
Abstract
Cholinergic innervation of the heart has been analyzed using cholinergic markers including acetylcholinesterase, choline acetyltransferase (ChAT), and vesicular acetylcholine transporter (VAChT). In the present study we demonstrate putative cholinergic nerves in the rat heart using an antibody to ChAT of a peripheral type (pChAT), which is the product of a splice variant of ChAT mRNA and preferentially localized to peripheral cholinergic nerves. Expression of mRNAs for pChAT and the conventional form of ChAT (cChAT) were verified in the rat atrium by RT-PCR. Localization of both protein products in the atrium was confirmed by Western blotting. Virtually all neurons and small intensely fluorescent cells in the intrinsic cardiac ganglia were stained immunohistochemically for pChAT. The density of pChAT-positive fibers was very high in the conducting system, high in both atria, the right atrium in particular, and low in the ventricular walls. pChAT and VAChT immunoreactivities were closely associated in some fibers and fiber bundles in the ventricular walls. These results indicate that intrinsic cardiac neurons homogeneously express both pChAT and cChAT. Furthermore, innervation of the ventricular walls by pChAT- and VAChT-positive fibers provides morphological evidence for a significant role of cholinergic mechanisms in ventricular functions.
Keywords
T
To date, four possible marker proteins have been used to reveal cholinergic nerves: (1) ACh degrading enzyme, acetylcholinesterase (AChE); (2) ACh synthesizing enzyme, choline acetyltransferase (ChAT); (3) vesicular ACh transporter (VAChT), which mediates transport of ACh to storage vesicles in nerve terminals; and (4) high-affinity choline transporter (CHT), which transports choline into neurons for the synthesis of ACh. AChE histochemistry has shown the distribution of putative cholinergic cells and fibers in the heart (Baluk and Gabella 1990; Crick et al. 1994,1996; Steele et al. 1994; Petrecca and Shrier 1998). However, AChE is not a specific marker for cholinergic nerves because it is also present in sensory and sympathetic fibers (Koelle 1955). ChAT is accepted to be the most reliable marker for cholinergic structures, but it has been recognized that most ChAT antibodies fail to identify cholinergic nerves in peripheral tissues (Arvidsson et al. 1997; Hoover et al. 2004). A few antibodies to ChAT have been used successfully to analyze positive cells and fibers in the heart of the guinea pig (Mawe et al. 1996; Horackova et al. 1999; Leger et al. 1999; Calupca et al. 2000; Hoover et al. 2004), rat (Richardson et al. 2003), and mouse (Mabe et al. 2006). In these studies, all intrinsic cardiac neurons were consistently reported to be immunoreactive for ChAT. However, only a few previous studies described the distribution of ChAT-positive fibers in the heart outside the cardiac ganglia (Hoover et al. 2004).
A splice variant of ChAT cDNA, which lacks exons 6-9 in the coding region, has been cloned from rat pterygopalatine ganglion (Tooyama and Kimura 2000). Because of its predominant localization in peripheral neurons, the protein product of the mRNA variant was designated ChAT of a peripheral type (pChAT). The conventional ChAT protein, found in both central and peripheral neurons, was called ChAT of the common type (cChAT). Although the antibody against pChAT is capable of detecting some positive neurons in the central nervous system (Kanayama et al. 2003; Yasuhara et al. 2003), pChAT immunohistochemistry (IHC) proves to be a powerful tool to visualize peripheral cholinergic structures (Nakanishi et al. 1999; Nakajima et al. 2000; Chiocchetti et al. 2003; Yasuhara et al. 2004). By pChAT IHC, for example, three types of cholinergic neurons can be classified according to the expression patterns of cChAT and pChAT in the enteric nervous system (Chiocchetti et al. 2003). Because intrinsic cardiac neurons show functional diversity and multiple chemical coding like enteric neurons (Steele et al. 1994; Horackova et al. 1999; Richardson et al. 2003), it is of interest to examine whether intrinsic cardiac neurons express pChAT in addition to cChAT. It is possible that heterogeneity in expression of pChAT in cardiac neurons would contribute to functional diversity of such neurons.
In the present study, therefore, we examined the possible expression of pChAT and its mRNA in intrinsic cardiac neurons of the rat by RT-PCR, Western blot analysis, and IHC. Colocalization of pChAT immunoreactivity with various cholinergic and non-cholinergic neurochemical markers was examined in intrinsic cardiac neurons using double-IHC techniques. In addition, IHC distribution of pChAT-positive fibers in the rat heart was examined and compared with those of other cholinergic markers including AChE, cChAT, and VAChT. We particularly focused on the cholinergic innervation of rat ventricular myocardium.
Materials and Methods
Animals
Male Wistar rats (Clea Japan Inc.; Tokyo, Japan) weighing 250-350 g were used. Procedures involving animals and their care were conducted in conformity with the standards for animal experiments in our university and are in compliance with the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals (1996). The animals were kept on a 12/12 hr light/dark schedule (lights on at 6 AM).
RT-PCR
Under deep anesthesia with pentobarbital (80 mg/kg, IP), two rats were perfused via the ascending aorta with 10 mM PBS, pH 7.4, to clear the blood. After perfusion, the hearts were removed and both atria were dissected out. Total RNA was isolated from these tissues using the acid guanidium thiocyanate-phenol method (Chomczynski and Sacchi 1987). Five μg of total RNA was then reverse transcribed for the first-strand cDNA synthesis using 80 U of SuperScript II (Gibco BRL; Gaithersburg, MD) and 500 ng of random hexamer (Amersham Biosciences Corp.; Piscataway, NJ). The exact sequences of the primers used for ChAT gene amplification are described.
Primers used were as follows: sense primer 5′-TGGGTCTCTGAATACTGGCTGAATG-3′ (nucleotide number 454-478 in exon 3-4) and antisense primer 5′-CTCACTCACTGAGTCAGCCCTGAC-3′ (nucleotide number 1282-1305 in exon 10). The number of nucleic acids of the rat ChAT cDNA and exon organization of the rat ChAT gene are based upon the reports by Brice et al. (1989) and Hahn et al. (1992), respectively. Expected sizes of the cChAT and pChAT PCR products are 852 bp and 224 bp, respectively.
