
Abstract
We studied the immunohistochemical localization of CD44 and the ezrin–radixin–moesin (ERM) family of actin binding proteins in mouse enamel organ, using con-focal laser scanning microscopy and transmission electron microscopy to clarify their role in cytoskeletal organization. At the differentiation stage of ameloblasts, immunoreactivity to CD44 was detected on the plasma membrane of the inner enamel epithelium, the stellate reticulum, the stratum intermedium, and the external enamel epithelium. In accordance with the differentiation of preameloblasts into secretory ameloblasts, immunoreactivity increased in the stratum intermedium cells. At the maturation stage, intense immunoreactivity was observed on the papillary layer cells. For the ERM family, the stratum intermedium and the papillary layer cells were stained with anti-ezrin and -radixin monoclonal antibodies but not with the anti-moesin antibody. Electron microscopic observations revealed that CD44, ezrin, and radixin were localized in the region at which preameloblasts came into contact with the stratum intermedium at the differentiation stage. At the secretory and maturation stages, they were concentrated in the microvilli of the stratum intermedium and the papillary layer cells. These findings suggest that the CD44–ezrin–radixin–actin filament system is involved in cell–cell interaction between preameloblasts and the stratum intermedium, and in the cytoskeletal organization of the cells in the stratum intermedium and the papillary layer.
Keywords
E
We previously reported that the plasma membranes of stratum intermedium cells and papillary layer cells contain carbohydrates (Nakai et al. 1985; Nakamura and Ozawa 1990), CD44, and heparan sulfate (HS) chains (Nakamura et al. 1995). CD44 (Haynes et al. 1991; Underhill 1992) is believed to function as a hyaluronate receptor because it demonstrates a homology in the extracellular domain with cartilage link and proteoglycan core proteins (Goldstein et al. 1989; Stamenkovic et al. 1989). This molecule is a multifunctional adhesion molecule, binding to Type I collagen, fibronectin, and serglycin (Carter and Wayner 1988; Jalkanen and Jalkanen 1992; Toyama–Sorimachi et al. 1995) and is known to be involved in the attachment of hematopoietic stem cells to stromal cells (Miyake et al. 1990), in lymphocyte homing to Peyer's patches and the high endothelial venules of peripheral lymph nodes (Jalkanen et al. 1987), in cell migration (Jacobson et al. 1984), and in metastatic spread (Günthert et al. 1991). Most adhesion molecules participate in signal transduction through cytoskeleton. Likewise, CD44 was suggested to be directly associated with actin filaments (Tarone et al. 1984) via an ankyrin-like molecule in its cytoplasmic domain (Lacy and Under-hill 1987; Kalomiris and Bourguignon 1988) Recently, Tsukita et al. (1994) reported that the ERM family, consisting of ezrin (Bretscher 1983), radixin (Tsukita et al. 1989), and moesin (Lankes et al. 1988) and belonging to the band 4.1 superfamily (Funayama et al. 1991; Sato et al. 1992; Takeuchi et al. 1994a), might connect CD44 with actin filaments in microvilli at cell–cell and cell–matrix adhesion sites. Although Nishikawa et al. (1990) reported the localization of radixin in secretory ameloblasts, the relationship between CD44 and the ERM family remains to be determined in cells of the enamel organ.
In this study we investigated the localization of CD44 and the ERM family in the mouse incisor enamel organ to clarify their relationships, using monoclonal antibodies directed against mouse CD44, ezrin, radixin, and moesin. (Takeuchi et al. 1994b), by confocal laser scanning microscopy (CLSM), and by transmission electron microscopy (TEM).
Materials and Methods
Animals and Tissue Preparation
Ten 4-week-old male ddY mice, weighing approximately 17 g each, were immunohistochemically investigated. The mice were anesthetized with sodium pentobarbital and perfused through the left ventricle, first with Ringer's solution. Mice used for CD44 detection were perfused with 4% paraformaldehyde, 0.1% glutaraldehyde, and 0.1% tannic acid in 0.05 M phosphate buffer (pH 7.3) for 10 min. For detection of the ERM family, mice were perfused with 4% paraformaldehyde in 0.05 M phosphate buffer. Their mandibles were dissected, immersed in the same fixative for 4 hr at 4C, and decalcified in 4.13% EDTA (pH 7.3) for 5 days at 4C. Incisor enamel organs were used in the following ways.
Immunohistochemical Studies
Cryosections were preincubated in PBS containing 1% BSA for 15 min at room temperature (RT) and then incubated with fluorescein isothiocyanate (FITC)-conjugated rat monoclonal antibody to mouse CD44 (Pharmingen; San Diego, CA) diluted 1:100 for 12 hr at 4C. For the ERM family, sections were incubated with rat monoclonal antibodies against mouse ezrin, radixin, and moesin, (kindly provided by Dr. Tsukita, Kyoto University). After being washed with PBS, they were incubated with FITC-conjugated goat anti-rat IgG (Kirkegaard & Perry Laboratories; Gaithersburg, MD) diluted 1:40 for 1 hr, rinsed with PBS, and examined by CLSM (Olympus, LSM-GB 200; Tokyo, Japan).
