
Abstract
The oligosaccharides of the mucous gastric glycoproteins are involved in the protection of the gastric mucosa and are altered in different diseases. Therefore, it is important to know their composition in health, to better determine the alterations induced by the disease. Moreover, analysis of the molecular composition of the fundic gland cells has been previously used to obtain new insights into the origin of the different cell types. The aim of the present study was the localization in the subcellular structures of the fucose residues of the oligosaccharides in human fundic glands. For this, lectin cytochemical methods were used at the light and electron microscopic levels. They were combined with enzymatic and chemical treatments to characterize the nature of the oligosaccharide chains containing the fucose residues. The presence of this carbohydrate belonging to N- or O-linked oligosaccharides has been demonstrated in the secretory granules of the surface, gastric pit, mucous neck, and transitional cells of the fundic mucosa, and in the intracellular canaliculi and tubulovesicular system of the parietal cells. These fucose residues were added in the trans-Golgi regions to the elongating chains. Additional fucose linked to the innermmost N-acetylglucosamine of the N-linked oligosaccharides was found in the chief cells, being incorporated in the cis-Golgi. The findings in the transitional cells corroborate the origin of the chief cells from the mucous neck cells.
T
The mucous glycoproteins are secreted by the mucous cells of the epithelium and the gastric glands. In the human fundic region of the stomach, the glands are composed of mucous neck cells, chief cells, parietal cells, and endocrine cells. More recently, the existence of transitional or prezymogen cells has been described (Rubin et al. 1968; Cornaggia et al. 1986; Karam and Leblond 1993a, b; Ogata 1997; Yang et al. 1997; Madrid et al. 1990,1998b). These transitional cells can be considered as an intermediate stage in the hypothetical differentiation process from the mucous neck cells to the chief cells. In this way, the study of the molecular composition of this cell type will contribute to the understanding of its role in the human fundic glands.
The aim of the work was the ultrastructural localization of fucose (Fuc) residues in the human fundic glands by lectin cytochemistry to obtain new insights into their role in the formation of the mucous barrier and on the possible role of the transitional cells in chief cell differentiation. Chemical and enzymatic procedures were used to investigate the N- and/or O-link of the fucosylated oligosaccharides to the protein core. For this, three Fuc-specific lectins were used at both light and electron microscopic levels: Aleuria aurantia agglutinin (AAA), Lotus tetragonolobus agglutinin (LTA), and Ulex europaeus agglutinin-I (UEA-I).
Materials and Methods
Reagents
Polyethylene glycol (mw 20,000) and tetrachloroauric acid were obtained from Merck (Darmstadt, Germany), UEA-I, Fuc, 3,3′-diaminobenzidine (DAB), bovine serum albumin (BSA), LTA-, UEA-I-horseradish peroxidase (-HRP) conjugates, and anti-mouse immunoglobulin IgG+M-HRP goat antibody were purchased from Sigma (Poole; Dorset, UK). Anti-A, anti-B, and anti-H blood group mouse monoclonal antibodies (mAbs) were from Dako (Santa Barbara, CA). Endob-β-N-acetylglucosaminidase F/peptide N-glycosidase F from Flavobacterium meningosepticum, AAA-digoxigenin (DIG)-labeled lectin, unlabeled anti-DIG mouse antibody, unlabeled anti-DIG sheep antibody, DIG antibody labeling kit, and anti-DIG-HRP-labeled goat antibody were purchased from Boehringer Mannheim (Barcelona, Spain). Goat anti-mouse IgG+M-gold (15-nm) and anti-sheep IgG-gold complexes (15-nm) were from BioCell (Cardiff, UK).
Monodisperse colloidal gold solution (mean diameter 20 nm) was prepared according to Frens (1973) and the mean particle diameter was checked by means of an IMCO 10 image analyser computer. UEA-I-gold conjugates were prepared as reported previously (Castells et al. 1992).
LTA-DIG and UEA-I-DIG complexes were prepared by mixing 1 mg of the lectins with 0.044 mg DIG-NHS according to the labeling kit from Boehringer Mannheim Biochemica. The lectin-DIG complexes were purified with a Sephadex G-25 column and the samples with a higher absorbance at 280 nm were selected.
Lectin Specificity
The three lectins recognize Fuc residues. However, their binding affinity can change depending on the carbohydrates and bond type to which Fuc residues are linked. LTA and UEA-I preferentially label Fuc residues located in the outer regions of the oligosaccharide chains, whereas AAA can also bind to those Fucα(1-6)-linked to the innermost GlcNAc residues of the core of N-linked oligosaccharides (Pereira and Kabat 1974; Sugii and Kabat 1982; Osawa and Tsuji 1987).
Tissue Samples and Preparation
Samples of histologically normal fundic mucosa were obtained from 10 cases of total or partial gastrectomy for duodenal ulcers. The normal histology of the tissues was checked by light microscopy.
For light microscopy, the specimens were fixed in 10% formalin in PBS, pH 7.4, for 6 hr. Then they were embedded in paraffin and 5-μm-thick sections were obtained.
For electron microscopy, the tissues were fixed in 2% glutaraldehyde in PBS for 2 hr. After washing, the specimens were immersed in 0.5 M NH4Cl for 1 hr to block free aldehyde groups. Osmium tetroxide postfixation was avoided to preserve antigenicity and lectin reactivity. Then, after washing in PBS, the specimens were embedded in Lowicryl K4M (Carlemalm et al. 1982). Ultrathin sections were mounted on formvar-carbon-coated nickel grids.
Lectin Labeling
Light Microscopy. For cytochemical labeling of the sections, two procedures were used. A one-step method was used for UEA-I- and LTA-HRP-labeled lectins, as reported previously (Madrid et al. 1989). A two-step method was used with AAA-DIG-labeled lectin according to Sata et al. (1990), by using anti-DIG-HRP-labeled goat antibody as second layer. In both procedures, HRP was developed with DAB and hydrogen peroxide and the sections were slightly counterstained with hematoxylin.
Electron Microscopy. For the ultrastructural study, colloidal gold was the selected marker, using two different procedures. UEA-I-gold (1:10) complexes were used in a one-step method as previously described (Madrid et al. 1990,1994). The second method was UEA-I-DIG (1:4), LTA-DIG (undiluted), and AAA-DIG (20μg/ml) in combination with either unlabeled anti-DIG mouse antibody (20μg/ml) or anti-DIG sheep antibody (7μg/ml). The corresponding third layer was goat anti-mouse IgG+M-gold conjugate (1:10) and donkey anti-sheep IgG-gold complex (1:10), respectively (Leis et al. 1997).
Controls
The following controls were used: (a) substitution of conjugated and unconjugated complexes (lectins and antibodies) by the corresponding buffer; and (b) preincubation of the lectins with the corresponding hapten sugar inhibitor (Fuc) at a concentration of 0.2 M and the antibodies with the corresponding antigen.