The reaction mixture consisted of 2 ng/μl of the template cDNA, 0.8 μM each of the primers, 0.2 mM each of four deoxynucleotide triphosphates, and 0.025 U/μl AmpliTaq Gold polymerase in 1X PCR buffer (Applied Biosystems; Foster City, CA). After preincubation for 10 min at 95C, 40 cycles of PCR were performed with the profile of thermal cycles consisting of denaturation at 95C for 30 sec, annealing at 64C for 30 sec, and extension at 72C for 90 sec. PCR products were electrophoresed on a 3% agarose gel and stained with ethidium bromide. After dissecting out the bands from the gel, the target DNA in each band was eluted using the GeneClean II kit (Bio101 Inc.; La Jolla, CA) and cloned using a TA cloning system (Invitrogen; San Diego, CA). The pCR 2.1 plasmid vector containing the target DNA insert was transfected into the host
Western Blotting
Two rats were perfused with 10 mM PBS under deep anesthesia with pentobarbital (80 mg/kg, IP). The atria were dissected out and homogenized in 10 volumes of ice-cold 50 mM Tris-HCl (pH 7.4) containing 0.5% Triton X-100 and protease inhibitor mixture tablets (one tablet/10 ml; Complete Mini, Roche Diagnostics, Mannheim, Germany). After centrifugation for 20 min at 12,000 × g at 4C, the supernatants were collected as a crude protein fraction. This crude extracted protein was electrophoresed on a 10% SDS-PAGE under reducing conditions and then transferred onto a nitrocellulose membrane (Hybond-ECL; Amersham). The membrane was blocked for 1 hr with 20% Blocking Reagent-N102 (NOF Corp.; Tokyo, Japan) in 25 mM Tris-buffered saline (TBS, pH 7.4) at room temperature. It was then incubated overnight with the rabbit antiserum against pChAT (diluted 1:20,000) (Tooyama and Kimura 2000) or a goat polyclonal anti-cChAT antibody (diluted 1:500; AB144P, Chemicon, Temecula, CA) (Bruce et al. 1985) in 25 mM TBS containing 0.1% Tween 20 (TBST) at 4C. After washing with 25 mM TBST, the membrane was reacted for 2 hr with a peroxidase-coupled anti-IgG of an appropriate species (diluted 1:50,000; ImmunoPure, Pierce, Rockford, IL). Peroxidase labeling was detected by chemiluminescence using the SuperSignal West Pico Chemiluminescent Substrate (Pierce).
Tissue Preparations for Histochemistry
A total of eight rats were used. Animals were perfused under pentobarbital anesthesia (80 mg/kg, IP) on crushed ice through the ascending aorta with 10 mM PBS, followed by a fixative of 4% paraformaldehyde in 0.1 M PB, pH 7.4. After perfusion, the heart tissues were removed.
Three types of heart sections were prepared for enzyme histochemistry and IHC. First, the hearts from four rats were immersed en bloc for 2 days in the same fixative at 4C and then cryoprotected by immersion for 2 days in 0.1 M PB containing 15% sucrose at 4C. The heart tissues were then embedded in gelatin by immersion for 3 hr in 0.1 M PB containing 10% gelatin at 37C, followed by placement for 1 hr in a cold chamber at 4C. The gelatin-embedded tissues were re-fixed overnight with 4% paraformaldehyde in 0.1 M PB at 4C and then washed overnight in 0.1 M PB containing 15% sucrose at 4C. Twenty-μm-thick sections were cut transversely in a cryostat, parallel to the inferior diaphragmatic surface of the heart beginning superior to the SAN and ending inferior to the atrioventricular bundle of His (AVB). Cryostat sections were collected and stored in 0.1 M PBS containing 0.3% Triton X-100 (PBST).
Second, the atria were isolated from the heart tissues of four rats and pinned flat on a cork plate with the epicardium (dorsal side) up. These flat-mount atrial tissues, along with the remaining ventricular tissues, were then immersed for 2 days in 4% paraformaldehyde in 0.1 M PB at 4C. After cryoprotection by immersion for 2 days in 0.1 M PB containing 15% sucrose at 4C, atrial tissues were cut tangentially to the atrial wall into 20-μm-thick sections in a cryostat. For the third type of sections, ventricular tissues were cut longitudinally into 20-μm-thick sections in a cryostat, parallel to the long axis of the heart. The sections were collected and stored in 0.1 M PBST.
AChE Histochemistry
AChE histochemistry was performed according to the method of Tago et al. (1986). In brief, the sections were washed in 0.1 M maleate buffer (pH 6.0) and incubated for 2 hr at room temperature in a solution containing 18 μM acetylthiocholine iodide, 5μMK3Fe (CN)6, 30 μM CuSO4, and 50 μM sodium citrate in 0.1 M maleate buffer. After washing with 50 mM Tris-HCl buffer (pH 7.6), the sections were reacted with a mixture of 0.02% diaminobenzidine, 0.3% nickel ammonium sulfate, and 0.0045% H2O2 in 50 mM Tris-HCl buffer. Stained sections were washed in tap water, dried through a graded series of alcohol, cleared with xylene, coverslipped with Entellan (Merck; Darmstadt, Germany), and examined under a microscope.
IHC
Primary antibodies used in the present study are listed in Table 1. The sections were incubated for 2 days with each primary antibody at a working dilution at 4C, for 2 hr with a biotinylated secondary antibody of an appropriate species (diluted 1:1000; Vector Laboratories, Burlingame, CA) and for 1 hr with the avidin-biotinylated peroxidase complex (diluted 1:2000; ABC Elite, Vector Laboratories). Dilution of the reagents and washing of tissue sections between each step were done with PBST. Color development and processing of the stained sections were done as described above for AChE histochemistry. Adjacent sections were processed using the hematoxylin-eosin (H-E) stain or Masson's trichrome stain to identify the locations of cardiac ganglia and structures of the conducting system.
For IHC controls, pChAT antiserum was replaced with preimmune serum or with pChAT antiserum that had been preincubated overnight with the antigenic peptide of pChAT (Tooyama and Kimura 2000). No positive staining was observed in these control studies.
Double-Fluorescence IHC and Combination With AChE Staining
Atrial and ventricular sections were processed with double-fluorescence IHC procedures. Atrial sections were incubated overnight with the pChAT antibody combined with an antibody against protein gene product 9.5 (PGP9.5), cChAT, VAChT, tyrosine hydroxylase (TH), or the neuronal isoform of nitric oxide synthase (nNOS) (Table 1) at 4C. For the ventricular sections, a combination of pChAT and VAChT was examined. After washing, the sections were reacted for 2 hr in a mixture of Alexa Fluor 568-conjugated donkey anti-rabbit IgG (for pChAT) and either Alexa Fluor 488-conjugated donkey anti-mouse IgG (for PGP9.5, TH, and nNOS) or Alexa Fluor 488-conjugated donkey anti-goat IgG (for cChAT and VAChT) (diluted 1:500; Molecular Probes, Eugene, OR). After washing, the sections were mounted on gelatin-coated glass slides, coverslipped with glycerin, and imaged on a LSM510 confocal laser-scanning microscope equipped with an argon laser (458/488/514 nm) and a green helium/neon laser (543 nm) (Carl Zeiss; Oberkochen, Germany). Single optical slice images were taken using 40X Plan-Neofluar oil interface objective lens (Carl Zeiss) at a z-axis step of 0.8-μm depth. Brightness and contrast of the final images were adjusted using Adobe Photoshop 6.0 software (Adobe Systems; San Jose, CA).
Details of the primary antibodies used in the present study
pChAT, choline acetyltransferase of a peripheral type;
cChAT, choline acetyltransferase of the common type;
VAChT, vesicular acetylcholine transporter;
nNOS, neuronal nitric oxide synthase;
TH, tyrosine hydroxylase.
On some double-labeled sections of the atrium, staining for AChE activity was performed. After the sections were imaged and photographed under a laser-scanning microscope, coverslips were carefully removed by dipping in PBST. The sections on glass slides were then processed by AChE histochemistry as described above.