Immunocytochemical Studies
Microslicer sections or cryosections were preincubated in PBS containing 1% BSA for 6 hr at 4C and incubated with biotin-conjugated rat anti-mouse CD44 monoclonal antibody (Pharmingen) diluted 1:100 for 24–48 hr at 4C, followed by horseradish peroxidase (HRP)-conjugated streptavidin (Nichirei; Tokyo, Japan) or 10–nm gold-conjugated streptavidin for 24 hr at 4C and washed with PBS. For the ERM family, sections were incubated with rat monoclonal antibodies against mouse ezrin, radixin, and moesin for 24 hr at 4C, followed by HRP-conjugated goat anti-rat immunoglobulins (Biosource International; Camarillo, CA). As for the observation through the HRP method, specimens were first immersed in a DAB solution (0.05% diaminobenzidine in 0.05 M Tris-HCl buffer, pH 7.6), and then in a DAB-H2O2 solution for 10 min at RT. After postfixation with 1% OsO4 in 0.1 M phosphate buffer (pH 7.4) for 1 hr at 4C, the tissue slices were dehydrated in graded acetone and then embedded in Poly/Bed 812 (Polyscience; Warrington, PA). Ultrathin sections were stained with lead citrate and observed under an H-7100 TEM (Hitachi; Tokyo, Japan) at an accelerating voltage of 75 kV.
Controls
Control sections were not incubated with primary antibody, nor were they incubated with normal rat serum.
Results
Immunohistochemical Localization of CD44 in Enamel Organ
At the ameloblast differentiation stage, the enamel organ consisted of four types of cells: pre-ameloblasts, stratum intermedium, stellate reticulum, and outer enamel epithelium. Pre-ameloblasts were column-shaped and faced the odontoblasts. Stratum intermedium cells, roughly polyhedral in shape, were located at the proximal end of the pre-ameloblasts and formed two or three lines. Immunoreactivity to CD44 was detected on the plasma membrane of all cells in the enamel organ (Figure 1a). Preameloblasts and the stratum intermedium showed stronger reactivity than other cells. From the late differentiation stage to the early secretory stage, immunoreactivity on the stratum intermedium gradually increased. At the secretory stage, the staining was most pronounced on the stratum intermedium in the enamel organ (Figure 1b). At the maturation stage of ameloblasts, the stratum intermedium, stellate reticulum, and outer enamel epithelium turned into irregularly shaped polygonal cells, called papillary layer cells. These three types of cells were no longer discernible as separate layers. Papillary layer cells exhibited intense immunoreactivity (Figures 1c and 1d). Minimal immunoreactivity was detected on the lateral plasma membrane of ruffle-ended ameloblasts (RA) and smooth-ended ameloblasts (SA). The ruffled border of RA and the distal plasma membrane of SA did not appear to show any immunoreactivity (Figures 1c and 1d).
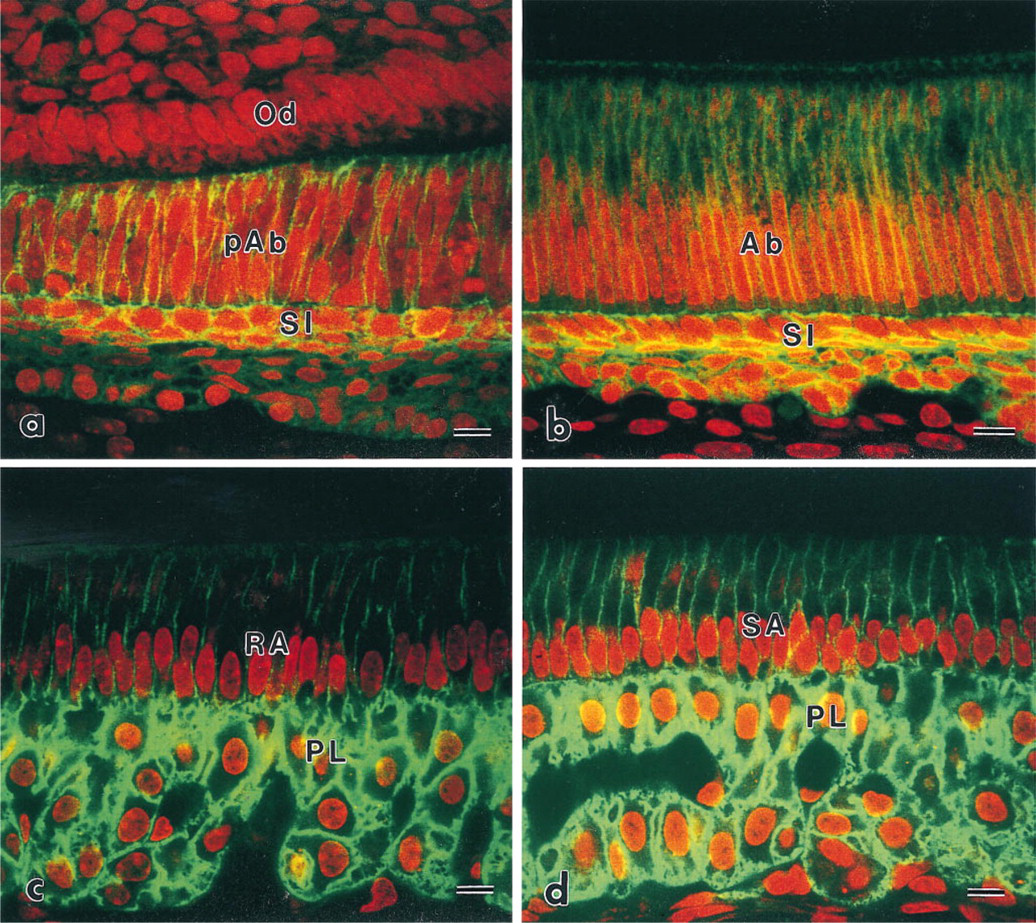
Confocal laser scanning micrographs indicating the localization of CD44. (
Immunohistochemical Localization of the ERM Family in Enamel Organ
Immunoreactive patterns of ezrin and radixin in the stratum intermedium and the papillary layer were similar to each other (Figures 2 and 3). At the differentiation stage, immunoreactivity to ezrin and radixin was restricted to the region of contact between pre-ameloblasts and the stratum intermedium (Figures 2a and 3a). The intensity was gradually increased on two or three layers of the stratum intermedium cells. The stratum intermedium cells at the secretory stage and the papillary layer at the maturation stage were strongly immunolabeled by anti-ezrin and anti-radixin antibodies (Figures 2b–2b and 3b–3d). The lateral plasma membrane of SA and RA also showed weak reactivity to radixin and moesin. The ruffled border of RA also showed immunoreactivity to the ERM family. No apparent reactivity to moesin was seen on the cells of the stratum intermedium and the papillary layer, although blood capillaries showed immunoreactivity to moesin (Figure 4). Odontoblasts showed immunoreactivity to radixin and moesin (Figures 3a and 4a).