Immunocytochemical Labeling
To determine the blood group of the specimens and their secretor status, the tissues were stained with anti-A, anti-B, and anti-H antibodies. Because this is not the subject of this work, the method will not be described. The sections were stained according to Madrid et al. (1994, 1998b).
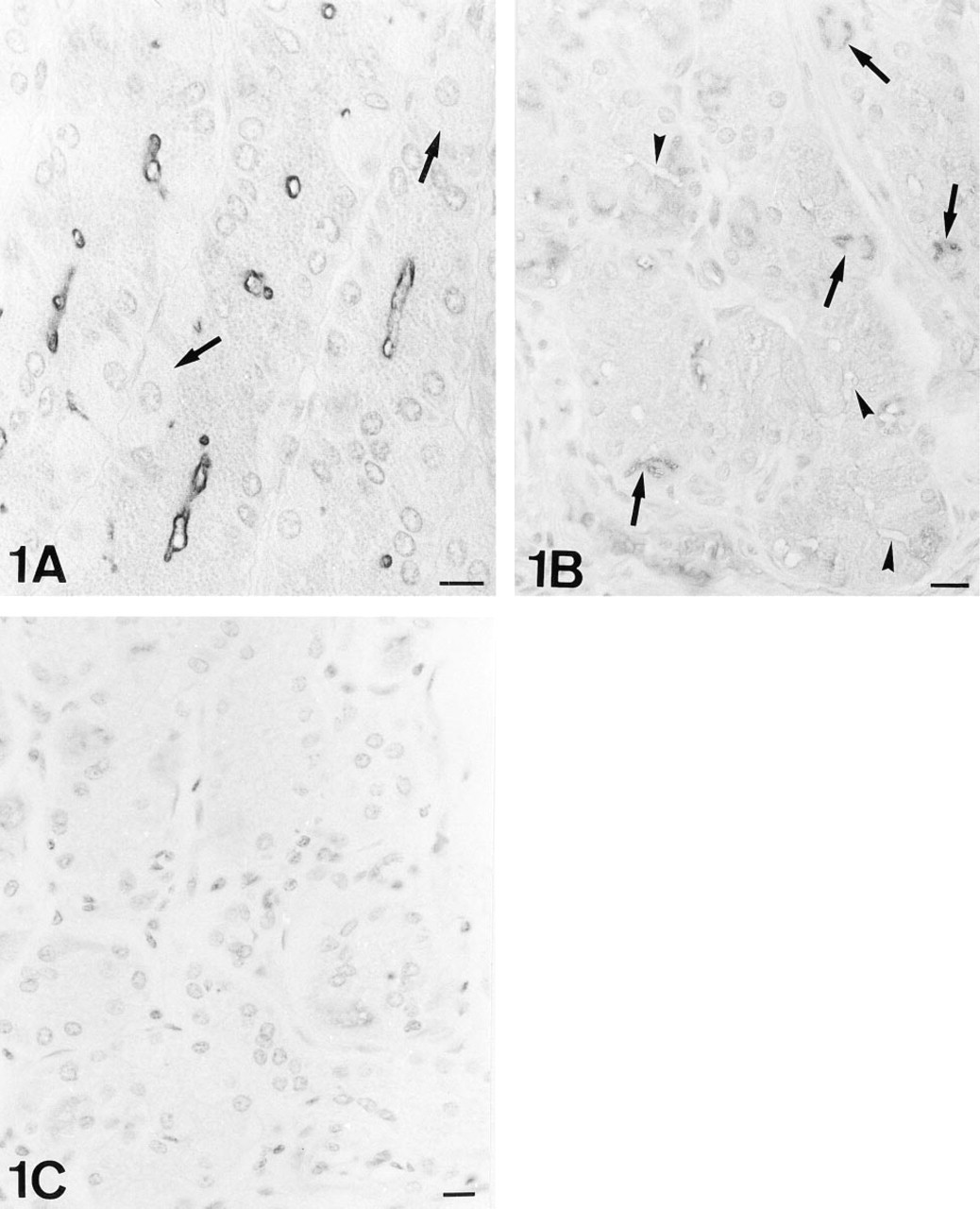
UEA-I-HRP. Note the labeling on the luminal surface of the chief cells. The parietal cells of this microscopic field are negative (arrows). Nomarski interference optics.
Removal of O- and N-linked Oligosaccharides
β-Elimination is a chemical method that selectively removes the O-linked oligosaccharides from the tissue (Ono et al. 1983). The sections were incubated in 0.5 N NaOH in 70% ethanol at 4C for 14 or 21 days. Sections of rat small intestine stained with Helix pomatia agglutinin (HPA) were used as controls. We have previously demonstrated that when the β-elimination procedure is correctly performed, the rat Paneth cells lose their characteristic HPA staining (Leis et al. 1997).
The enzyme endo-β-acetylglucosaminidase F/peptide N-glycosidase F was used as previously reported (Lucoq et al. 1987; Martínez-Menárguez et al. 1993) at pH 9. At this pH, peptide N-glycosidase F (PNGase-F) predominated over endo-β-acetylglucosaminidase F activity, so that cleavage of β-aspartyl glycosyl linkage (GlcNAc β 1-N-Asn) was preferred. In this manner, N-linked oligosaccharides were removed from the peptide core. Rat testis sections were used as control. PNGase-F activity renders these rat testes negative to AAA (Martínez-Menárguez et al. 1993).
Results
Blood Groups and Secretor Status of the Samples
The blood group of the samples was elucidated from the reactivity of the anti-blood group antibodies in the vessels and the erythrocytes. The secretor status was determined by the reactivity of these antibodies in the gastric secretions. The lectins used showed affinity for the H-antigen. Therefore, the expression of the H-antigen in tissues from secretor individuals could induce lectin reactivity in these tissues. To avoid this interaction, non-O blood group individuals were selected for the study. The selected cases were four A secretors, three A non-secretors, and three B non-secretors.
Lectin Cytochemical Localization of the Fuc Residues
UEA-I. The surface and gastric pits cells were labeled both in the mucous granules and the apical membrane (Figure 2). The Golgi apparatus was unlabeled or showed a few gold granules in the trans face or in vesicles exiting from it (Figure 2). No modification of the binding pattern was observed with the β-elimination or PNGase-F treatments.
The transitional and mucous neck cells were also positive in the cytoplasm and the apical membrane, but the β-elimination pretreatment abolished the staining in the luminal surface. Their secretory granules were labeled in the electron-lucent areas (Figure 3), although the intensity was lower in the transitional cells. However, after PNGase-F treatment, some labeling was also found on the electron-dense core. The Golgi apparatus was negative, but some gold granules were present in the vesicles exiting from it.
The chief cells were reactive only in the luminal surface, which became negative with β-elimination (Figures 1A and B).
The parietal cells were negative (Figure 1A), and only occasionally the intracellular canaliculi and apical membrane were stained. Immature parietal cells are characterized by the presence of electron-lucent granules, resembling mucous granules. These cells were labeled in the apical membrane. After PNGase-F treatment, some lectin labeling sites were localized on the intracellular canaliculi and the tubulovesicular system of parietal cells, and in the granules of immature parietal cells. After β-elimination, the intracellular canaliculi were strongly positive (Figure 1B).
