
Abstract
Wound healing conditions generate a stressful environment for the cells involved in the regeneration process and are therefore postulated to influence the expression of heat shock proteins (Hsps). We have examined the expression of four Hsps (Hsp27, Hsp60, Hsp70 and Hsp90) and a keratin (keratin 6) by immunohistochemistry during cutaneous wound repair from Day 1 to Day 21 after wounding in the mouse. Hsps were constitutively expressed in normal mouse epidermis and their patterns of expression were modified during the healing process. The changes were not directly linked to the time course of the healing process but rather were dependent on the location of cells in the regenerating epidermis. In the thickened epidermis, Hsp60 was induced in basal and low suprabasal cells, Hsp70 showed a reduced expression, and Hsp90 and Hsp27 preserved a suprabasal pattern with an induction in basal and low suprabasal cells. All Hsps had a uniform pattern of expression in the migrating epithelial tongue. These observations suggest that the expression of Hsps in the neoepidermis is related to the proliferation, the migration, and the differentiation states of keratinocytes within the wound.
Keywords
T
During wound healing, cells are exposed to a variety of stresses originating from the wound environment. Cells within and surrounding the wound are subjected to dehydration and exposure to oxygen because the epithelial barrier is broken down (Hunt and Hussain 1992). The blood clot not only dehydrates but also stops the blood supply to the wound area. Consequently, nutrients are reduced, especially in a context of increased cell density (Ordman and Gillman 1966; Tarin and Croft 1970; Clark 1993). Therefore, the tissue glucose level falls (down to 1-2 mM), the partial pressure of oxygen (pO2) is reduced (10torr), and that of carbon dioxide (pCO2) is increased (80 torr) (Hunt and Hussain 1992). In the face of this metabolic crisis, cells use anaerobic glycolysis and produce lactic acid, which lowers the pH to 6.8 (Hunt and Hussain 1992). Reactive oxygen metabolites (O2 -, H2O2, HOCl, · OH, and NO) and hydrolytic enzymes, representing respectively oxidative and enzymatic stresses for the cells, are secreted by neutrophils and macrophages during the inflammatory phase (Elstner 1991; Wahl and Wahl 1992). Several inflammation mediators, cytokines and growth factors, e.g., prostaglandins, leukotrienes, PDGF, FGF, EGF, TGFβ, IGF, CSF, IFN, TNF, and IL, are found in wound fluids (Bennett and Schultz 1993; Kiritsy and Lynch 1993; Moulin 1995), and most of these stimulate proliferation and protein synthesis representing other stimuli that are susceptible to induce stress response mechanisms.
Cells defend themselves from stresses by expressing heat shock proteins (Hsps). These proteins, first reported to be induced by heat shock (Ritossa 1962), are believed to represent a mechanism of response to cellular stresses that protects intracellular proteins from damaging events. Several inducers of Hsps have been described, e.g., alcohol, UV radiation, oxidants, cell cycle, growth factors, infection, inflammation, recovery from anoxia and glucose starvation (Schlesinger 1990; Charveron et al. 1995; Maytin 1995; Minowada and Welch 1995b). Mammalian Hsps are classified into five main families on the basis of their molecular weights, Hsp27, Hsp60, Hsp70, Hsp90, and Hsp110, each having a variety of possible functions as reviewed in the literature (Lindquist and Craig 1988; Welch 1992; Hendrick and Hartl 1993; Craig et al. 1994). A chaperone function is generally admitted for many Hsps, as shown by their binding to several proteins and their involvement in conformational changes of proteins during folding, activation/inactivation, disaggregation, renaturation, and intracellular transport (Ciocca et al. 1993; Hendrick and Hartl 1993; Kampinga 1993). This chaperone function is based on their interaction with hydrophobic segments on proteins and could explain some of the cell defense mechanisms provided by Hsps.
Considering that several stressful cellular conditions are present in the wound environment, we examined the expression of Hsps during wound healing of mouse skin. Hsp60, Hsp90, Hsp27, and Hsp70 were detected by immunofluorescence during inflammation and organization of the inflammatory exudate that leads to granulation tissue formation and reepithelialization. This report focuses on the epidermal compartment.
Materials and Methods
Surgery
Six-week-old CD-1 mice (Charles River Laboratories of Canada; St-Constant, PQ, Canada) were used in these studies. Anesthesia was produced with 0.9 ml of ketamine (20 mg/ml) and xylazine (2.5 mg/ml) injected into the leg. One full-thickness skin wound (4 cm2 surface area) was made on the back. Wounds were left to heal by second intention without any device. Mice were sacrificed by overdose of anesthetics and biopsied at different times from 24 hr to 21 days after wounding (two mice per time of biopsy). Wound healing tissue and normal skin were embedded in OCT and frozen sections (4 μm thick) were prepared. This study was repeated twice. The Ethics Committee of Laval University approved animal experiments in accordance with the guidelines of the Canadian Council on Animal Care.