Results
Biochemical Detection of Two Forms of ChAT and Their mRNAs in the Rat Atrium
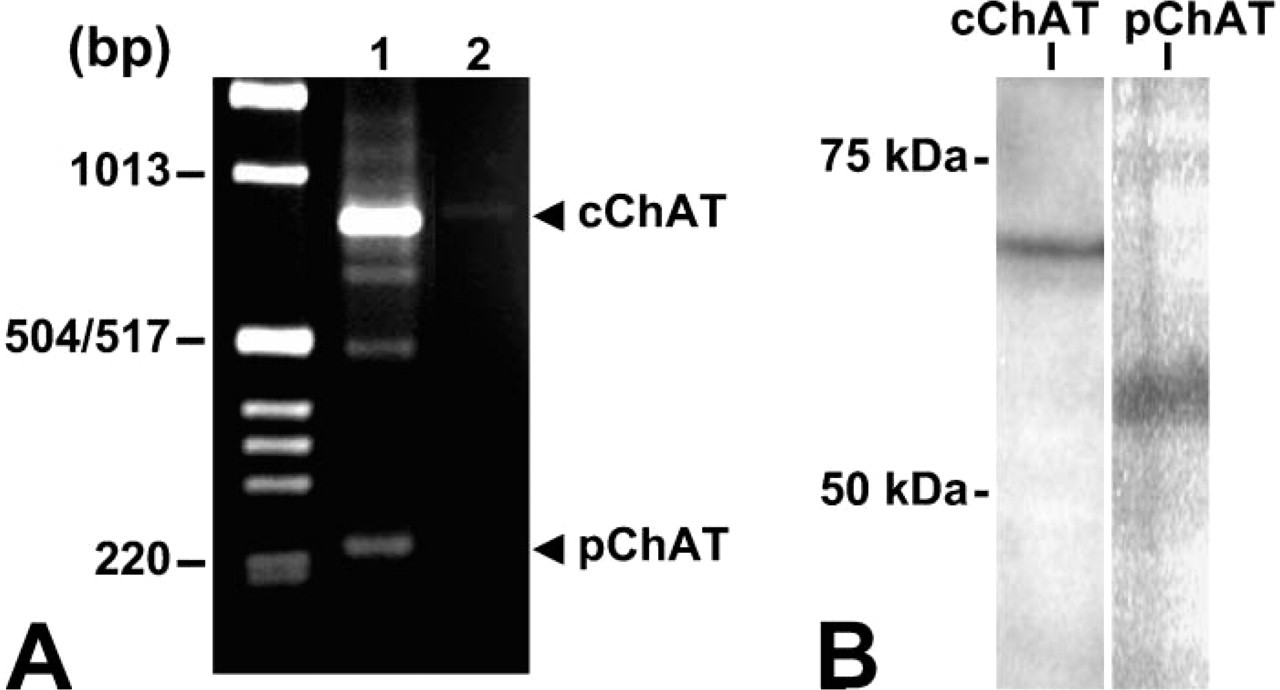
Figure 1A shows the result of RT-PCR in rat atrial tissues. Two bands were detected at the expected sizes for the cChAT (852 bp) and pChAT (224 bp) gene products, respectively, in the atrium. Nucleotide sequence analysis revealed that the larger and intense band represented the cChAT gene product. The smaller band was identified as the PCR product of the pChAT cDNA.
By Western blotting, pChAT antiserum detected a band at ∼55 kDa in the atrial tissues (Figure 1B). This size (55 kDa) is slightly larger than the deduced molecular size of pChAT (49 kDa), but agrees well with previous studies in the retina, iris, and trigeminal ganglion (Yasuhara et al. 2003,2004). The cChAT antibody detected a band of 68 kDa in the atrium, as previously reported in the striatum, retina, and iris (Yasuhara et al. 2003,2004).
pChAT IHC in Intrinsic Cardiac Neurons
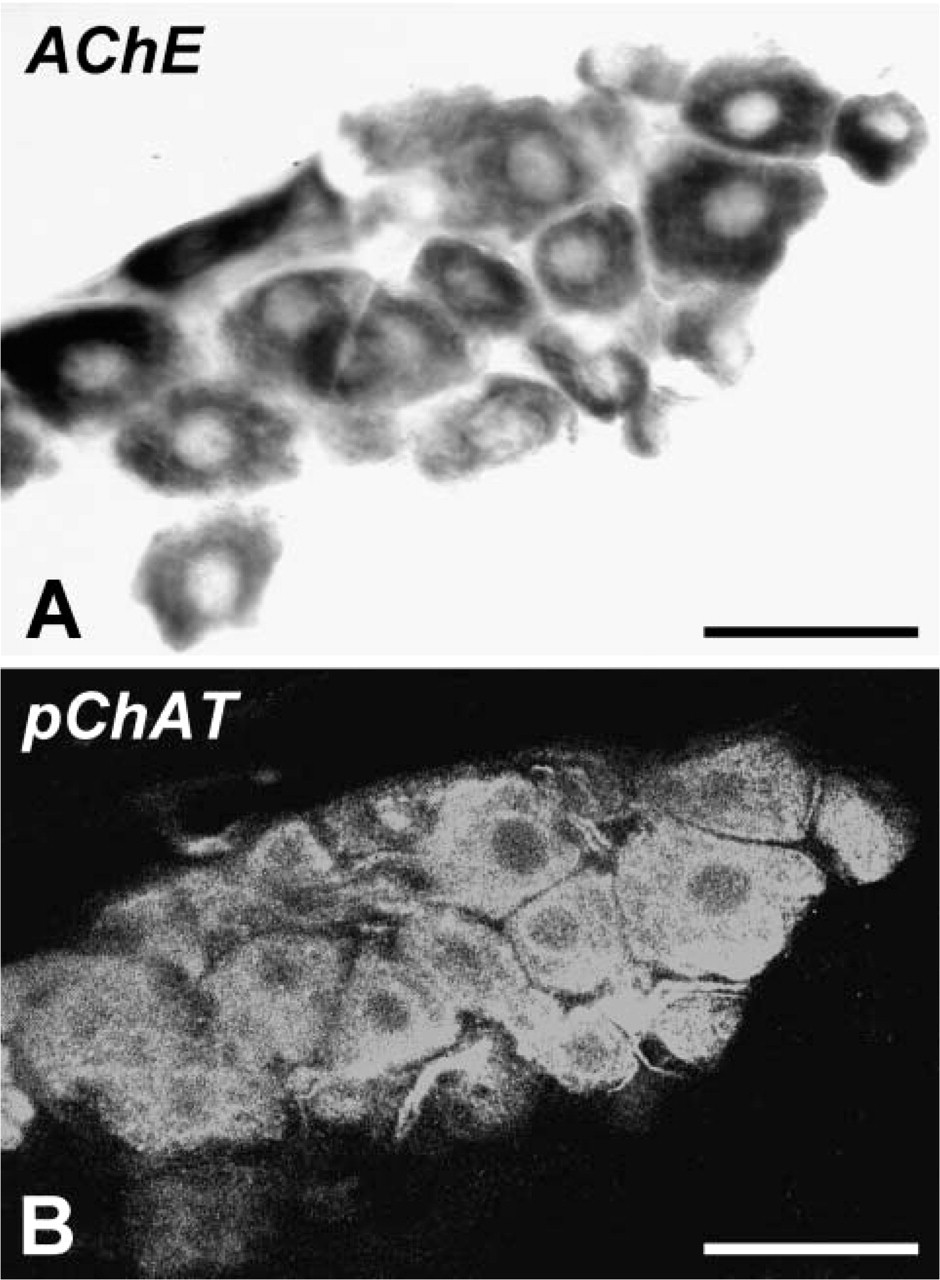
By H-E staining and AChE histochemistry, most intrinsic cardiac ganglia were identified in (1) the subepicardium of the right atrium between the venae cavae and (2) the dorsal part of the interatrial septum. Locations of rat cardiac ganglia were consistent with previous reports (Cheng et al. 1999; Richardson et al. 2003). pChAT IHC revealed many intrinsic neurons in the atrial wall, with a distribution pattern similar to that of AChE-positive cells. Somata of positive cells with diameters ranging from 20 to 50 μm were predominantly spheroid in shape. In double labeling, occurrence of pChAT immunofluorescence (Figure 2B) was overlapped completely with that of AChE activity (Figure 2A) in the somata of cardiac neurons.
