
Abstract
There is increasing interest in localizing nerves in the intestine, especially specific populations of nerves. At present, the usual histochemical marker for cholinergic nerves in tissue sections is acetylcholinesterase activity. However, such techniques are applicable only to frozen sections and have uncertain specificity. Choline acetyltransferase (ChAT) is also present in cholinergic nerves, and we therefore aimed to establish a paraffin section immunocytochemical technique using an anti-ChAT antibody. Monoclonal anti-choline acetyltransferase (1.B3.9B3) and a biotin-streptavidin detection system were used to study the distribution of ChAT immunoreactivity (ChAT IR) in paraffin-embedded normal and diseased gastrointestinal tracts from both rats and humans. Optimal staining was seen after 6-24 hr of fixation in neutral buffered formalin and overnight incubation in 1 μg/ml of 1.B3.9B3, with a similar distribution to that seen in frozen sections. In the rat diaphragm (used as a positive control), axons and motor endplates were ChAT IR. Proportions of ganglion cells and nerve fibers in the intramural plexi of both human and rat gastrointestinal tracts were also ChAT IR, as well as extrinsic nerve bundles in aganglionic segments of Hirschsprung's disease. Mucosal cholinergic nerves, however, were not visualized. In addition, non-neuronal cells such as endothelium, epithelium, and inflammatory cells were ChAT IR. We were able to localize ChAT to nerves in formalin-fixed, paraffin-embedded sections. The presence of ChAT IR in non-neuronal cells indicates that this method should be used in conjunction with other antibodies. Nevertheless, it proves to be a useful technique for studying cholinergic neuronal distinction in normal tissues and pathological disorders.
Keywords
T
Although these antibodies are useful for studying general innervation, they do not discriminate among the subpopulations of both intrinsic and extrinsic gastrointestinal nerves that mediate their effects via multiple classical and peptide neurotransmitters (Furness and Costa 1987). However, recent studies employing immunohistochemical methods have shown alterations in tissue levels or nerve staining patterns for specific neuropeptides in gastrointestinal disorders, such as inflammatory bowel diseases (Sjolund et al. 1983; Goldin et al. 1989; Strobach et al. 1990; Kubota et al. 1992). Furthermore, studies in experimental animals suggest that neurotransmitter metabolism might be affected by the inflammatory process (Collins et al. 1989,1992; Swain et al. 1991,1992). It has even been suggested that cholinergic nerves may participate in the interrelationship between the autonomic nervous system and the immune system (Rinner and Schauenstein 1993). Such studies demonstrate the importance of looking at specific populations of nerves in researching gastrointestinal disorders.
Acetylcholine is a major neurotransmitter in the enteric nervous system. Cholinergic nerves mediate increased gut activity, such as contraction (Debas and Mulvihill 1991), and are associated with mucosal ion transport (Isaacs et al. 1976; Cooke 1984; Chandan et al. 1991a,b; Javed and Cooke 1992). At present, acetylcholinesterase (AChE) activity is the usual marker of cholinergic nerves and has become a widely accepted technique for diagnosis of Hirschsprung's disease (Lake et al. 1978), in which extrinsic nerve fibers penetrate the full thickness of the aganglionic segment of colon (Mackenzie and Dixon 1987; Tam and Boyd 1990). However, it has long been recognized that the histochemical demonstration of AChE stains a variety of cholinergic and noncholinergic peripheral neurons, as reviewed by Costa et al. (1987) and recently discussed by Bleys et al. (1994). This lack of reliable specificity precludes the application of the AChE reaction to labeling of cholinergic nerves in the enteric nervous system, which is innervated by both cholinergic and noncholinergic nerves (Furness and Costa 1987).
Choline acetyltransferase (ChAT) is considered to be a more specific and reliable marker of cholinergic nerves (Schemann et al. 1993). However, this enzyme is found in relatively small amounts in neural tissue, complicating the development of immunostaining methods in the enteric nervous system (Schemann et al. 1993). Therefore, immunocytochemistry for ChAT has as yet been applied only to frozen sections and whole mounts in a limited number of studies in the enteric nervous system (Furness et al. 1984,1985; Schemann et al. 1993,1995; Mann et al. 1995). Whole mounts and frozen section preparations do not permit the same microscopic resolution as paraffin sections and preclude the use of archival tissue. Accordingly, a paraffin section immunocytochemical method to localize cholinergic nerves would be very useful in studying normal and diseased gastrointestinal tracts and cholinergic neuronal distinction in a variety of other pathological disorders. Therefore, we developed an immunocytochemical technique to localize ChAT in human and rat formalin-fixed, paraffin-embedded tissue sections using monoclonal antibody 1.B3.9B3 (Ostermann-Latif et al. 1992). Using this technique, we found ChAT immunoreactivity (ChAT IR) in both nerves and nonneuronal cells.
Materials and Methods
Tissues
Diaphragm (as positive control) (Molenaar 1990), jejunum, ileum, and colon were taken from Lewis, Sprague-Dawley, and Wistar rats. Jejunum was also taken from Sprague-Dawley rats 6 days after infection with Trichinella spiralis (Ts) (Swain et al. 1992) and from Wistar rats 2–3 weeks after infection with Nippostrongylus brasiliensis (Nb) (Stead et al. 1991). The animals were sacrificed by ether anesthesia or by cervical dislocation, then promptly dissected. Tissues were immediately fixed by immersion.