LTA. The surface and gastric pit cells were slightly positive in the apical membrane, trans-Golgi and secretory granules, and in the mucous neck and the transitional cells. The secretory granules of the mucous neck cells were labeled in the electron-lucent areas. After PNGase-F treatment, reactivity was also found on the electron-dense core of the granules. The transitional cells showed a binding pattern similar to the mucous neck cells, except for the electron-dense regions, which were slightly reactive to the lectins. The intensity of labeling increased according to the size of the electron-dense region; the larger ones showed a higher labeling (Figure 4). After PNGase-F treatment, the labeling in the electron-dense core was different, depending on size. The labeling in the smaller electron-dense cores (a similar size to that of the mucous neck cells) was increased (similarly to that of the mucous neck cells), whereas in the larger electron-dense regions no significant change was found.
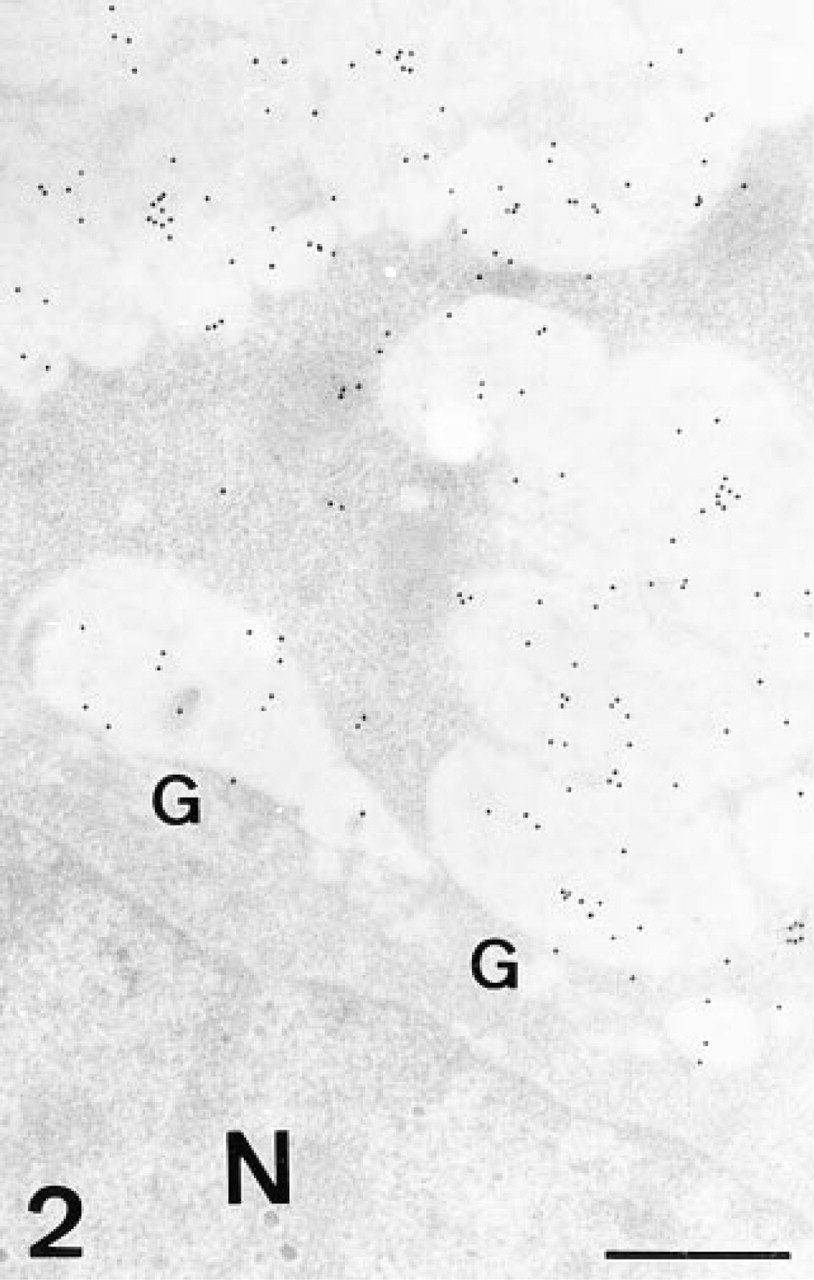
UEA-I, gastric pit cell. Gold granules label the secretory granules and the vesicles exiting from the trans-Golgi face. G, Golgi stack; N, nucleus. Bar = 0.5 μm.
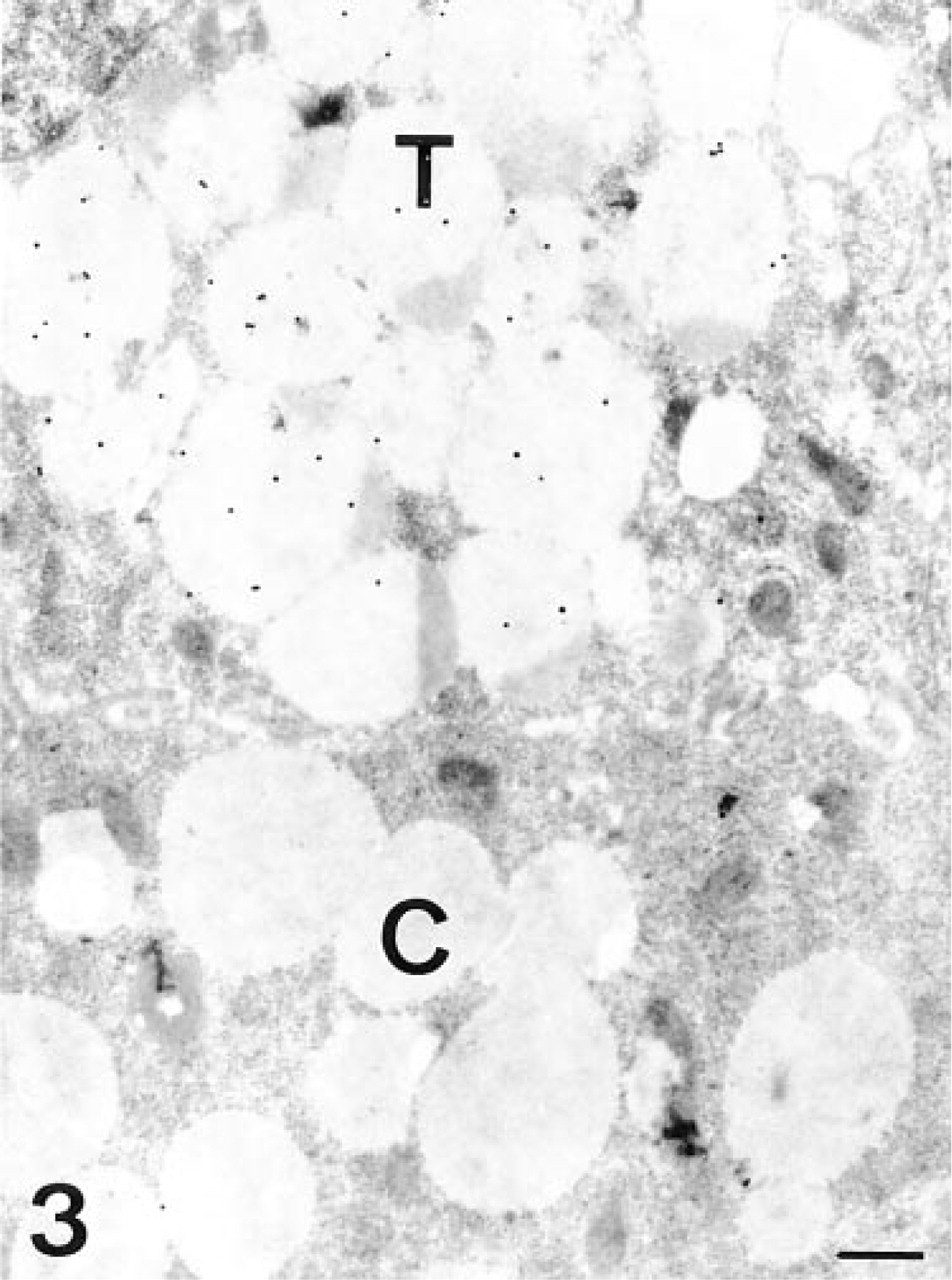
UEA-I, transitional and chief cells. The chief cell (C) is unlabeled, whereas the transitional cell (T) shows some gold granules on the electron-lucent regions of its secretory granules. Bar = 0.5 μm.