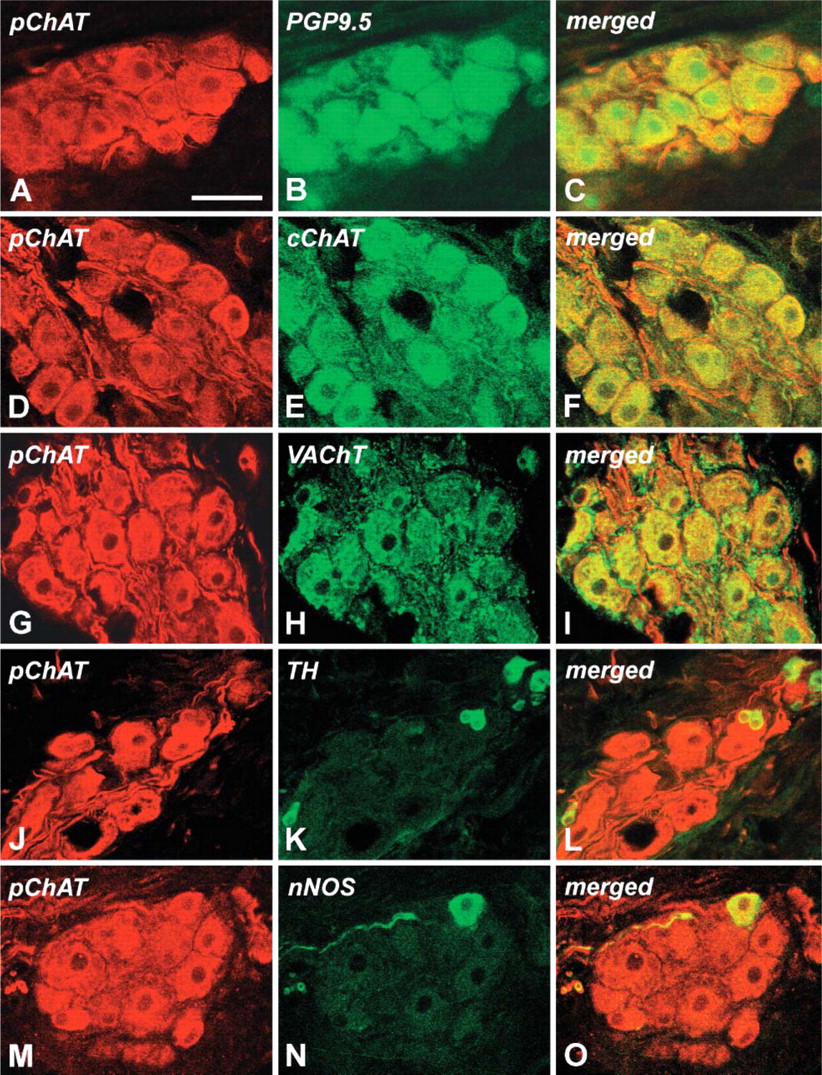
Figures 3A-3C show double labeling of cardiac neurons for pChAT and the general neuronal marker PGP9.5. pChAT immunoreactivity was localized exclusively in the cytoplasm (Figure 3A), whereas PGP9.5 immunoreactivity was localized in the cytoplasm and nucleus of positive neurons (Figure 3B). As shown, all neurons exhibiting PGP9.5 were also immunoreactive for pChAT and vice versa. Similarly, pChAT immunoreactivity was observed in all of the ganglion neurons immunoreactive for cChAT (Figures 3D-3F) and VAChT (Figures 3G-3I). The pChAT antibody intensely stained smooth fiber bundles in the ganglion (Figures 3D and 3G), whereas the cChAT antibody stained thin, sometimes varicose, fibers in the ganglion (Figure 3E). The antibody to VAChT stained neuronal somata rather weakly, but intensely stained a number of presumable terminal boutons surrounding cell somata double labeled for pChAT and VAChT (Figures 3H and 3I). The antibody to TH stained a few small cells in the ganglia that had round or spheroid cell somata with diameters ranging from 10 to 20 μm and one or two short processes (Figure 3K). Double labeling revealed that such small cells showed weak immunoreactivity for pChAT (Figures 3J-3L). In the ganglia, nNOS-positive neurons were occasionally found that were also immunoreactive for pChAT (Figures 3M-3O). Together it is highly probable that all neurons in the intrinsic cardiac ganglia, as well as small TH-positive cells, were immunoreactive for pChAT.
(
A combination of acetylcholinesterase (AChE) histochemistry (
Distribution of Nerve Fibers Positive for pChAT and Other Cholinergic Markers in the Heart Outside Intrinsic Cardiac Ganglia
Distribution and relative abundance of pChAT-, cChAT-, and VAChT-immunoreactive nerve fibers and AChE-positive fibers in rat heart outside intrinsic cardiac ganglia are summarized in Table 2. pChAT-immunoreactive nerve fibers were particularly abundant in the conducting system, including the SAN (Figure 4A), AVN (Figure 4B), and AVB (Figure 4C). In these structures, cChAT-positive fibers were also abundantly distributed (Figures 4D-4F). At low magnification, the AVN and AVB were diffusely stained by IHC for pChAT (Figure 4C) and cChAT (Figure 4F). Thick nerve bundles running near the AVB were intensely stained for pChAT (Figure 4C). VAChT-immunoreactive fibers and AChE-positive fibers were also densely distributed in the conducting system (data not shown).
The second highest density of pChAT-positive fibers was observed in the right atrial wall. Figure 5 shows comparison of the distribution patterns of AChE-(Figure 5A), pChAT- (Figure 5B), cChAT- (Figure 5C), and VAChT- (Figure 5D) positive fibers in tangential sections of the right atrium. Many thick bundles and a dense plexus of fine and occasionally varicose fibers were more or less stained for each marker protein. The thick fiber bundles were intensely labeled for AChE (Figure 5A) and pChAT (Figure 5B), whereas the VAChT antibody was powerful in labeling fine varicose fibers (Figure 5D). In gelatin-embedded transverse sections, such pChAT-positive thick bundles ran in the epicardium of the right atrium. The density of pChAT-positive fibers was less in the left atrium than in the right atrium. The right and left auricles also possessed dense networks of pChAT-positive fibers. The ascending aorta, pulmonary trunk, and aortic valve possessed no detectable nerve supply of pChAT-positive fibers.