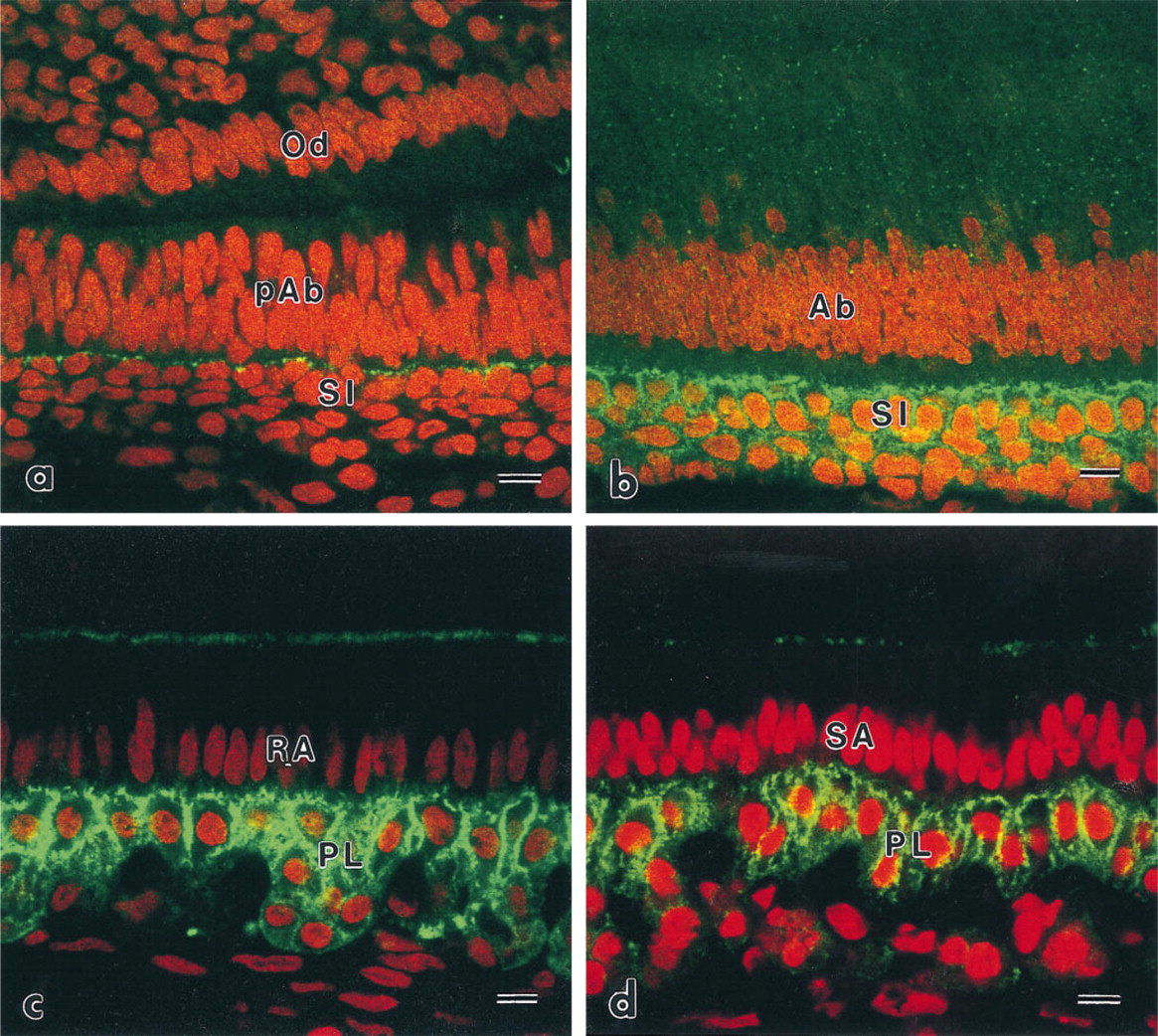
Confocal laser scanning micrographs indicating the localization of ezrin. (
Immunocytochemical Localization of CD44 and the ERM Family
At the differentiation stage, immunoreactivity to CD44 was observed on the plasma membranes of ameloblasts and stratum intermedium. Reactivity was also detected in the region at which the stratum intermedium cells and pre-ameloblasts came into contact with each other (Figure 5a). Electron microscopy showed that ezrin and radixin were localized in the cytoplasmic processes of pre-ameloblasts where they came into contact with the stratum intermedium (Figure 5b). At the secretory and the maturation stage, the plasma membranes of the stratum intermedium and the papillary layer cells were intensely immunolabeled by anti-CD44 monoclonal antibody (Figures 6a and 6b). In addition, gold particles indicating localization of CD44 were mainly detected on the extracellular side of the plasma membrane of the microvilli, containing well-developed actin filaments (Figure 7a). Immunoreactivity to ezrin and radixin was seen in the cytoplasm of the microvilli (Figure 7b).
