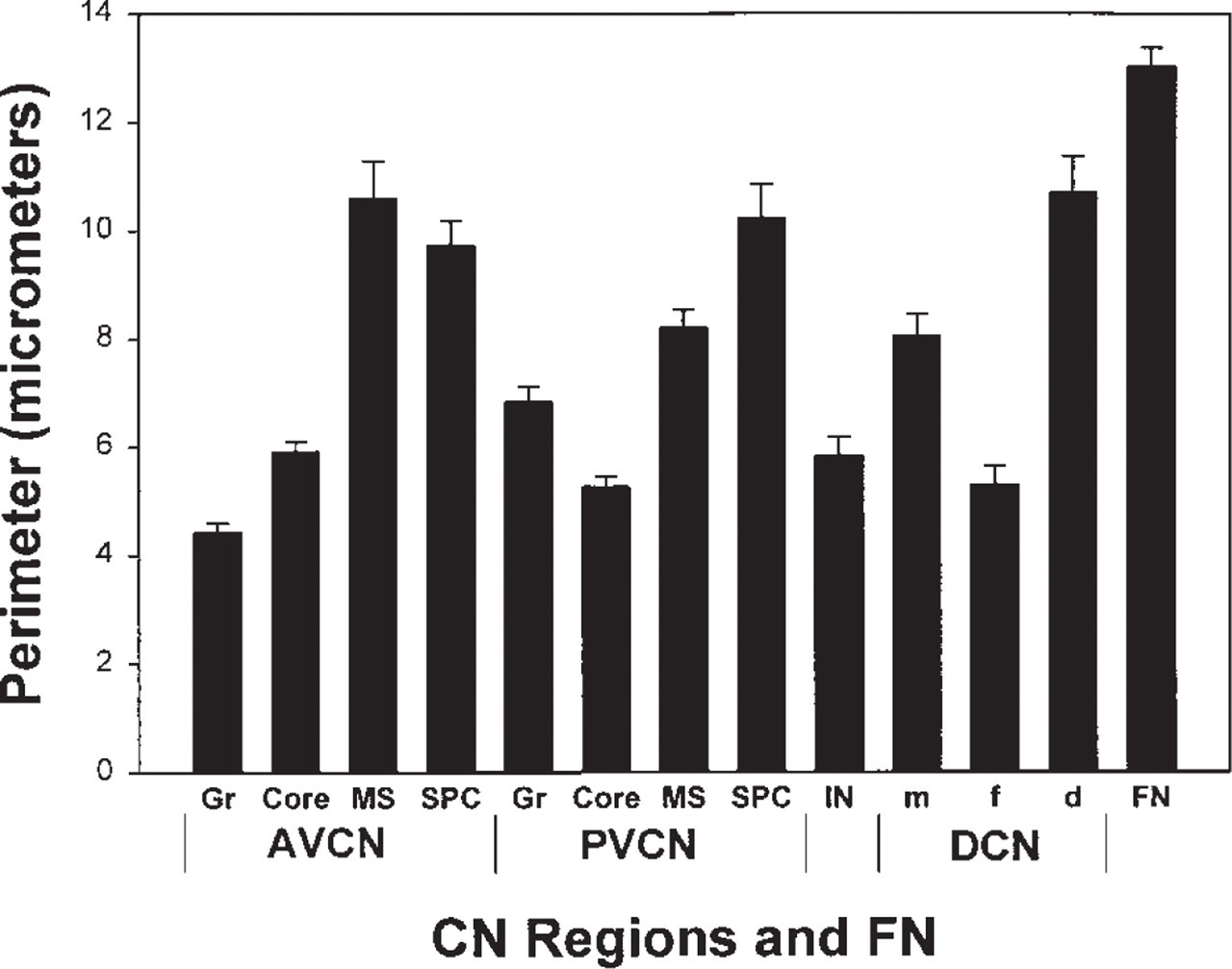
Sizes of VAChT-labeled puncta in rat CN. The mean perimeter (μm ± SEM) of VAChT-labeled puncta in each location was obtained by tracing their contours using Neurolucida 2.1. The numbers of puncta measured ranged from 27 to 45 and averaged 38.
The results for VAChT labeling are similar to those for muscarinic AChR and ChAT (Yao and Godfrey 1995). However, our results and those of Gilmore et al. (1996) suggest that VAChT and ChAT immunohistochemistries are not always well matched. For example, in fiber tracts such as the facial nerve root (not shown) and the OCB (Figure 2A), VAChT labeling is much less intense than that of ChAT (Yao and Godfrey 1995; Yao et al. 1996). Morphologies of labeled puncta appeared clearer with VAChT than with ChAT (Gilmore et al. 1996). These differences may be reflections of different metabolisms of the two proteins. Rapid axonal transport of VAChT may explain its low labeling densities in fiber tracts (Gilmore et al. 1996), and high concentrations of VAChT-labeled synaptic vesicles in terminals may result in dense punctate labeling (Figure 8).