The chief cells were labeled in the apical membrane. Some gold granules were occasionally observed on the secretory granules, close to the membrane. The only change observed after PNGase-F was the appearance of slight labeling on the content of the secretory granules.
The parietal cells were labeled in the apical membrane, in the intracellular canaliculi, and in the tubulovesicular system. In the immature parietal cells, some gold granules were deposited on the electron-lucent granules, usually close to their membrane. No significant changes were observed with PNGase-F.
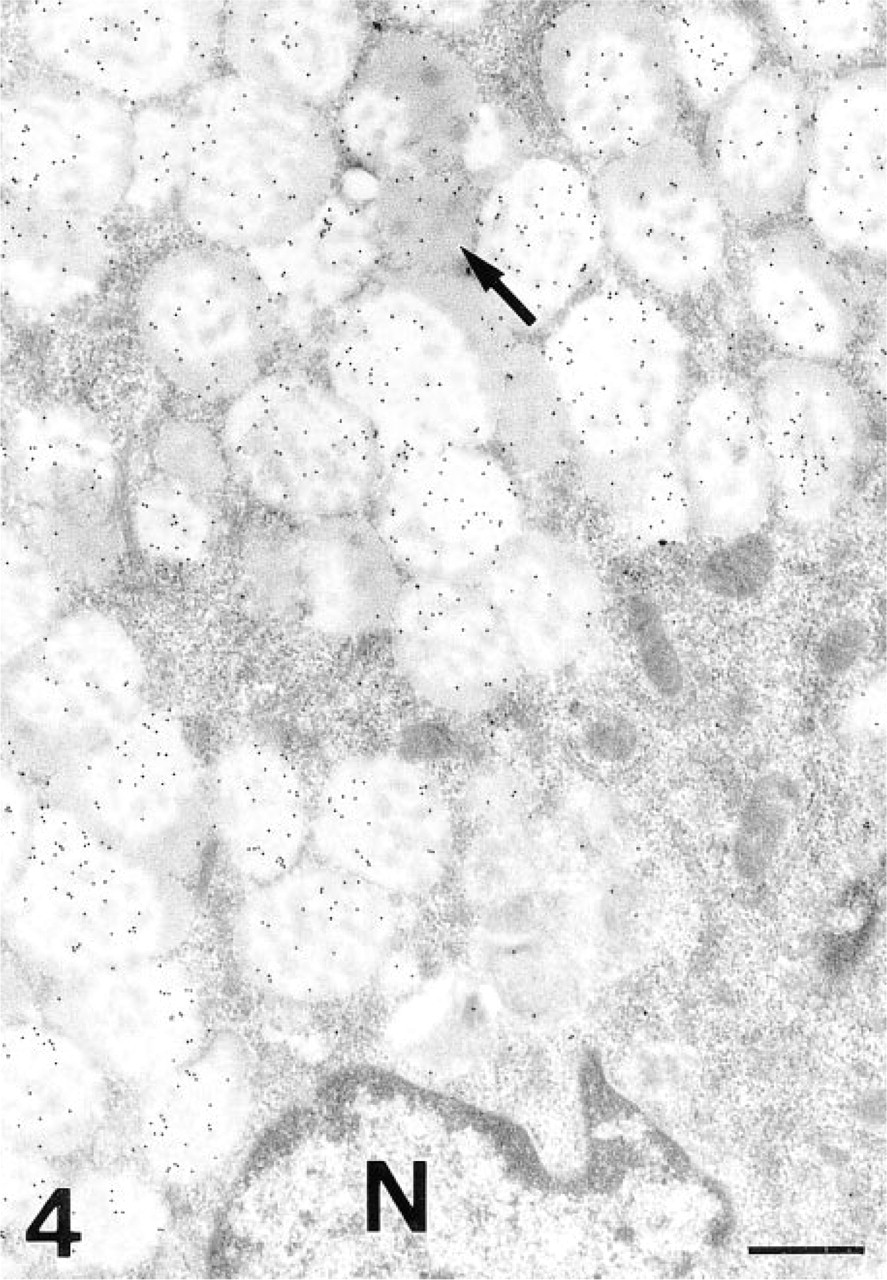
LTA, transitional cell. The gold granules are deposited on the electron-lucent regions of the secretory granules. Some electron-dense areas are also labeled (arrow). N, nucleus. Bar = 0.5 μm.
β-Elimination pretreatment abolished the LTA-staining.
AAA. The surface and pit cells were strongly labeled in the secretory granules. Labeling was also observed in the apical membrane and trans-Golgi. After PNGase-F, only a very slight decrease in the labeling was observed.
The mucous neck cells were strongly labeled in the electron-lucent regions of the secretory granules and to a lesser degree in the electron-dense ones. The apical membrane was reactive. The Golgi apparatus was labeled from the cis to the trans cisternae. After PNGase-F, a decrease in both regions of the secretory granules was observed.
The transitional cells showed labeling in the secretory granules in both the electron-dense and the electron-lucent regions (Figure 5). The cisternae of the Golgi apparatus and the apical membrane were also labeled. After PNGase-F treatment, the reactivity decreased in both regions of the secretory granules and in the apical membrane (Figure 6).