Blocks of human tissues were retrieved from the pathology files of Chedoke-McMaster Hospitals and the University of Leeds. These included 12 cases of Hirschsprung's disease (both aganglionic and noninvolved segments) and nine control colon segments which were selected and screened for the presence/absence of ganglion cells in H & E-stained slides. A variety of normal tissue blocks with no specific pathological abnormalities were also taken. These included bladder, cervix, gallbladder, heart, kidney, liver, lung, skeletal muscle, pancreas, parathyroid, placenta, spleen, sympathetic ganglion, thymus, thyroid, tongue, tonsil, trachea, umbilical cord, and uterus.
Tissue Fixation and Processing
For rat tissues, a variety of fixatives were tried: absolute ethanol (6 hr), Bouin's fluid (6 hr), 10% neutral buffered formalin (NBF; 6 and 24 hr) and 1% paraformaldehyde (6 hr). The tissues were then processed to paraffin overnight. Human tissues were fixed in NBF (McMaster) or 10% aqueous formalin (Leeds) for 6-72 hr, then processed to paraffin. Sections were cut at 2-3 μm, collected on aminopropyltriethoxysilane- coated slides, and dried for 6-24 hr in a 45C oven. Additional pieces of rat gut were quenched in liquid nitrogen and sectioned in a cryostat at 5 μm.
Immunocytochemistry
Two antibodies against ChAT were evaluated: monoclonal antibody 1.B3.9B3 (Boehringer-Mannheim; Dorval, PQ, Canada) and rabbit polyclonal antiserum AB143 (Chemicon; El Segundo CA). Sections were dewaxed in xylene, rinsed in ethanol, and endogenous peroxidase activity was blocked in 0.5% hydrogen peroxide in methanol (30 min). Sections were rinsed in 70% ethanol, tapwater, distilled water, and Tris-buffered saline, pH 7.6 (TBS). Next, the sections were incubated in 20% normal rabbit serum (NRS) or 20% normal goat serum (NGS) in TBS to block nonspecific binding and then incubated at room temperature in 0.25-10 μg/ml 1.B3.9B3 diluted in 5% NRS in TBS (16-24 hr) or 1:100-1:10,000 AB143 diluted in 5% NGS in TBS (1 hr). For 1.B3.9B3, shorter (1 hr) and longer (45 hr at 4C) incubations were tried. The primary antibodies were detected with Zymed Histostain SP kits (Dimension Laboratories; Missisauga, ON, Canada). This entailed incubation in biotinylated rabbit anti-mouse or goat anti-rabbit immunoglobulins (Igs; 10 min) and rinses with TBS before application of the streptavidin-peroxidase complex (5 min). To prevent crossreactivity with endogenous rat Igs, 10% rat serum was added to the biotinylated rabbit anti-mouse secondary antibody. Color was developed for 30–60 min in aminoethylcarbazole.
In some 1.B3.9B3 experiments, double cycling of the secondary antibody and streptavidin complex was used to enhance staining. Other techniques included the use of the proteolytic enzymes trypsin (Sigma, St Louis MO; 0.1% in TBS) and pepsin (Sigma; 0.1% in 0.1 M HCl) and of the detergent Triton X-100 (BioRad, Missisauga, ON, Canada; 0.1%) in an effort to facilitate penetration of the antibody. We also tried secondary fixation with aqueous picric acid or mercuric chloride. Cryostat sections were fixed in acetone at 4C and stained under the same conditions as 1.B3.9B3 (1-hr incubation).
Controls included adsorption of 1.B3.9B3 with human placental ChAT (EC 2.3.1.6; Sigma) at 1 μg/ml, 10 μg/ml, and 100 μg/ml, and parallel staining with isotype-matched (IgG1) irrelevant antibody (anti-hepatitis B surface antigen; V2.6E4.2E9.2C5; Sera-Lab, Dimension Laboratories) at the same concentrations as 1.B3.9B3.
Results
Technical Considerations
Similar patterns of 1.B3.9B3 immunoreactivity (ChAT IR) were seen in paraffin sections of rat tissues after all types of fixation, and in frozen sections. The best staining was observed in tissues fixed for 24 hr in NBF (Figure 1A), with 6-hr NBF fixation also giving good results. Optimal staining was obtained by incubating sections in 1 μg/ml of 1.B3.9B3 for 16-18 hr. The use of trypsin, pepsin, Triton X, secondary fixatives (aqueous picric acid and mercuric chloride), and double cycling of the secondary and tertiary reagents did not significantly enhance the staining.
ChAT IR was mostly blocked by soluble adsorption with 10 μg/ml of human placental ChAT and was completely blocked by 100 μg/ml (Figure 1B). Tissues stained with the antibody against hepatitis B surface antigen were negative, although this antibody produced strong staining in the hepatitis B-positive liver control.
AB143 stained muscle intensely. Adsorbing this antiserum with dried skeletal muscle at 100 μg/ml did not improve results, and we therefore decided to work only with the monoclonal.