Antibodies
The primary antibodies used for the immunodetection of the different Hsps and keratin 6 were all polyclonal rabbit antibodies. The anti-human keratin 6 antibody was a gift of Dr. Pierre A. Coulombe (Paladini et al. 1996). The anti-human Hsp60 antibody (#37) is directed against the entire protein. Briefly, a human Hsp60 cDNA clone (gift of Dr. L. Weber; Reno, Nevada) was inserted in a pET vector and the expressed fusion protein used as immunogen as described previously (Tanguay et al. 1993). The anti-human Hsp90 antibody (#801) is directed against the 165-amino-acid C-terminal domain of the β-chain (Tanguay et al. 1993). The anti-rodent Hsp27 antibody (L2R3) is directed against the C-terminal part specific for rodents (AGKSEQSGAK) (Lavoie et al. 1995). The anti-human Hsp70 antibody (#799) is directed against the 216-amino-acid C-terminal domain and recognizes specifically the inducible form of the protein (Tanguay et al. 1993; Biggiogera et al. 1996). The secondary antibody used for immunoblotting on nitrocellulose membranes was a horseradish peroxidase-conjugated goat anti-rabbit IgG antibody (Sigma-Aldrich Canada; Oakville, Ontario, Canada). Rhodamine-conjugated goat anti-rabbit IgG antibody (Canadian Chemicon; Missisauga, Ontario, Canada) was used as a secondary antibody for indirect immunofluorescence.
Gel Electrophoresis and Immunoblot Analysis
The specificity of the primary anti-Hsp antibodies was demonstrated using total proteins from normal mouse skin and Western blot analysis as described previously (Tanguay et al. 1993; Michel et al. 1996). Skin tissue was homogenized on ice in Tris buffer (30 mM, pH 6.8) with sodium azide (0.5 mg/ml) and protease inhibitors: ethylenedinitrilo disodium tetraacetate (0.93 mg/ml), N-ethylmaleimide (1.25 mg/ml), benzamidine (0.78 mg/ml), and phenylmethylsulfonyl fluoride (0.17 mg/ml). The homogenized tissue was dissolved in sodium dodecyl sulfate (3%) and glycerol (126 mg/ml) in a boiling water bath for 10 min. Total proteins (60 μg/well) were separated by one-dimensional gel electrophoresis in a 5% acrylamide stacking gel (pH 6.8) and a 10% running gel (pH 8.8) according to Laemmli (1970). Proteins were fixed on gels and visualized by Coomassie blue staining. Proteins from unfixed gels were electroblotted onto nitrocellulose membranes (Amersham Canada; Oakville, Ontario, Canada) according to the method of Towbin et al. (1979). The membranes were stained with Ponceau red, rinsed, and blocked with skim milk (5%). Immunodetection of Hsp60, Hsp90, Hsp27, and Hsp70 was performed with the polyclonal primary antibodies (1/1000), horseradish peroxidase-conjugated secondary antibody (1:5000), and the ECL chemiluminescence detection kit (Amersham Canada) according to the manufacturer's recommendations.
Immunofluorescence Microscopy
Indirect immunofluorescence labeling of frozen sections (fixed for 10 min in −20C methanol) was realized using polyclonal anti-keratin 6 (1:500), Hsp60 (1:100), Hsp90 (1:100), Hsp27 (1:100), and Hsp70 (1:100) antibodies and rhodamine-conjugated secondary antibody (1:100) as described previously (Moulin et al. 1997). In controls, normal rabbit serum was used at the same dilution as the primary antibody. Cell nuclei were labeled with 0.5 μg/ml Hoechst 33258 (Sigma-Aldrich Canada). The immunofluorescence microscopic studies were performed with a Nikon UFX-IIA microscope and all photographs were taken at the same exposure time determined for each magnification power.
Results
Antibody Specificities
The specificity of the primary antibodies against Hsp27, Hsp60, Hsp70, and Hsp90 was demonstrated by Western blot analysis of total proteins from normal mouse skin and is shown in Figure 1.
Keratin 6
Keratinocytes of normal skin did not express keratin 6, except in the hair follicles, but this keratin was present in the regenerating epidermis. Figure 2 shows a low magnification of normal skin adjacent to the regenerating epidermis (48 hr after wounding) labeled with an anti-keratin 6 antibody. The different locations with regard to the wound margin that will be stated in this report are identified. Because this photograph was taken 48 hr after wounding, the thickened epidermis (TE) above the granulation tissue was short compared to that present above the dermis (at the wound margin). The length of the TE over the granulation tissue increased as a function of time after wounding. Keratinocytes of normal epidermis were not labeled, whereas keratin 6 was expressed by low suprabasal cells in the TE (see Figure 2 inset). Behind the migrating epithelial tongue (MET), a suprabasal pattern of keratin 6 was preserved. In the MET, uniform labeling of all cell layers was observed. Therefore, the pattern of expression of keratin 6 varied within the epithelium regenerating over the granulation tissue, which constitutes the neoepidermis.