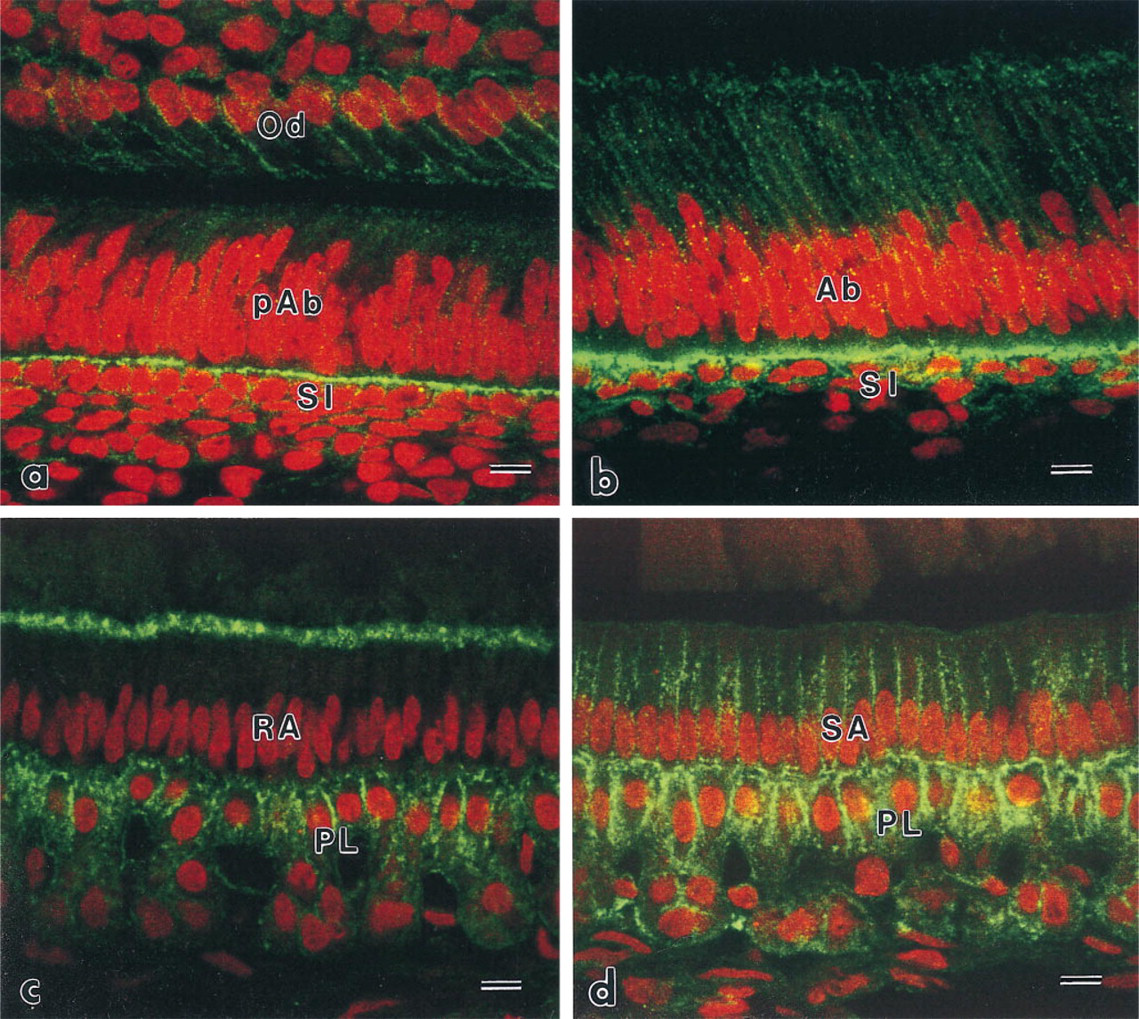
Confocal laser scanning micrographs indicating the localization of radixin. (
Control
No specific immunoreactivity was observed in control sections incubated without any primary antibody or with normal rat serum.