ChAT IR in Rat Tissues
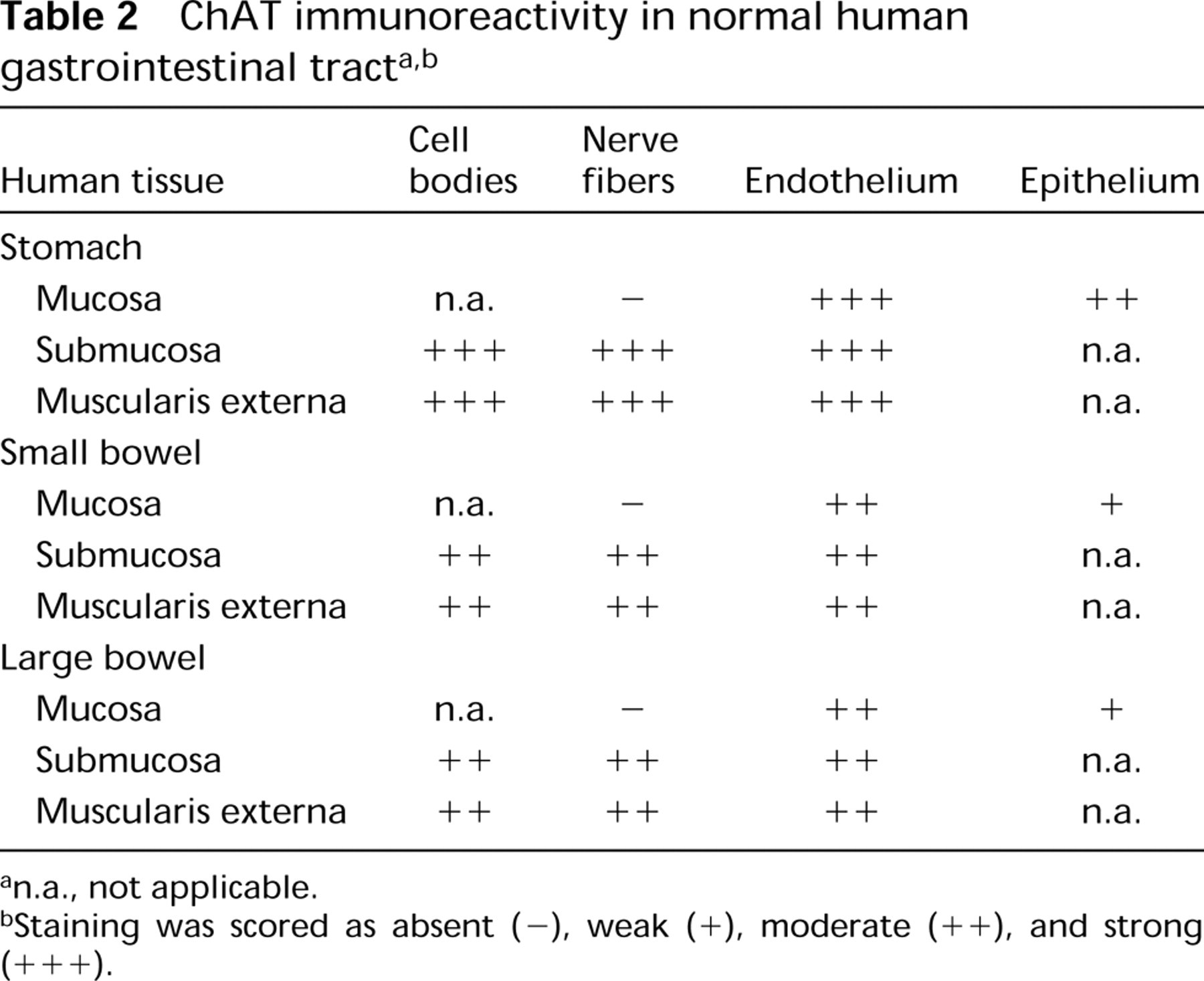
The positive control tissue (diaphragm) had consistently strong ChAT IR axons and neuromuscular junctions at the muscle cells (Figures 1A and 1C). In the jejunum, some neuronal cell bodies in both the submucosal and myenteric plexi were well stained (Figures 2B–2D), with occasional ChAT IR nerve terminals impinging on the somata (Figures 2B and 2C). Villous epithelial cells were strongly ChAT IR in both paraffin and frozen sections, and some inflammatory cells in the lamina propria were moderately ChAT IR (Figure 2A). Lymphoid cells in the epithelium and occasional cells in submucosa or muscularis externa were also ChAT IR. However, mucosal nerves were not visualized in either paraffin or frozen sections. In ileum and colon the patterns were very similar, but epithelial ChAT IR was minimal. For results on NBFfixed rat gastrointestinal tissues, refer to Table 1.
Similar results were seen in both Ts- and Nb-infected jejuna. Submucous and myenteric neurons were moderately ChAT IR and mucosal nerves were not visualized. However, many inflammatory cells in the lamina propria and submucosa were also ChAT IR (Figure 3A). Many of these appeared to be eosinophils. Epithelial staining was minimal and patchy, with regenerative epithelium essentially negative, but intraepithelial lymphocytes (IELs) were strongly positive (Figure 3B). Sections of nematodes also stained (data not shown).