As compared with the atrium, cholinergic innervation of the ventricular walls appeared relatively low. Figure 6 shows the distribution patterns of AChE-(Figures 6A, 6E, 6I, and 6M), pChAT- (Figures 6B, 6F, 6J, and 6N), cChAT- (Figures 6C, 6G, 6K, and 6O), and VAChT- (Figures 6D, 6H, 6L, and 6P) positive fibers in the ventricular walls. The ventricular walls contained a few fibers positive for AChE, pChAT, and VAChT, whereas cChAT-positive fibers were not visible or very sparse, if present, in the ventricular walls. In the right ventricular wall, AChE activity and pChAT immunoreactivity were observed mainly in thick bundles of smooth nerve fibers in the subendocardium (Figures 6A and 6B) and epicardium. The right ventricular myocardium contained a small number of smooth nerve fibers positive for AChE and pChAT. Nerve fiber bundles in the right ventricular wall were also labeled with the antibody to VAChT (Figure 6D), but this antibody labeled varicose fibers in the myocardium as well.
In the left ventricular wall, the distribution pattern of positive fibers for each protein was similar to that in the right ventricular wall. In the epicardial region, some nerve fiber bundles were stained for AChE (Figure 6E), pChAT (Figure 6F), and VAChT (Figure 6H). In the myocardium, AChE activity was found in networks of nerve fibers often running parallel to muscle fibers (Figure 6I). A small but significant number of pChAT-positive smooth fibers also ran parallel to muscle fibers (Figure 6J). VAChT immunoreactivity was observed mainly in fine varicose fibers in the myocardium (Figure 6L). The density of pChAT-positive fibers in the myocardium was lower than that of AChE-positive fibers or VAChT-positive fibers. A few fine positive fibers for AChE (Figure 6M), pChAT (Figure 6N), or VAChT (Figure 6P) occasionally reached the wall of coronary vessels in the left ventricular wall. The endocardial region of the left ventricle possessed almost no fibers stained for any of these molecules. The mitral and tricuspid valve cusps were supplied by a few AChE- and pChAT-positive fibers.
Distribution and relative abundance of pChAT-, cChAT-, VAChT-, and AChE-positive nerve fibers in the heart

Frequency of positive nerve fibers: −, rare or none; +, few; ++, moderate; +++, numerous.
Confocal images of intrinsic cardiac neurons stained by double-fluorescence IHC. (
To examine the relationship between pChAT- and VAChT-positive fibers in the left ventricular wall, double-fluorescence IHC was performed in ventricular sections. Figure 7 shows examples of positive fibers. In Figures 7A-7F, thick fiber bundles in the epicardium were stained for pChAT but not for VAChT. In contrast, fine varicose fibers were intensely labeled for VAChT and weakly for pChAT (Figures 7D-7F). In Figures 7G-7I, a thick fiber bundle was labeled for both pChAT and VAChT. In the myocardium, many varicose fibers were labeled for VAChT, but not for pChAT (Figures 7G-7L). A few smooth fibers in the myocardium were immunoreactive for both pChAT and VAChT (Figures 7J-7L).
Discussion
The present study demonstrates the expressions of pChAT and its mRNA in the rat heart by RT-PCR, Western blotting, and IHC. The RT-PCR study reveals the expressions of pChAT and cChAT mRNAs in the atrium. A previous study using laser-assisted micro-dissection and RT-PCR has shown that rat cardiac neurons express the non-coding R-exon of the ChAT gene (Kummer et al. 1998). The present study demonstrates for the first time that rat cardiac neurons express a splice variant of the ChAT mRNA in the coding region, in addition to the full-length transcript. The current Western blot analysis shows that the pChAT and cChAT antibodies recognize 55 kDa and 68 kDa proteins in the atrium, respectively. Therefore, it is confirmed that pChAT and cChAT, as well as their mRNAs, are expressed in the rat atrium.
pChAT IHC reveals positive neuronal somata in the intrinsic cardiac ganglia. Intrinsic cardiac neurons have been recognized as parasympathetic postganglionic efferent neurons that play roles in inhibiting cardiac functions. Previous IHC studies have indicated that all of the cardiac neurons are immunoreactive for cChAT in the guinea pig (Mawe et al. 1996; Horackova et al. 1999; Leger et al. 1999) and rat (Richardson et al. 2003). In addition, recent studies have described VAChT-immunoreactive intracardiac neurons in the rat (Arvidsson et al. 1997; Schäfer et al. 1998) and CHT-immunoreactive ones in the guinea pig and mouse (Hoover et al. 2004; Mabe et al. 2006). The present study provides evidence in support of the notion that all neurons in the cardiac ganglia are cholinergic, by demonstrating that the population of neurons immunoreactive for pChAT completely overlapped with that for PGP9.5, cChAT, and VAChT in the intrinsic ganglia of the rat heart.
Thus, it is likely that intrinsic cardiac neurons are a homogeneous population of neurons with regard to the cholinergic phenotype. Nevertheless, previous data have indicated that the intrinsic cardiac ganglia possess a heterogeneous population of neurons with regard to their morphology as well as their electrophysiological and pharmacological properties, including their chemical coding (Horackova and Armour 1995). For example, intrinsic neurons in the heart have been shown to contain, in various combinations, nNOS (Hassall et al. 1992; Klimaschewski et al. 1992; Mawe et al. 1996) and several neuropeptides including somatostatin, neuropeptide Y, and vasoactive intestinal peptide (Weihe et al. 1984; Steele et al. 1994; Horackova et al. 1999). A possible colocalization of pChAT and nNOS is of interest because a population of cChAT-positive neurons is known to overlap only partially with that of nNOS-positive neurons in the enteric nervous system (Costa et al. 1992; Chiocchetti et al. 2003). The present results show that the cardiac ganglia of the rat contain a few nNOS-positive neurons that are immunoreactive for pChAT. It is also noted that the presence or absence of nNOS does not affect the intensity and staining pattern for pChAT immunoreactivity in neuronal somata. Thus, it is suggested that pChAT and cChAT are homogeneously expressed in individual neurons in the cardiac ganglia, irrespective of the presence or absence of other neurochemical markers. This contrasts sharply with cholinergic neurons in the enteric nervous system where three types of cholinergic neurons can be classified according to the expression patterns of cChAT and pChAT (Chiocchetti et al. 2003). In the heart, it is probable that every cardiac neuron acts as a cholinergic nerve, the function of which is heterogeneously modulated by other coexisting proteins.