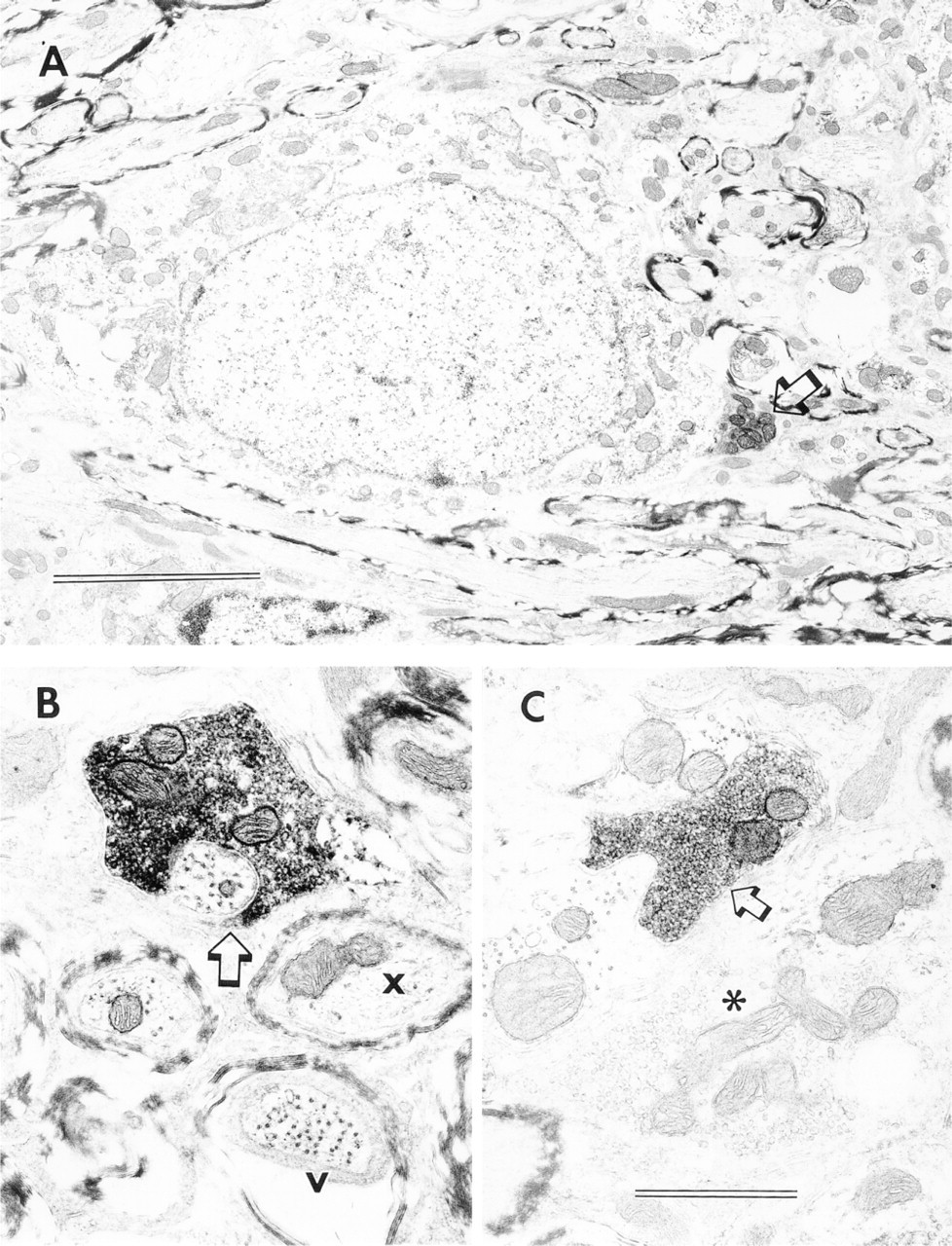
Subcellular localizations of VAChT in the CN. (
Presynaptic m2 AChRs have been previously found in rat CN and are believed to be localized to a subpopulation of cholinergic fibers and terminals (Levey et al. 1995; Yao et al. 1996). Comparison of current results for VAChT immunoreactivity with the previous m2 results reveals some agreements and disagreements in the rat CN. Both VAChT and m2 labeling, in the form of fibers and puncta, are prominent in the subpeduncular corner and the medial sheet of AVCN and PVCN, but there is no prominent VAChT labeling of the subgranular layer in AVCN or PVCN, as seen with m2 immunolabeling (Yao et al. 1996). Because VAChT is associated more with terminals than with fibers, this discrepancy fits with the impression that the m2 labeling in the subgranular layer is related to fibers more than to terminals (Yao et al. 1996). VAChT-labeled puncta diffusely arranged in and near the granular regions of the VCN may represent cholinergic terminals originating from incoming fibers that are not well labeled for VAChT.
There is less correlation between VAChT immunohistochemistry and AChE histochemistry, becausethere is no dense VAChT labeling to match the relatively dense labeling for AChE in the granular regions of VCN and the DCN fusiform soma layer (Yao et al. 1996).
In conclusion, we have found VAChT immunolabeling concentrated in terminals associated with membranes of small round vesicles. In the rat CN, VAChT-labeled cholinergic terminals vary in distribution pattern and size in different locations, suggesting possible variations in cholinergic influences among different CN regions.
Footnotes
Supported by NIH grant DC 00172.
We thank Dr William Gunning and Mr Edward Calomeni (Department of Pathology, Medical College of Ohio) for their generous provision of electron microscopy and photographic facilities. The care and use of animals were approved by the National Institutes of Health (NIDCD) and by the Medical College of Ohio Institutional Animal Care and Use Committee.