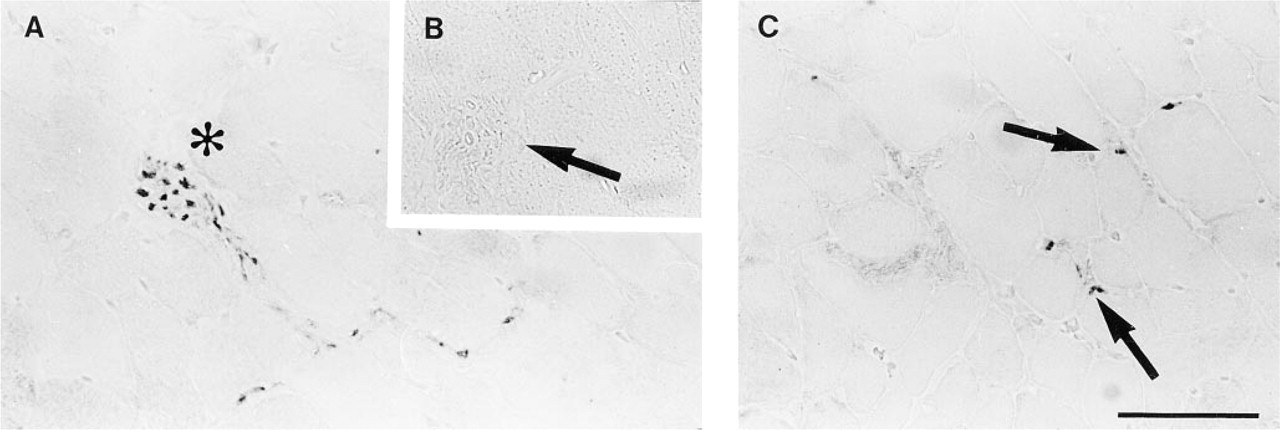
ChAT IR in rat diaphragm. (
ChAT IR in normal rat jejunum. (
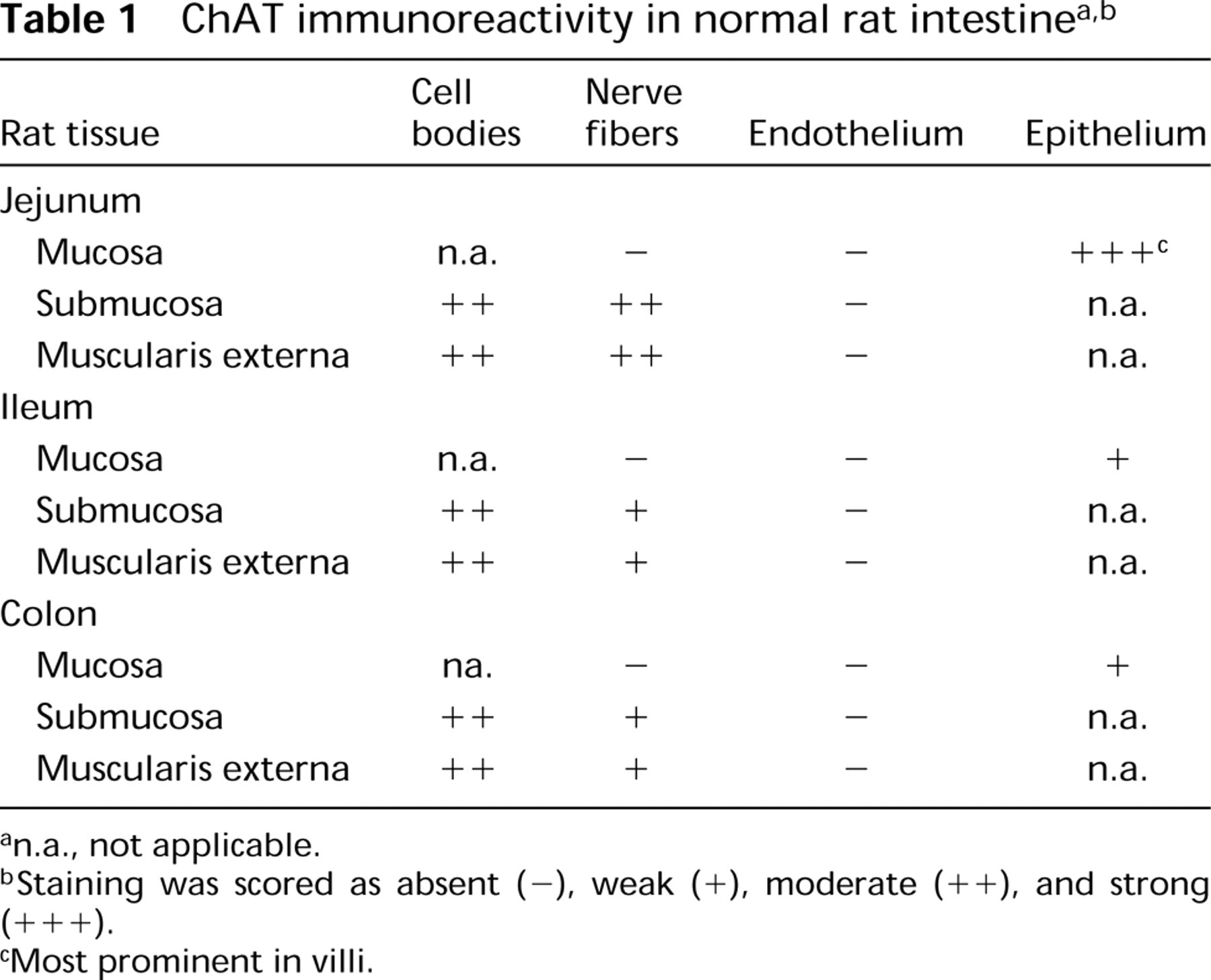
n.a., not applicable.