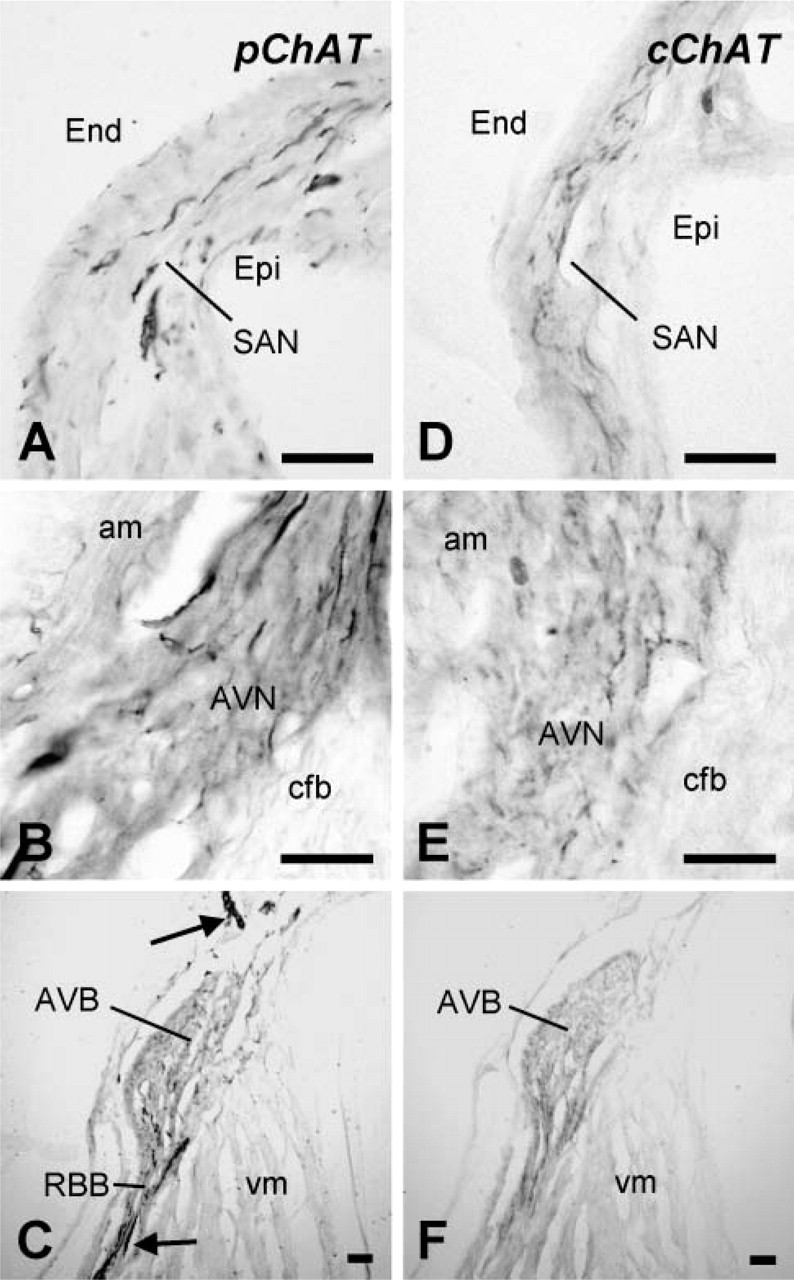
The conducting system in gelatin-embedded transverse sections of the rat heart stained for pChAT (
In addition to postganglionic neurons, small intensely fluorescent (SIF) cells, which express aspects of the catecholaminergic phenotype and contain TH at high concentrations, have been shown to occur throughout the atria, primarily associated with the intrinsic ganglia (Baluk and Gabella 1990; Mawe et al. 1996; Horackova et al. 1999; Leger et al. 1999; Slavíková et al. 2003). Previous studies showed discrepant results on the presence or absence of cChAT immunoreactivity in SIF cells. In the guinea pig heart, for example, cChAT immunoreactivity has been variously reported as in none (Leger et al. 1999), some (Horackova et al. 1999), or most (Mawe et al. 1996) SIF cells. In the present study, SIF cells in the cardiac ganglia exhibit intense immunoreactivity for TH and weak immunoreactivity for pChAT. The functions of cardiac SIF cells and the significance of coexpression of cholinergic and catecholaminergic phenotypes in such cells remain to be elucidated.
The mechanism and functional significance of generation of the two splice variant products in a single cell remain to be clarified. It is possible that pChAT may be produced to regulate ChAT activity. Alternatively, pChAT might exert different functions than cChAT in the same cell. We have previously indicated that pChAT has a low but significant enzyme activity (Yasuhara et al. 2003,2004). In addition, the pChAT preparation purified by immunoprecipitation from rat dorsal root ganglia exhibited significant ChAT activity (Bellier JP, et al., unpublished data). Therefore, it is likely that pChAT does act as an enzyme. Recently, Matsuo et al. (2005) demonstrated that pChAT differs from cChAT in intracellular transportation, probably reflecting a difference in physiological roles between pChAT and cChAT. Thus, it is tempting to speculate that pChAT and cChAT are responsible for ACh production in distinct intracellular compartments.
In the present study, pChAT- and cChAT-positive fibers are distributed very densely in the conducting system and densely in the right atrium. This is in accordance with previous studies that demonstrated the dense innervation of the conducting system by putative cholinergic fibers using AChE histochemistry (Wharton et al. 1981; Crick et al. 1994,1996; Petrecca and Shrier 1998) and IHC for cChAT (Hoover et al. 2004) and CHT (Hoover et al. 2004; Mabe et al. 2006). These morphological observations are compatible with physiological data showing that vagal nerve stimulation activates cholinergic input to the SAN and AVN, thus decreasing heart rate and atrioventricular conduction velocity (Ardell 2001).