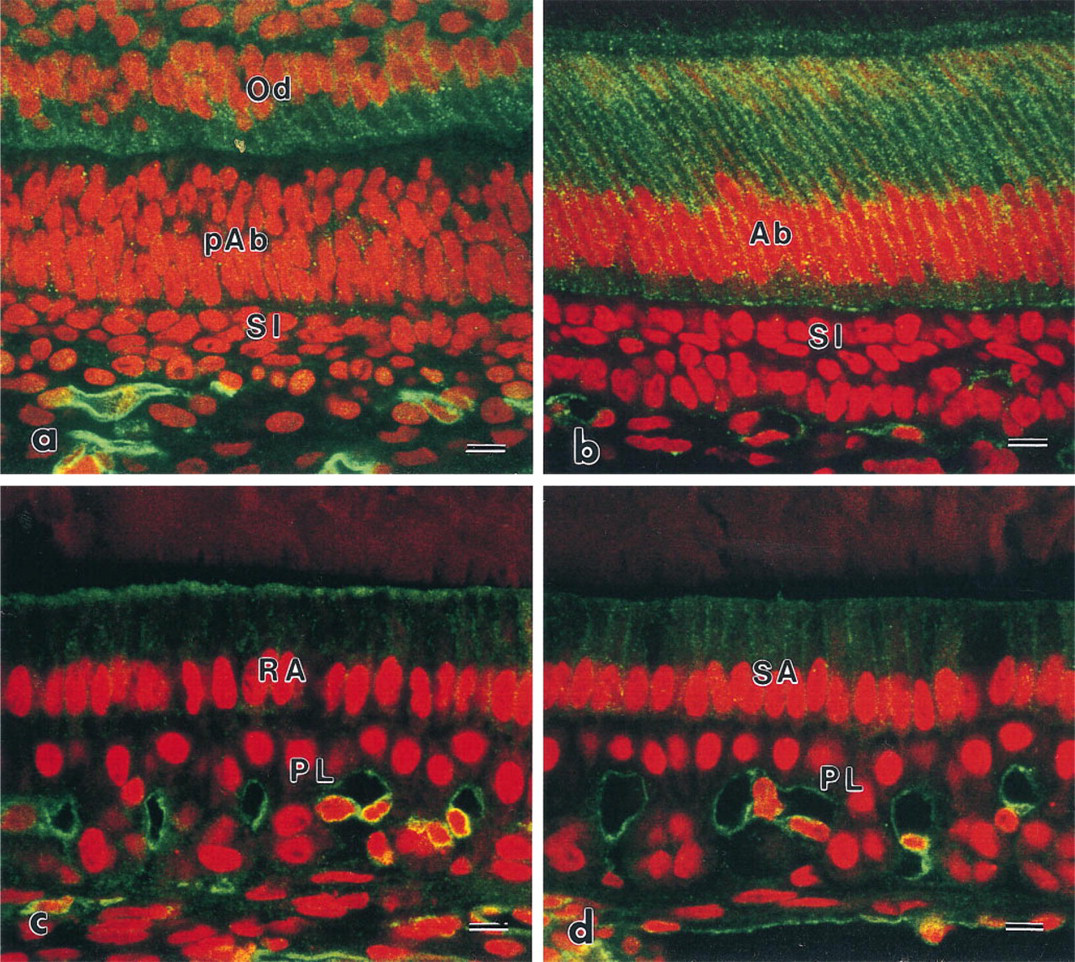
Confocal laser scanning micrographs indicating the localization of moesin. (
Discussion
We used mouse incisor enamel organ in this study because monospecific monoclonal antibodies to the mouse ERM family now available. We confirmed that CD44 was also localized in epithelial cells of the mouse incisor enamel organ, as previously reported in rats (Nakamura et al. 1995). Immunoreactivity in the stratum intermedium and the papillary layer was increased according to the differentiation of ameloblasts. They may intensely express CD44 on their plasma membranes according to their morphological changes. Although the confocal microscopic images appear to show some labeling in the intercellular space, this is apparently due to strong immunoreactivity in the papillary layer and a problem with the resolution of the confocal laser microscope. We also ascertained by electron microscopy that CD44 was mainly localized in the plasma membranes. CD44 is one of the hyaluronate receptors (Goldstein et al. 1989; Stamenkovic et al. 1989). Hyaluronates are believed to play an important role in the development of various kinds of tissue (Knudson and Toole 1987), although the precise mechanism is not yet understood. Preliminary results obtained by using a biotinylated hyaluronic acid binding region show that hyaluronates are localized in the satellite reticulum and the papillary layer (data not shown). CD44 in the enamel organ may be involved in the storage of hyaluronates. The development of tissue is believed to be regulated by systemic hormones and local growth factors as well as cell–cell or cell–matrix interactions. TGF-β1 (Cam et al. 1990; Vaahtokari et al. 1991; Jepsen et al. 1992), BMP-2, −4 (Vainio et al. 1993), hepatocyte growth factor (Sonnenberg et al. 1993; Tabata et al. 1996), and midkine (Mitsiadis et al. 1995) have been suggested as important local growth factors in the development of the enamel organ. We previously reported that CD44, as well as syndecan-1 (Bai et al. 1994), in the stratum intermedium and the papillary layer may also be membrane-type HSPGs (Nakamura et al. 1995). Our previous report also revealed that basic fibroblast growth factor, one of the heparin binding growth factors, can be bound to their plasma membranes via HS chains. Other reports also suggest that syndecan-1 may regulate epithelial–mesenchymal interaction during tooth development (Thesleff et al. 1988; Vainio et al. 1989). Therefore, HS chains of CD44 and syndecan-1 in the stratum intermedium and the papillary layer may also be involved in the regulation of ameloblast differentiation by preserving heparin binding growth factors, such as BMPs, hepatocyte growth factor, and midkine.