Staining was scored as absent (—), weak (+), moderate (++), and strong (+++).
Most prominent in villi.
ChAT IR in Human GI Tract
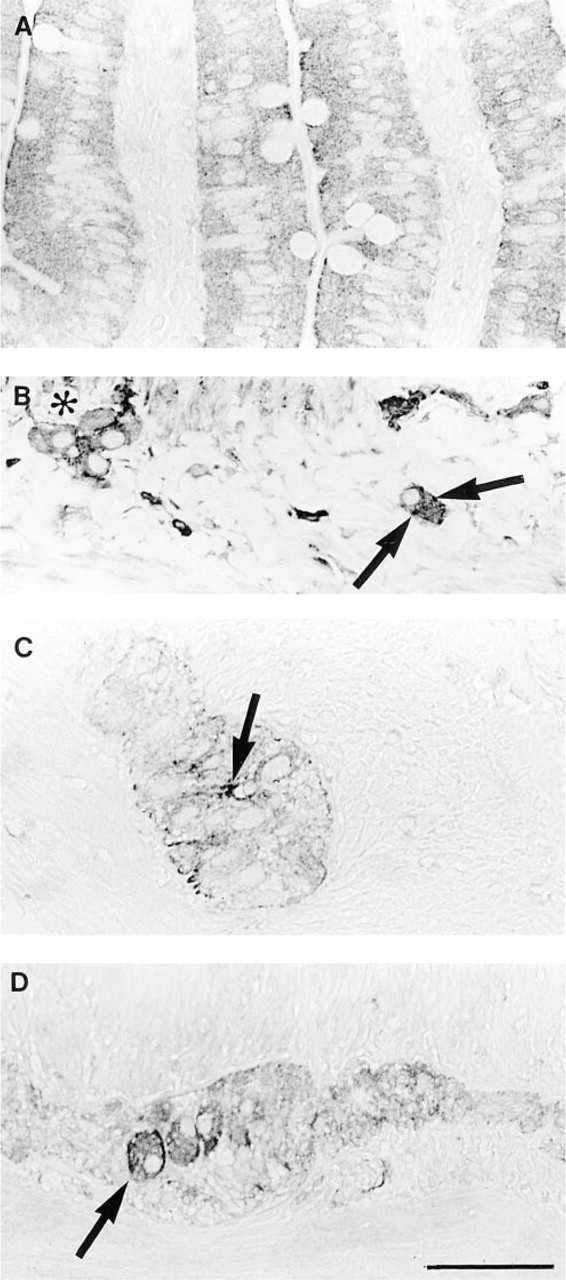
Esophagus, rectum, and anus all had ChAT IR nerve and endothelium, and esophagus and rectum both had ChAT IR epithelium. Esophagus also had weak striated muscle staining and rectum had ChAT IR cells in the lamina propria. Anus showed moderately ChAT IR connective tissue cells and weakly stained muscle. Gallbladder had ChAT IR endothelium only. A summary of the results for human stomach and for small and large bowel is presented in Table 2. In stomach, inflammatory cells were weakly positive in the lamina propria. Epithelium was moderately ChAT IR, and parietal cells exhibited stronger staining. Submucosal and myenteric neurons and endothelium were strongly positive. In the small bowel, inflammatory cells in the lamina propria were ChAT IR and epithelial staining was variable. Submucosal and myenteric neurons were moderately ChAT IR, mesothelium was ChAT IR, and endothelium stained moderately. In the large bowel, there was weak staining of cells in the lamina propria, variable staining of epithelium, and stronger staining of submucosal and myenteric neurons and endothelium (Figure 4).
ChAT IR in Other “Normal” Human Tissues
In view of the strong ChAT IR in human endothelium (often of equivalent intensity to the nerve staining) and patchy moderate epithelial staining, we screened nongastrointestinal tissues to compare the relative intensities of ChAT IR in endothelium, epithelium, and nerve. In these tissues, moderate to strong ChAT IR was found in nerve and endothelium, with weaker staining of some epithelia. Although various epithelial cells stained weakly to moderately, e.g., diffuse staining in hepatocytes, other epithelia, such as pancreas and endometrial glands, were not stained in the limited number of samples we examined. Stromal cells were variably ChAT IR in many tissues, and smooth and striated muscle sometimes exhibited weak staining. Serosal membranes were consistently moderately stained. In the sympathetic ganglion, the presynaptic nerves and synaptic junctions were ChAT IR, whereas the ganglion cells themselves were not stained.