The granules of the chief cells were positive for the lectin (Figure 7), as was the apical membrane. All the cisternae of the Golgi stacks were labeled by the lectin (Figure 7). When PNGase-F was used, the labeling in the Golgi apparatus and secretory granules almost disappeared (Figure 6). The labeling in the apical membrane also decreased.
The intracellular canaliculi and tubulovesicular system of the parietal cells were positive for the lectin (Figure 8). After PNGase-F, slight labeling remained. The electron-lucent granules observed in the immature parietal cells showed a similar binding pattern.
The β-Elimination procedure did not significantly modify the lectin binding pattern. Control sections were always negative for the three lectins at light (Figure 1C) and electron microscopic levels.
Discussion
Localization of Fucosylated Oligosaccharides
The three lectins identified Fuc residues in the secretory granules of the surface, gastric pit, mucous neck, and transitional cells. In these last two cells, labeling was observed over the electron-lucent regions of granules, although AAA also labeled the electron-dense regions. Some LTA-positive Fuc residues were located in the electron-dense core of the secretory granules of the transitional cells. The β-elimination procedure abolished this staining, indicating that these residues were located in O-linked oligosaccharides. It should be noted that the Fuc residues could also be identified in the electron-dense core of the secretory granules of the mucous neck cells after PNGase-F. This can be explained because of the removal of the N-linked oligosaccharides by the enzyme, which can make the carbohydrates accessible to LTA. With this lectin, some Fuc residues belonging to O-linked oligosaccharides were located in the electron-lucent granules of the immature parietal cells. The staining observed with AAA and UEA-I in the surface, gastric pit, mucous neck, and transitional cells was not abolished when pretreatment with β-elimination was used, suggesting the presence of Fuc residues also in N-linked oligosaccharides, in the same locations. It should be noted that β-elimination or PNGase-F treatment either did not modify or slightly decreased the AAA and UEA-I staining. This could be due to the unmasking of not previously detected oligo-saccharides, produced when other chains are removed. These unmasked oligosaccharide chains could contain Fuc residues that are now accessible to the lectin. This phenomenon has been previously described with other tissues and lectins (Leis et al. 1997). Fuc residues contained in N-linked oligosaccharides were also localized in intracellular canaliculi and the tubulovesicular system of the parietal cells, and in the granules of the immature parietal cells.
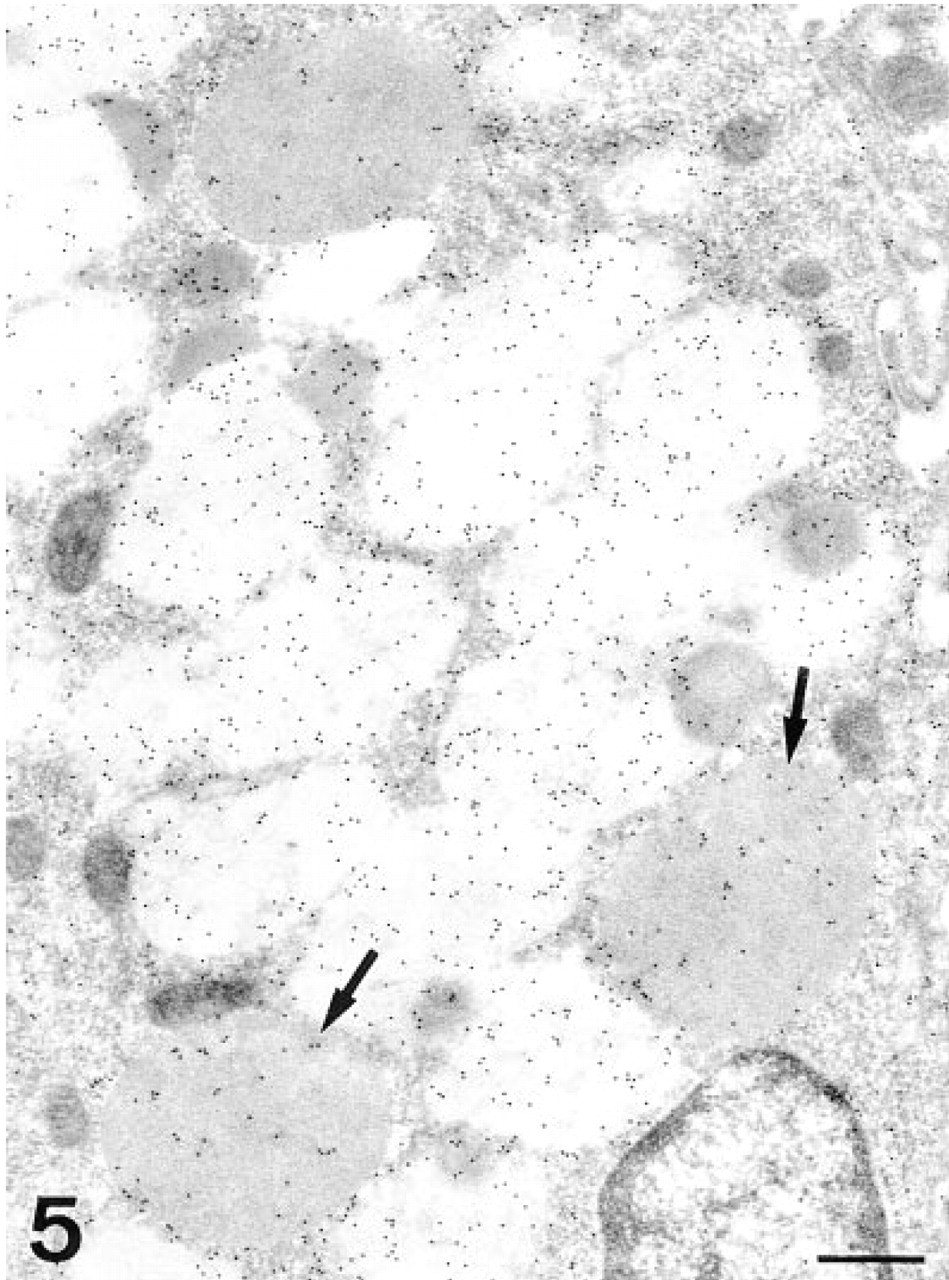
AAA, transitional cell. Although most of the labeling is observed on the electron-lucent regions of the secretory granules, some gold particles are also deposited in the electron-dense areas. Note that some granules are completely electron-dense (arrows). N, nucleus. Bar = 0.5 μm.