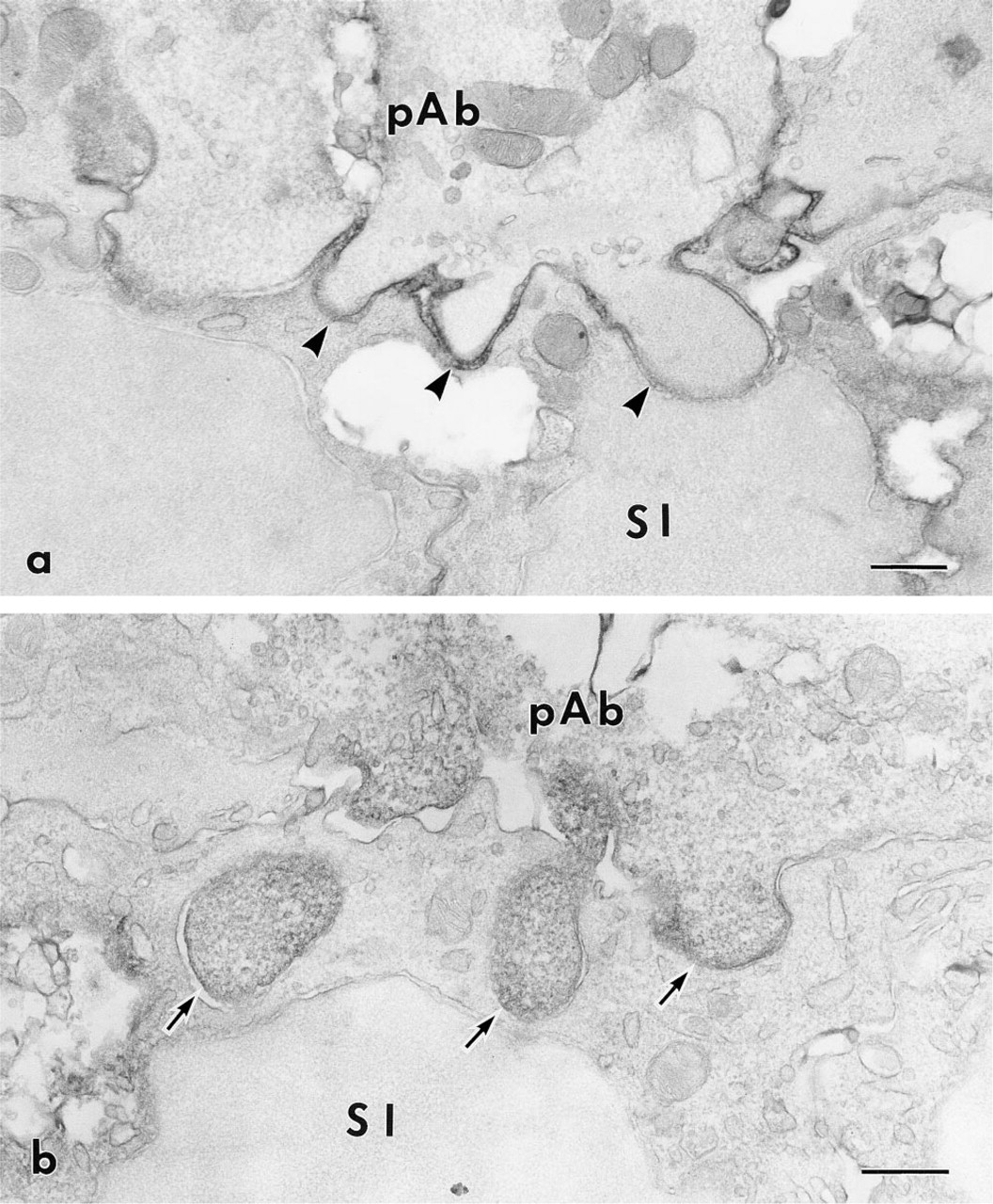
Electron micrographs indicating the localization of CD44 and radixin at the differentiation stage of ameloblasts. (
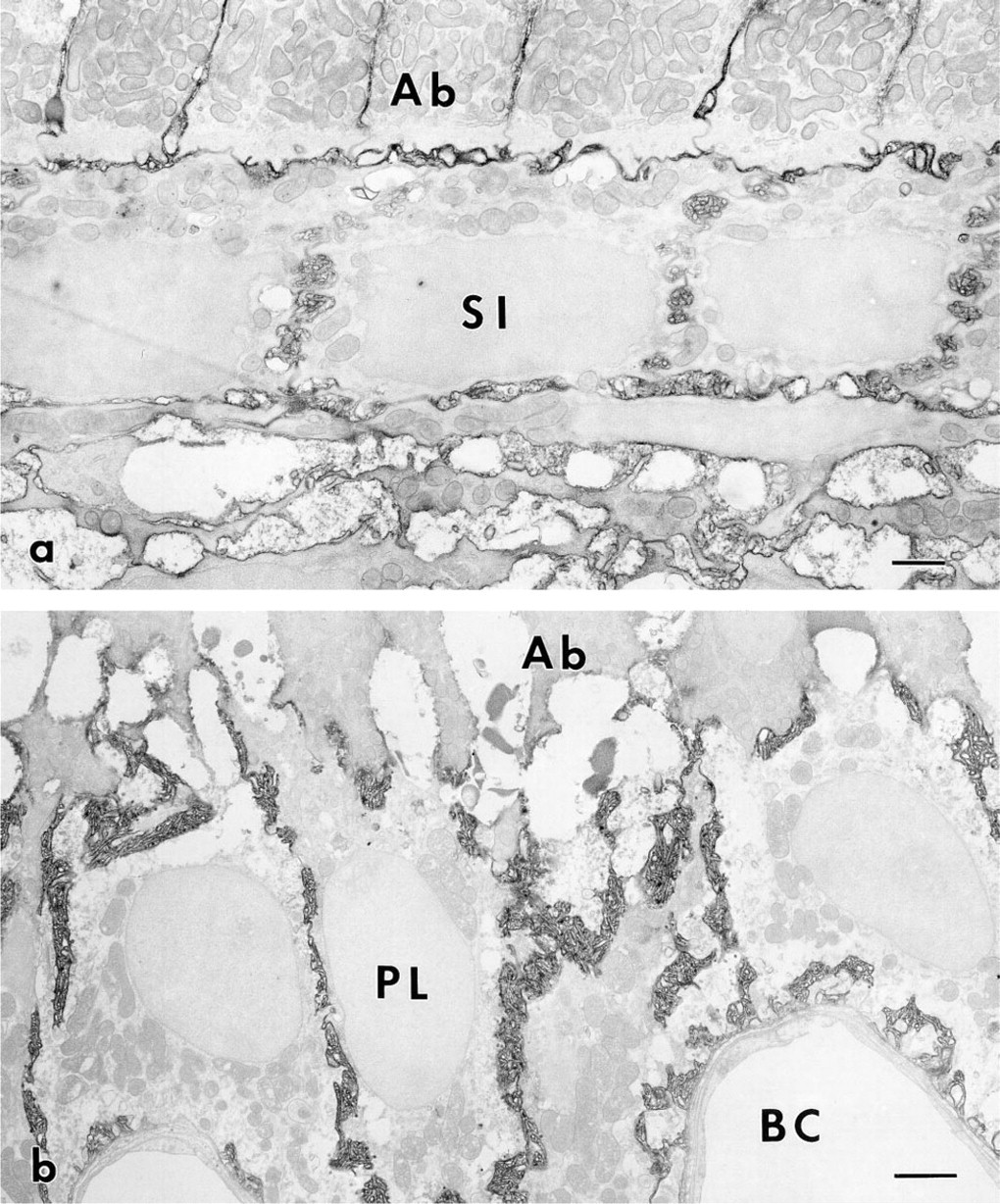
Electron micrographs indicating the localization of CD44 at the secretory stage and the maturation stage. (
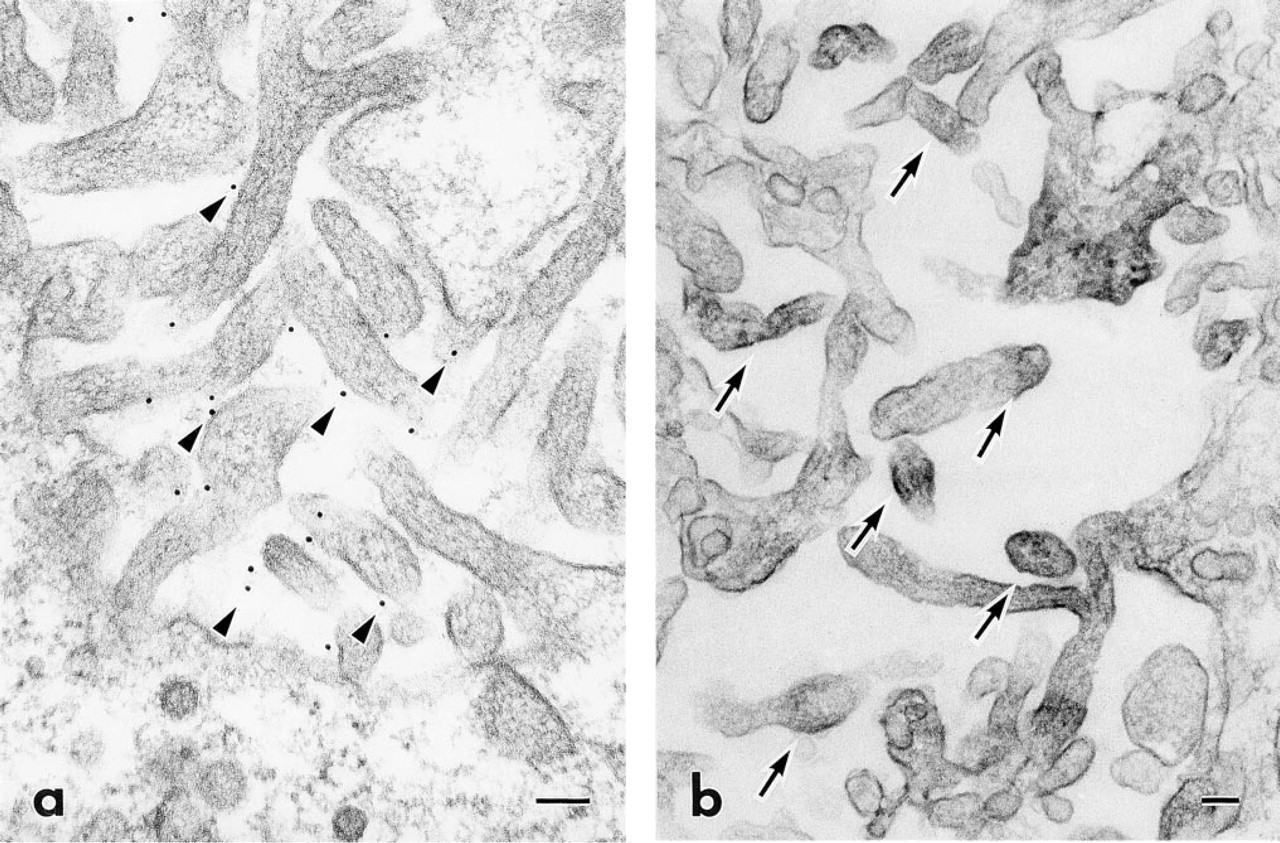
High-magnification electron micrographs indicating the localization of CD44 and radixin in microvilli of the papillary layer. (
The stratum intermedium and the papillary layer have been believed to be involved in the transport of mineral and organic materials because of their characteristics, such as alkaline phosphatase (Ten Cate 1962; Kurahashi and Yoshiki 1972), Ca-ATPase (Sasaki and Garant 1986; Takano et al. 1986), Na,K-ATPase (Ga-rant et al. 1987) on their plasma membranes, well-developed mitochondria, and microvilli. These characteristics are commonly found in cells participating in transport functions. HS chains would also bear a net negative charge because they contain sulfated (-SO3 -) glucosamines. Negatively charged plasma membrane could trap calcium ions and positively charged organic materials. Moreover, autoradiographic reports, using 45Ca (Bawden and Wennberg 1977; Kawamoto and Shimizu 1987), and the histochemical calcium-detecting study using potassium pyroantimonate (Ozawa et al. 1984), suggest that cations are caught by the stratum intermedium and papillary layer. The stratum intermedium and the papillary layer might therefore play a role in the mineral transport system because of their charged plasma membranes.