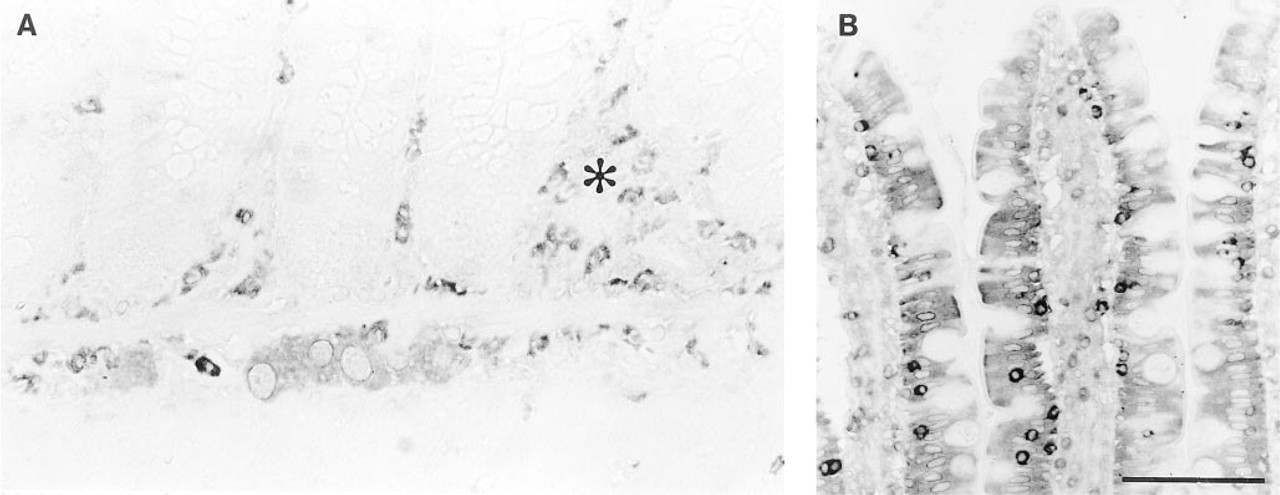
ChAT IR in nematode-infected rat jejunum. (
Hirschsprung's Disease
The aganglionic sections had very strong ChAT IR nerve bundles in the submucosa and muscularis externa, but also strongly stained endothelium and variable epithelial staining (data not shown) (Figure 5). The noninvolved (proximal) colon showed the same characteristic staining pattern as normal gut, with positive submucosal and myenteric neurons, as well as ChAT IR endothelium and epithelium. The mucosal nerves did not stain in either aganglionic or proximal segments.
Discussion
Monoclonal antibody 1.B3.9B3 was applied to formalin- fixed, paraffin-embedded sections to localize ChAT immunoreactivity in the gastrointestinal tract. Using this novel staining technique on normal tissues, we succeeded in immunostaining nerve fibers and a proportion of cell bodies in submucous and myenteric plexi, as well as nerve and neuromuscular junctions in rat diaphragm. We also observed strong ChAT IR in endothelial cells in human tissues, moderate staining in epithelia in both human and rat intestine, and staining of inflammatory cells, such as eosinophils and intraepithelial lymphocytes.
For several reasons, we believe that 1.B3.9B3 immunoreactivity reflects a true distribution of ChAT. The similar patterns of staining seen in paraffin and frozen sections of rat gut indicate that fixation and processing did not affect the specificity of the antibody. This is supported by the abolition of staining by preadsorption of 1.B3.9B3 with human placental ChAT and the lack of staining with an irrelevant, isotypematched monoclonal antibody (anti-hepatitis B surface antigen). The localization of ChAT IR in the submucosal and myenteric plexi is consistent with other reports that employed immunocytochemistry on whole mounts (Furness et al. 1984,1985; Schemann et al. 1993,1995; Mann et al. 1995), including studies that successfully used 1.B3.9B3 (Schemann et al. 1993,1995). In addition, ChAT IR and activity in endothelia of various tissues have been previously described (Parnavelas et al. 1985; Gonzalez and Santos-Benito 1987; Milner et al. 1989), and there are reports of ChAT in epithelial cells (Grando et al. 1993; Sakuragawa et al. 1997) and lymphocytes (Rinner and Schauenstein 1993).
n.a., not applicable.
Staining was scored as absent (—), weak (+), moderate (++), and strong (+++).