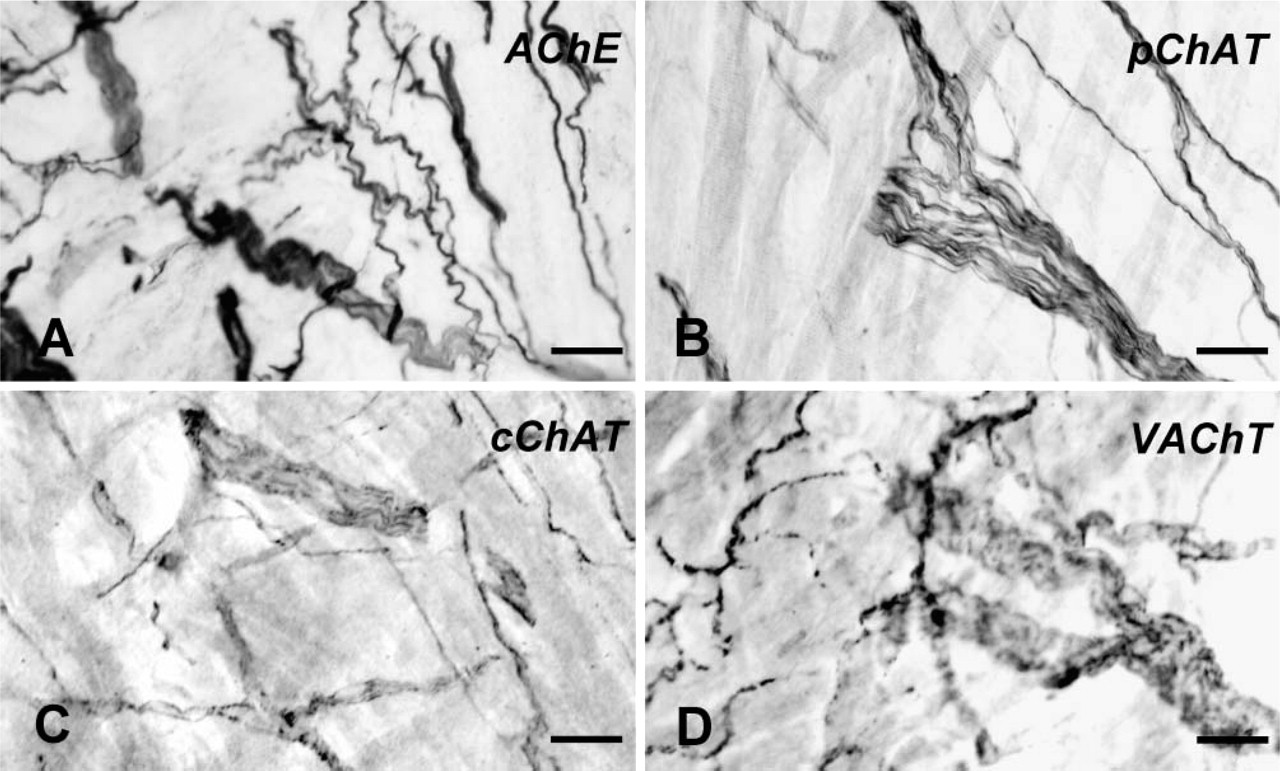
Distribution of the fibers positive for AChE (
The question we should discuss is how cholinergic fibers play roles in ventricular functions. Because of the relative absence of CHT-immunoreactive fibers in the guinea pig left ventricle, it was suggested that vagal efferent nerves are unlikely to exert direct effects on ventricular contractility (Hoover et al. 2004). In contrast, the functional and clinical significance of the parasympathetic nerves in control of ventricular function has been suggested on the ground that VAChT-positive fibers are densely distributed in the ventricles (Arvidsson et al. 1997; Schäfer et al. 1998). To our knowledge, distribution of cChAT-positive fibers in the ventricular walls has never been described.
In the present study we compared distribution patterns of the positive fibers for each cholinergic marker in the ventricular walls. The results confirm that rat ventricular myocardium contains a small number of pChAT-positive fibers. Coronary blood vessels in the ventricular myocardium are also supplied by pChAT-positive fibers. As has been reported (Sequeira et al. 2005), some VAChT-positive fibers reach the wall of coronary blood vessels in the ventricles as well. Our confocal study shows that pChAT and VAChT immunoreactivities are closely associated in some single fibers and in some nerve fiber bundles in the ventricular walls. In mammals such as the dog, cat, and human, a line of evidence indicates that the vagal nerve exerts direct effects on ventricular functions. For example, vagal stimulation decreases ventricular contractility, increases the myocardial interstitial level of ACh, suppresses some ventricular arrythmias, and has a cardioprotective effect independent of the heart rate-slowing mechanism (Vanoli et al. 1991; Schwartz and Zipes 2000; Kawada et al. 2006). In the rat, although no direct influence of vagal stimulation on ventricular contractility has been reported (Takahashi et al. 2003), vagal nerve stimulation has been reported to protect heart against ischemia-induced ventricular arrythmias (Ando et al. 2005). Thus, it is likely that the cholinergic mechanism has regulatory roles on ventricular functions with substantial species differences. The present results provide morphological evidence indicating that cholinergic fibers play roles in ventricular functions, even in the rat. The reason for the paucity of cChAT-positive fibers in the ventricles remains uncertain. It is possible that antigenic epitopes of cChAT are difficult for cChAT antibodies to recognize due to their three-dimensional structure.
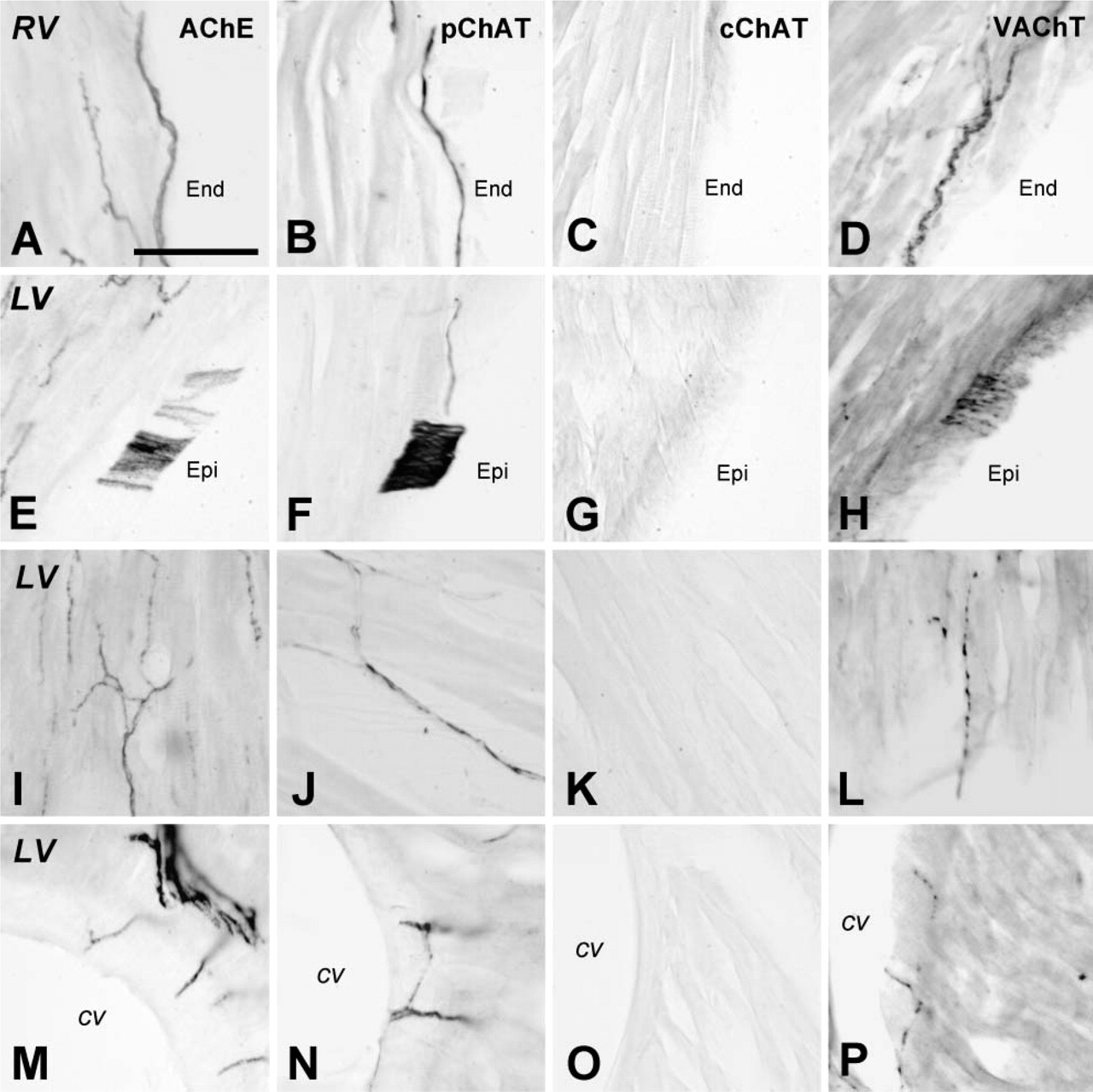
Distribution of putative cholinergic fibers in sections of the ventricular wall of the rat heart. (
Collectively, the present study provides additional evidence indicating that pChAT is a better marker for peripheral cholinergic structures than cChAT. When the staining pattern for pChAT was compared with that for VAChT, it was evident that pChAT immunoreactivity was present preferentially in neuronal somata and smooth fibers, whereas VAChT immunoreactivity was localized to terminal varicose fibers rather than to cell somata or to thick fiber bundles. Although it cannot be ruled out that pChAT also has the same immunodetection problem shared with cChAT, it is possible that the subcellular localization of pChAT in neurons is different from that of VAChT. The proteins for synthesis and storage of ACh, such as cChAT and VAChT, are known to localize to cholinergic varicosities. Therefore, the difference in subcellular localization between pChAT and VAChT might support our above-mentioned speculation that pChAT is responsible for ACh production in distinct intracellular compartments from terminal vesicles. Further analyses on the subcellular localization and molecular kinetics of pChAT will provide much information to clarify its functional significance in neurons.