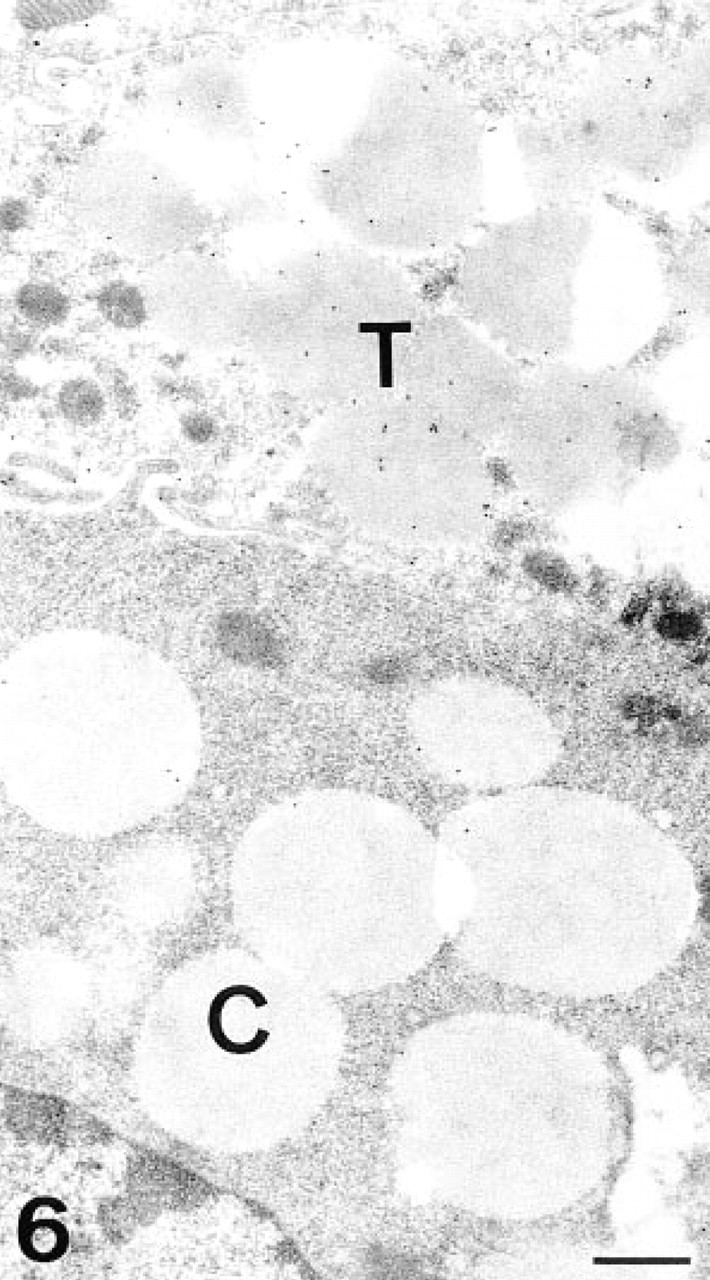
PNGase-F/AAA, transitional and chief cells. After the enzymatic treatment, the labeling in the transitional cell (T) has decreased, mainly in the electron-lucent regions. Note the absence of labeling in the chief cell (C). Bar = 0.5 μm.
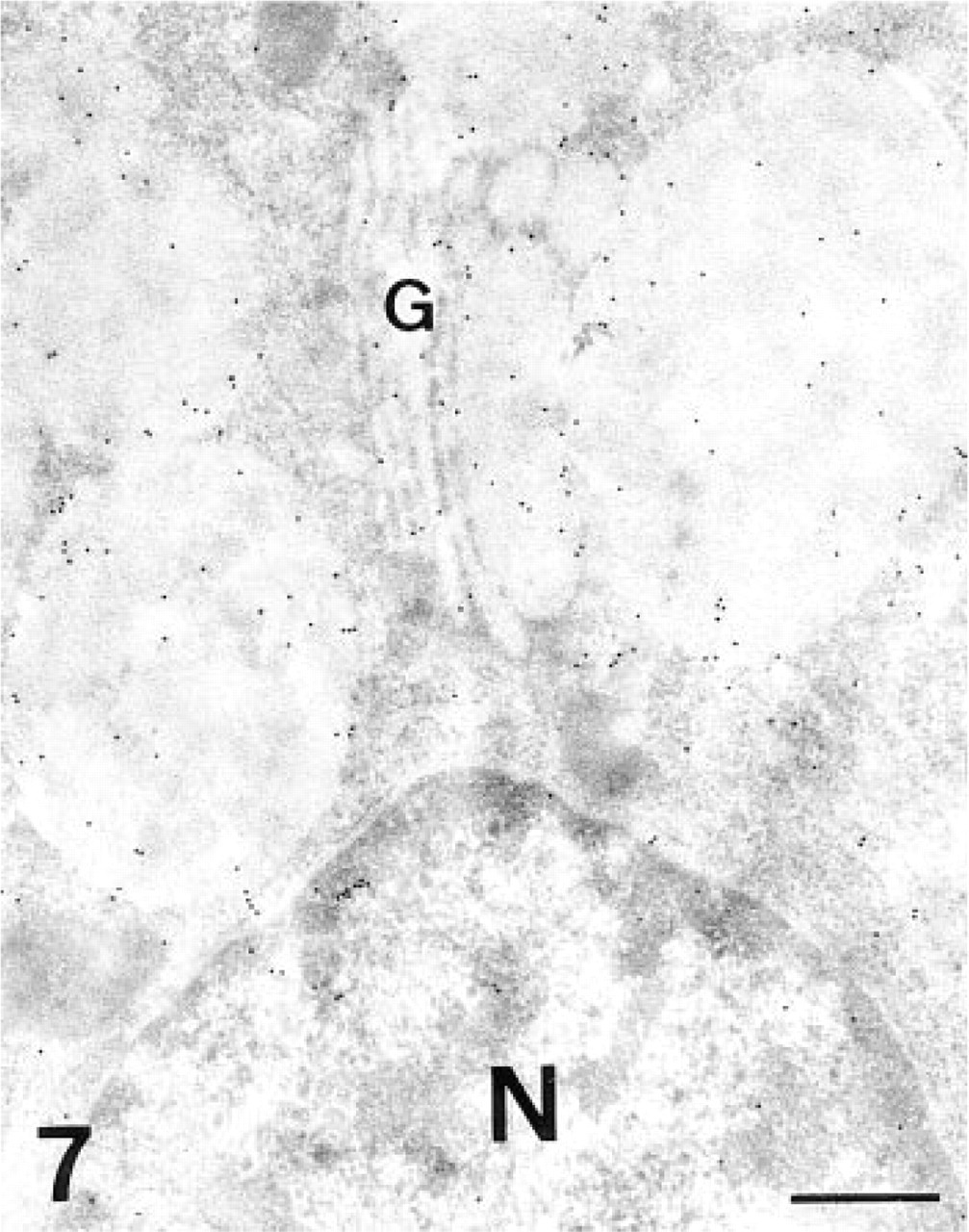
AAA, chief cell. Both the secretory granules and the Golgi stack (G) are labeled. N, nucleus. Bar = 0.5 μm.