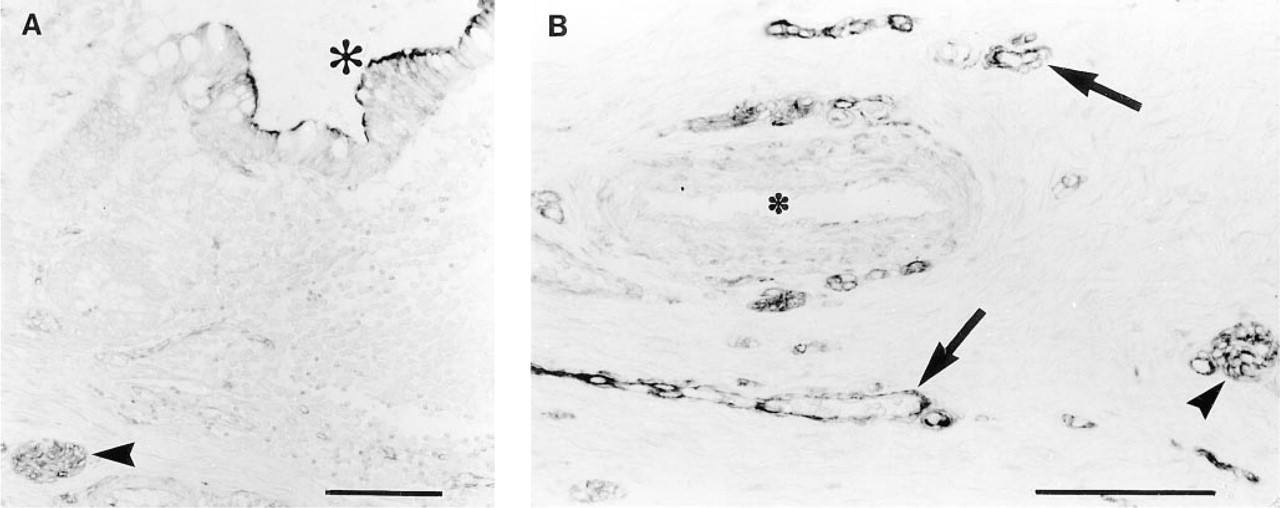
ChAT IR in normal human colon. (
Although we were able to consistently localize ChAT IR in the submucous and myenteric plexi, we did not observe convincing mucosal nerve staining in either human or rat tissues. In human tissues we often observed a few linear bands of staining in the mucosa, but in view of the strong vascular staining seen elsewhere, we interpreted this as endothelial immunoreactivity. Although functional, pharmacological, and physiological evidence suggests that a population of cholinergic nerves innervate the mucosa (Isaacs et al. 1976; Cooke 1984; Chandan et al. 1991a,b; Javed and Cooke 1992), we are unaware of any reports providing conclusive histological evidence. Reports using AChE histochemistry on normal mucosa either have dubious specificity (Isaacs et al. 1976) or report few or no cholinergic nerve fibers (Lake et al. 1978). Studies with ChAT immunohistochemistry on whole mounts have traced ChAT IR nerve processes to the mucosa but have not demonstrated ChAT IR nerve fibers in this compartment (Furness et al. 1985). Possibly, ChAT is not transported into mucosal nerve processes, or a non-1.B3.9B3-immunoreactive ChAT isoenzyme is present in mucosal nerves (for a discussion of ChAT expression, see Mallet et al. 1990). It is also possible that mucosal ChAT is non-1.B3.9B3 immunoreactive because it has interacted with other proteins or stuctures in the mucosa and has been conformationally altered, thus masking or hiding the epitope recognized by 1.B3.9B3. Probably, the lack of mucosal nerve staining is a consequence of the small amount of ChAT present in enteric neural tissues (Schemann et al. 1993).
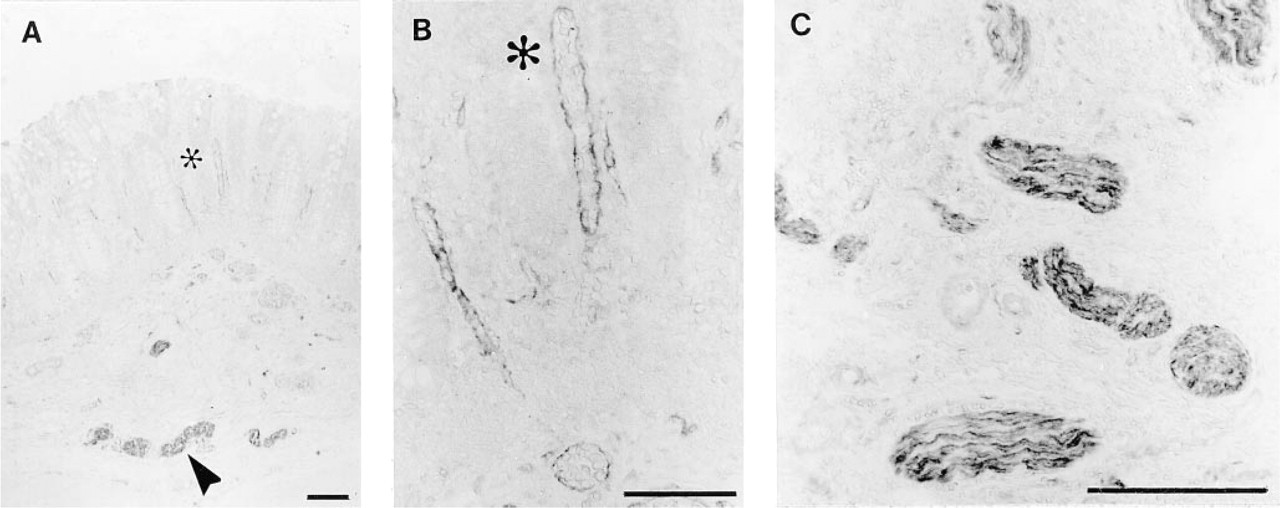
ChAT IR in an aganglionic section of Hirschsprung's disease colon. (
The pattern of epithelial ChAT IR seen in both human and rat may be connected to the role of acetylcholine in mucosal ion transport (Isaacs et al. 1976; Cooke 1984; Chandan et al. 1991a, b; Javed and Cooke 1992). In this context, it is interesting to note that acetylcholinesterase activity is present in some epithelia, including large bowel (Sine et al. 1991), epidermis (Grando et al. 1993), and amniotic epithelial cells (Sakuragawa et al. 1997), and that ChAT is also capable of hydrolyzing acetylcholine (Molenaar 1990). Skin and amniotic epithelial cells have been shown both to contain ChAT and to metabolize acetylcholine (Grando et al. 1993; Sakuragawa et al. 1997). However, the literature does not provide an explanation for the observed pattern of stronger staining in the jejunal villi and weaker or negative staining in the jejunal crypts, ileum, and colon. Although a role for ChAT in epithelial cells is unclear, the patchy distribution with adjacent positive and negative cells, as well as the similar distribution seen in frozen sections, indicates that this was not an artifact of tissue fixation and processing.