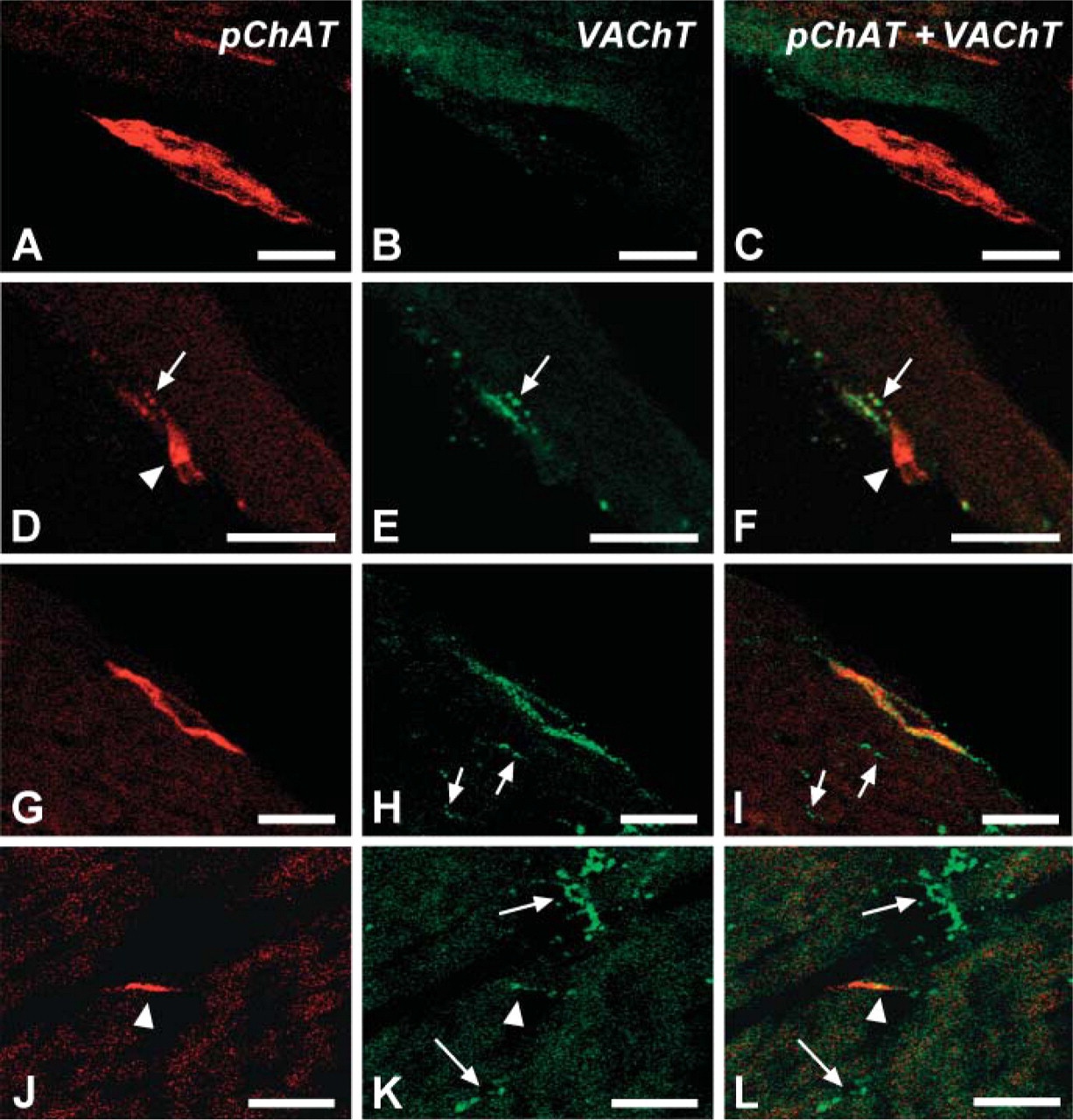
Confocal images of nerve fibers positive for pChAT (red) (
Finally, it should be noted that pChAT-immunoreactive fibers in the heart do not always represent parasympathetic postganglionic fibers. Without colchicine treatment, cholinergic parasympathetic preganglionic fibers are generally unstained with the pChAT antibody (Nakanishi et al. 1999; Yasuhara et al. 2004). However, it is possible that pChAT is localized to some sensory fibers in the heart, as is the case with AChE. AChE activity was observed in sensory neurons in the nodose ganglion of the vagus nerve (Koelle 1955) and in capsaicin-sensitive sensory fibers in the pericardium and atrioventricular valves of the heart (Papka et al. 1981). Similarly, we have reported that sensory neurons in the nodose ganglion display a positive reaction for pChAT (Nakanishi et al. 1999). The cholinergic traits of sensory neurons have also been noted in the dorsal root ganglion and trigeminal ganglion and, accordingly, we and other authors have proposed the concept of the cholinergic sensory system (Sann et al. 1995; Yasuhara et al. 2003,2004; Tata et al. 2004). As mentioned, we have suggested that pChAT has a ChAT enzyme activity (Yasuhara et al. 2003,2004; Bellier JP, et al., unpublished data), supporting the specificity of pChAT as a cholinergic marker. In the heart, however, it is unlikely that pChAT-positive sensory fibers contribute a major part of the positive fibers because the distribution pattern of pChAT-positive fibers appears different from those of substance P-containing sensory fibers (data not shown) and AChE-positive capsaicin-sensitive fibers (Papka et al. 1981). The contribution and significance of pChAT-positive sensory fibers in the heart should be investigated in future studies.
Footnotes
Acknowledgements
We thank Mr. T. Yamamoto, M. Suzaki, and R. Okamoto (Central Research Laboratory, Shiga University of Medical Science, Shiga, Japan) for technical advice and Dr. E.G. McGeer (University of British Columbia, Vancouver, Canada) for critical reading of the manuscript.