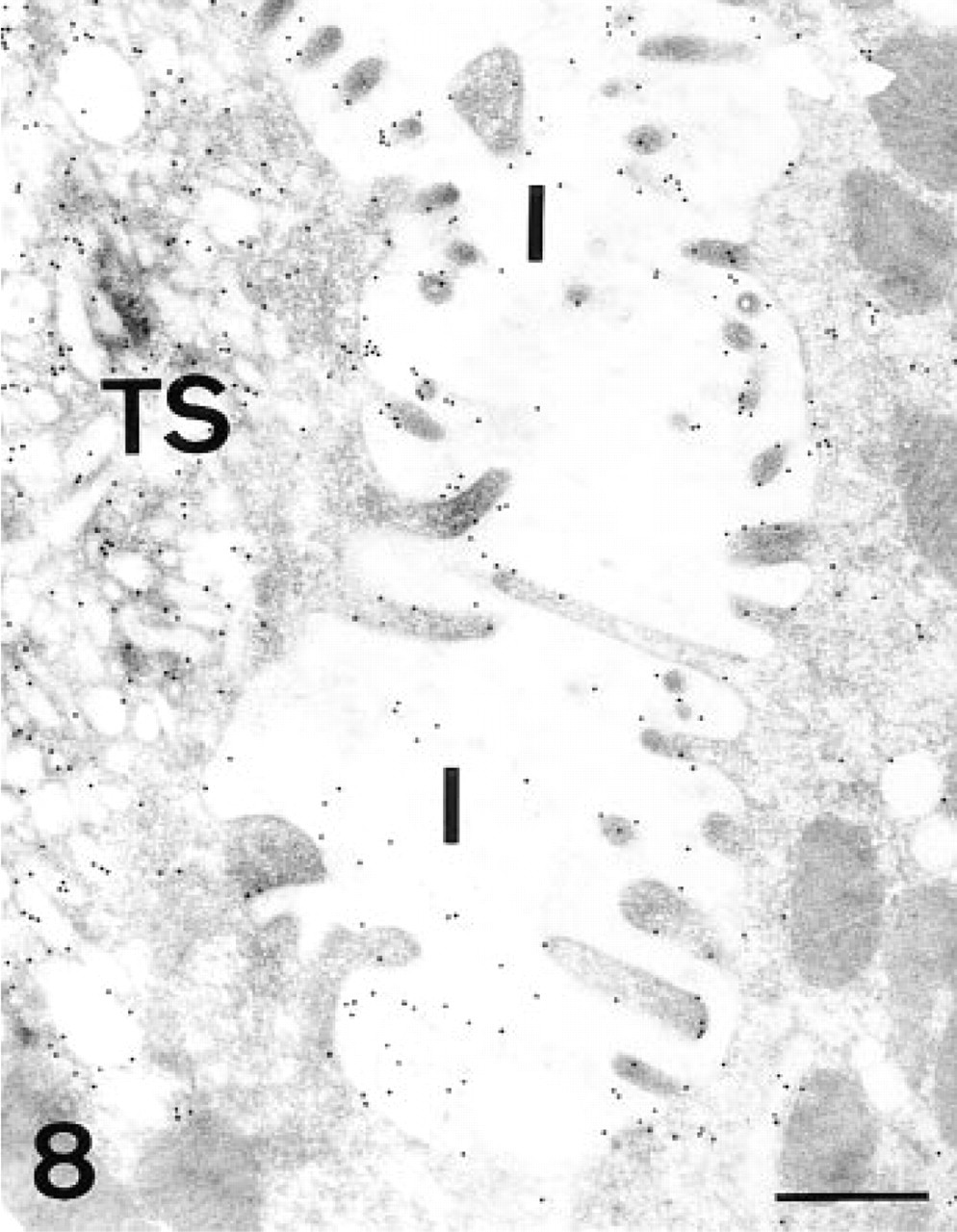
AAA, parietal cell. Gold granules are deposited on the membranes of both the intracellular canaliculi (I) and the tubulovesicular system (TS). Bar = 0.5 μm.
The most interesting finding with AAA was the labeling observed in the secretory granules of the chief cells. This labeling disappeared after PNGase-F treatment, indicating that the Fuc residues were contained in N-linked oligosaccharides. Neither UEA-I nor LTA labeled these carbohydrates. These two lectins preferentially label Fuc residues located in the outer region of the oligosaccharide chains (Pereira and Kabat 1974; Sugii and Kabat 1982), whereas AAA can also bind to those Fucα(1-6)-linked to the innermost GlcNAc residues of the core of N-linked oligosaccharides (Osawa and Tsuji 1987). Therefore, these findings suggest that the Fuc residues found in the secretory granules of the chief cells are linked to the core region of the N-linked oligosaccharides.
Lectin binding pattern in the human gastric mucosa a
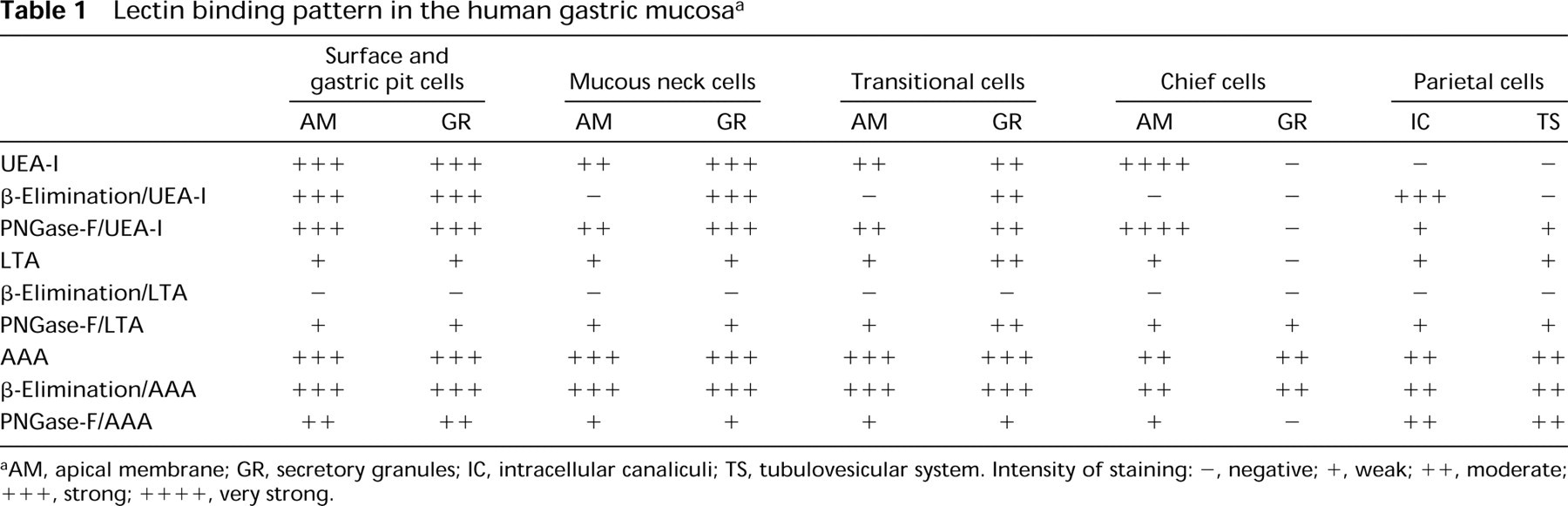
AM, apical membrane; GR, secretory granules; IC, intracellular canaliculi; TS, tubulovesicular system. Intensity of staining: -, negative; +, weak; ++, moderate; +++, strong; ++++, very strong.