The ChAT IR distribution in nematode-infected jejuna reveals that the total acetylcholine handling capacity may change in inflamed mucosae and that the major cell types capable of acetylcholine metabolism differ. For example, the epithelium of inflamed jejuna was weak and patchy, with regenerative epithelium clearly negative, but there was strong staining of intraepithelial lymphocytes. Furthermore, there were increased numbers of ChAT IR inflammatory cells in the mucosa and submucosa, many appearing to be eosinophils, which are found in significant numbers in the jejuna of nematode-infected rats (Nawa and Hirashima 1984). In light of these inflammatory changes, it is interesting that altered cholinergic metabolism in animal models of intestinal inflammation has been reported (Collins et al. 1989). Because ChAT can catalyze both the degradation and synthesis of acetylcholine (Molenaar 1990), our data suggest that information obtained from tissue homogenates should be interpreted with caution. Altered levels of acetylcholine or ChAT might be a reflection of the inflammatory process and not of altered nerve activity or distinction.
Hirschsprung's colon is characterized by a lack of nerve cell bodies in both submucosal and myenteric plexi (Mackenzie and Dixon 1987). The parasympathetic pelvic splanchnic nerves, which normally innervate the large bowel, do not synapse in the bowel wall, and instead penetrate the smooth muscle to innervate the mucosa (Mackenzie and Dixon 1987). These presumed extrinsic parasympathetic preganglionic axons were ChAT IR in the submucosa and muscularis externa of our samples of Hirschsprung's colon, but mucosal nerve fibers were not ChAT IR. This lack of mucosal nerve staining is surprising because the substantially increased numbers of AChE-positive nerve fibers in the mucosa of Hirschsprung's disease routinely serve as a useful marker for diagnosis (Lake et al. 1978). The discrepancy in staining distribution may be connected to the dubious specificity of the AChE technique (Costa et al. 1987), which suggests that the increased numbers of AChE-positive fibers may not be totally representative of actual cholinergic innervation. However, the complete absence of mucosal nerve staining is probably due to the paucity of ChAT in cholinergic nerves (Schemann et al. 1993), precluding the use of our method as an alternative to the AChE technique in the diagnosis of Hirschsprung's disease.
To investigate ChAT IR distribution in tissues other than the gastrointestinal tract, we studied a variety of normal tissues. In all tissues, nerves were consistently labeled, endothelium was ChAT IR, epithelia expressed different degrees of ChAT staining, and some stromal and inflammatory cells were also ChAT IR. Because the tissues were selected from the McMaster pathology files, variations in staining intensity from case to case presumedly reflect differences in tissue handling and length of fixation. This suggests that the method needs to be optimized, depending on tissue source and fixation time.
In summary, we have developed an immunocytochemical, paraffin section technique, using a monoclonal antibody against ChAT (1.B3.9B3), to localize cholinergic structures in human and rat gastrointestinal tract. In neural tissue, ChAT IR was found in a proportion of cell bodies and fibers in the myenteric and submucosal plexi in normal intestines, in thick nerve bundles in the muscularis externa and submucosa of Hirschspung's colon, and in nerve in a variety of tissues. Mucosal nerves, however, were not visualized.
In non-neuronal structures, ChAT IR was observed in endothelium, epithelium, and in certain inflammatory cells.
The paraffin section immunocytochemical technique described in this report, using 1.B3.9B3 to localize choline acetyltransferase immunoreactivity, has several potential applications. Two clear uses are as follows. In known neuronal tissues, 1.B3.9B3 could be used to identify cholinergic elements and to distinguish these from noncholinergic structures. Second, this method can also be applied to the investigation of potential neural tumors. However, because ChAT is known to be present in non-neuronal structures, as reproduced herein, 1.B3.9B3 would be best used in conjunction with an antibody panel including other neuronal, epithelial, and vascular markers, such as PGP 9.5, keratin, and Factor VIII.
Footnotes
Acknowledgments
Supported by the Medical Research Council of Canada and by the Crohn's and Colitis Foundation of Canada.
We would like to thank M.G. Blennerhassett and K.A. Davis for providing some samples and assistance, and A. Beltrano, E.C.C. Colley, M. Falbo, B. Hewlett,. Lhoták, and E. LaForme for their invaluable help. We also appreciate Boehringer-Mannheim's support in providing us with some of the reagents for our experiments.