Inner and outer enamel epithelia form a distinct multiple layer at the secretory stage, consisting of secretory ameloblasts, the stratum intermedium, the stellate reticulum, and the outer enamel epithelium. At the maturation stage, the latter three layers become the relatively homogeneous papillary layer, with an extensive network of microvilli. The ameloblast layer also shows morphological alterations, such as and ruffle- and smooth-ended ameloblasts. All of these events necessitate extensive rearrangements of the cell membrane and cytoskeleton. Recently, the cytoplasmic domain of CD44 was believed to bind to actin filaments via either the ERM family (Tsukita et al. 1994) or ankyrin-like molecules (Lacy and Underhill 1987; Kalomiris and Bourguignon 1988). To clarify the relationship between CD44 and the ERM family in the enamel organ, we observed the immunolocalizations of ezrin, radixin, and moesin by using three kinds of monospecific monoclonal antibodies (Takeuchi et al. 1994b). Although the co-localization of CD44 and the ERM family was not always seen in all cells of the enamel organ, we showed here that CD44 co-localized with ezrin and radixin in the microvilli, which has well-developed networks of actin filaments, of the stratum intermedium and the papillary layer. However, moesin was not localized in these cells, although immunoreactivity of moesin was seen in blood capillaries. These results raise the possibility that the CD44–ezrin, radixin–actin filament system may play an important role in the organization of actin filaments in their microvilli. Expression of both ezrin and radixin in these cells may represent redundant systems that could compensate for the loss of such proteins, as reported for lymphocytes (Takeuchi et al. 1994b). Furthermore, the co-localization of CD44, ezrin, and radixin was seen in the contact sites between pre-ameloblasts and the stratum intermedium. The evidence that pre-ameloblasts in enamel-free areas of mouse molars cannot differentiate into secretory ameloblasts without the support of the stratum intermedium (Sakakura et al. 1989) suggests that cell–cell communication and/or interaction is essential for differentiation of ameloblasts. Adhesion molecules and their signal transductions via cytoskeletons are believed to be important for the differentiation and the polarity of cells. In addition, the ERM family is known to be a good substrate for tyrosine kinase (Bretscher 1989; Gould et al. 1989) and to be associated with rho small GTP-binding proteins (Takaishi et al. 1985). Although the precise mechanism is not understood, the CD44–ezrin–radixin–actin filament system may be important for cell–cell interaction between pre-ameloblasts and the stratum intermedium by providing mechanical support and signal transduction via CD44 and the cytoskeleton, such as phosphorylation of the ERM family, to understand cell–cell interaction in the enamel organ.
Adhesion and cytoskeletal changes among secretory ameloblasts are believed to be related to their sliding and the production of sinuous enamel prisms (Warshawsky and Smith 1971; Risnes 1979; Nishikawa 1992). We were not convinced of the co-localization of CD44 and the ERM family in secretory ameloblasts. Although other layers in the enamel organ, the stratum intermedium and the stellate reticulum, may directly or indirectly induce the moving force of secretory ameloblasts, it requires further investigation as to whether the CD44–ezrin–radixin–actin filament system is involved in their sliding.
In conclusion, the CD44–ezrin–radixin–actin filament system is involved in cell–cell interaction between pre-ameloblasts and the stratum intermedium and in the cytoskeletal organization in the cells of the stratum intermedium and the papillary layer.
Footnotes
Acknowledgements
Supported in part by a research grant from the Ministry of Education, Science, Sports and Culture of Japan.
We would like to thank all the members of our laboratory (1st Department of Oral Anatomy, Niigata University School of Dentistry) for assistance, especially S. Ejiri for helpful discussions and Ms S. Maekawa for expert secretarial assistance. We are also grateful to Dr Sh. Tsukita (Department of Cell Biology, Faculty of Medicine, Kyoto University) for his generous gift of ant-mouse ERM monoclonal antibodies.