The three lectins frequently labeled the trans face of Golgi apparatus or the small vesicle exiting from it. However, all the cisternae of the Golgi apparatus of the chief cells were labeled with AAA, and the labeling disappeared when PNGase-F was used. This indicates that these Fuc residues linked to the innermost GlcNAc of the N-linked oligosaccharides are added in the cis Golgi, whereas the Fuc residues located in the outer region of the N- and O-linked oligosaccharides identified in the other cell types are added in the trans Golgi. The addition of Fuc in the trans-Golgi face of the surface human gastric cells has been previously reported (Ríos-Martin et al. 1993).
For a long time, the presence of N-linked oligosaccharides on mucin glycoproteins has been doubted. Evidence for the presence of N-linked oligosaccharides in secretory mucous glycoproteins comes from biosynthetic studies (Strous and Dekker 1992; Klomp et al. 1994). About 10 N-linked oligosaccharides are cotranslationally attached to the backbone moiety of rat gastric mucin (Strous and Dekker 1992). Our results provide new evidence for the presence of N-linked oligosaccharides in the human gastric mucins secreted by the surface, gastric pit, and mucous neck cells. Moreover, glycoproteins containing N-linked oligosaccharides are also secreted by the chief cells.
The three lectins used also recognize the H-antigen (Ito and Hirota 1992). The use of lectins as markers of blood groups can be controversial. The lectin binding patterns of UEA-I, LTA, and AAA are not identical to that previously described for the anti-H antibody in the human fundic glands (Madrid et al. 1998b). All the structures labeled by the antibody were also positive for the lectins, but some additional structures were labeled only by the lectins. This indicates that the lectins recognize other different Fuc residues in our tissue. On the other hand, other blood group-specific lectins are good markers of the corresponding blood group, e.g., Griffonia simplicifolia agglutinin I-B4 (Ito et al. 1997).
The biological significance of the secretion of fucosylated oligosaccharides by the human gastric glands could be related to the cytoprotection role of the gastric mucous glycoproteins. H. pylori infection is related not only to gastric ulcers but also, probably, to gastric cancer (Goldstone et al. 1996). There can be little doubt that, in the gastrointestinal tract, mucous secretions play an important role in restricting the movement of bacteria to the surface epithelium. The bacteria became trapped in the mucus and are eventually excreted in feces (Wallace and Granger 1996). Recently, it has been demonstrated that the H. pylori adherence to the gastric epithelial lining is mediated by the fucosylated Lewis b histo-blood antigen (Ilver et al. 1998). The fucosylated oligosaccharides identified in our study are secreted to originate the mucous barrier and could prevent the migration of the bacteria to the surface epithelium.
At the electron microscopic level, two methods for UEA-I-labeling were used. The UEA-I-DIG method shows more intense labeling than UEA-I-gold. The first method includes 15-nm colloidal gold particles, whereas in the second the colloidal gold particles were larger (20 nm). The use of smaller colloidal gold particles usually increases the binding efficiency (Madrid et al. 1998a) but the particles are more difficult to visualize.
Implications of the Biphasic Nature of the Granules
The transitional cells have been considered as a stage in the maturation process from the mucous neck cells to the chief cells (Suzuki et al. 1983; Karam and Leblond 1993c; Madrid et al. 1990,1998b). During this maturation process, the electron-dense core of the secretory granules or mucous neck cells progressively increases in size as the cells descend through the gland. Thus, the transitional cells can contain secretory granules with a different electron-lucent/electron-dense core ratio. The last step would be the absence of electronlucent regions, originating the chief cells (Suzuki et al. 1983; Karam and Leblond 1993c; Madrid et al. 1990,1998b). Our study demonstrates that the composition of the two different compartments (electron-dense and electron-lucent) of the granules in the transitional and mucous neck cells is different but that, at the same time, they are similar compared with the corresponding one of the other cell type. The different composition of these electron-dense and electronlucent regions has been described for other molecules when the electron microscope has been used, e.g., different sugar residues, pepsinogen, osteopontin (Madrid et al. 1990; Waalewijn et al. 1991; Qu-Hong et al. 1997). A similar finding has been described for other tissues (Castells et al. 1992,1994; Leis et al. 1997) or for the fundic glands of other mammalian species (Suganuma et al. 1985; Yamada et al. 1988; Wasano and Hirakawa 1997). The mucous neck cells of the rat stomach have been reported to contain similar biphasic granules, the electron-lucent regions containing oligosaccharides and the electron-dense core being liable to digestion with pronase (Yeomans 1974; Suganuma et al. 1985). Both the epithelial and mucous neck cells, but not the chief cells, have been reported to contain the same mucin peptide backbone (Toribara et al. 1993), probably in the electron-lucent regions of the granules.
Our present results and the previous data indicate that the composition of the electron-dense regions is similar to that of the secretory granules of the chief cells in human and other mammalian fundic glands (Suganuma et al. 1985; Yamada et al. 1988; Qu-Hong et al. 1997; Wasano and Hirakawa 1997; Madrid et al. 1990,1998b). The cells proliferate in the neck region of the fundic glands and the subsequent cells migrate outward to the epithelia and gastric pits and inward to the fundic glands (Karam and Leblond 1993a, b; Yang et al. 1997). During this migration the cells differentiate into mucous neck cells and then into transitional and chief cells. This is not merely a morphological change; the cells have to gradually change their secretory mechanism. First, they mainly synthesize the glycoproteins of the electron-lucent regions and, gradually, the synthesis of these glycoproteins is substituted by that of the secretory granules of the chief cells.
Concluding Remarks
In summary, in the present study the presence of fucosylated oligosaccharides in the surface and fundic glands cells of the human stomach has been demonstrated. These oligosaccharides are present in the secretory granules of the surface, gastric pit, mucous neck, and transitional cells, and they are N- or O-linked to the protein core. Moreover, Fuc residues linked to the innermost GlcNAc of the core of N-linked oligosaccharides were localized with AAA in the chief cells and in the electron-dense regions of the secretory granules in the transitional cells. The findings suggest that the maturation process from the mucous neck cells to the chief cells requires a complicated change in the biosynthetic machinery of the cell, and that the cells clearly recognize which glycoproteins have to be packed in the electron-dense or electron-lucent regions of the secretory granules.
Footnotes
Acknowledgments
Supported by grants PB 93-1123-C02-02 from Ministerio de Educatión y Cultura and UPV 075.327-EC236/95 from the University of the Basque Country. OL is supported by a fellowship from the Ministerio de Educación y Cultura (Spain).
We are greatly indebted to Ms C. Otamendi, Ms M.C. González, and Mr J.A. Madrid for excellent technical assistance.