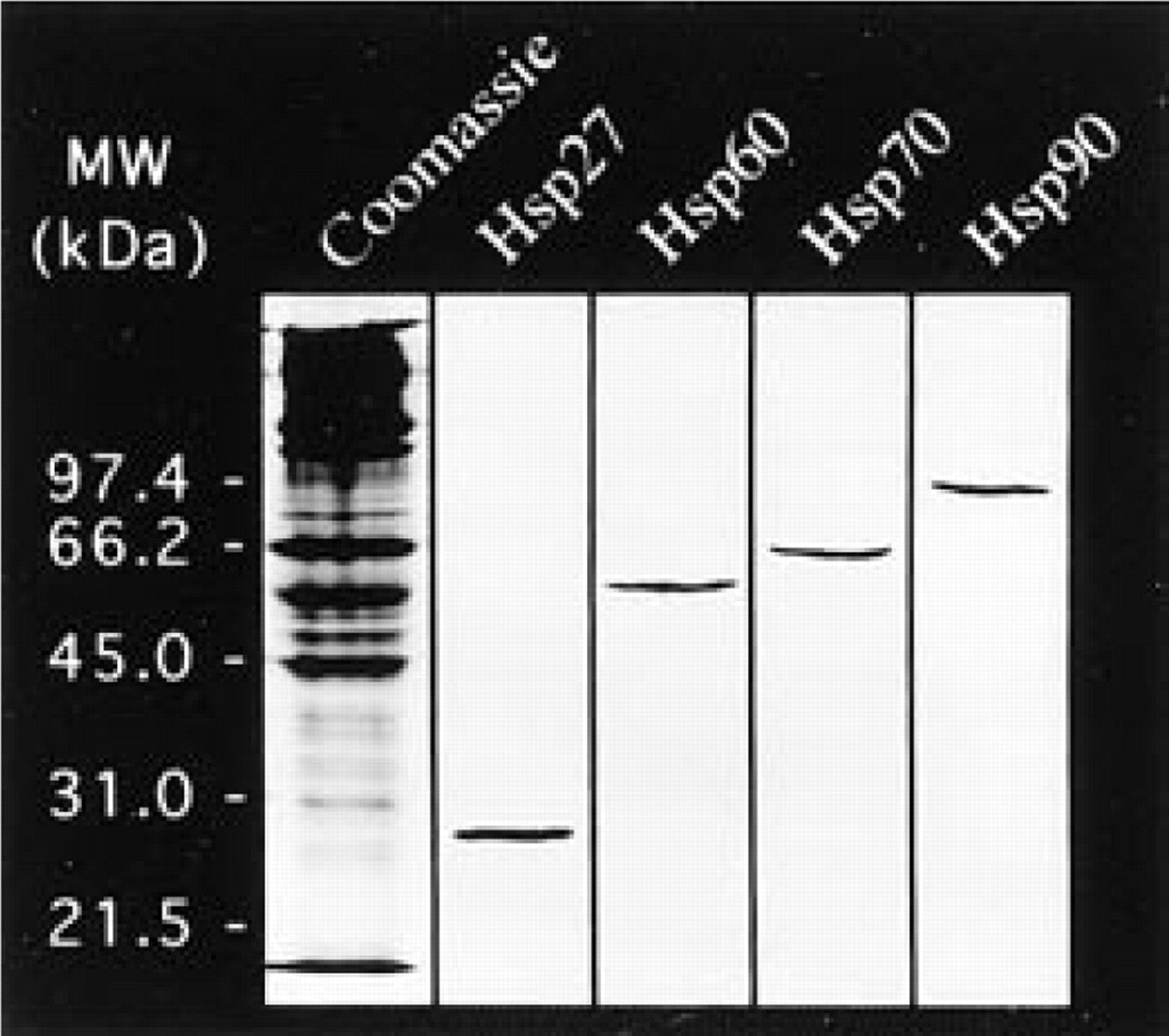
Western blot analysis of normal mouse skin (60 μg of total proteins/well). The specificity of the anti-Hsp27, Hsp60, Hsp70, and Hsp90 primary antibodies is demonstrated. Total proteins stained with Coomassie blue are shown in the left lane and the location of molecular weight standards is indicated.
Variations in Expression of Hsps as a Function of the Location of Keratinocytes in the Regenerating Epidermis
Each Hsp showed a specific pattern of expression in normal mouse epidermis, and different modifications of these patterns were observed in wound healing tissue. In the regenerating epidermis, the variations were dependent on the location of keratinocytes with regard to the wound margins rather than on the time after wounding. Therefore, our observations are presented in terms of location within the wound (TE, behind MET, MET, regenerating epidermis, and neoepidermis; see Figure 2).
Hsp60
In normal mouse epidermis, Hsp60 was present at a low level with a uniform distribution (Figure 3A). In the regenerating epidermis, the pattern of expression differed from that observed in normal skin. In the TE (Figures 3D and 3E), many granules were observed in the basal and low suprabasal keratinocytes, whereas the high suprabasal cells were only lightly labeled. The location of this deep granular labeling varied according to the length of the regenerating epidermis; the longer it was, the further towards the center of the wound the labeling was. The neoepidermis behind the MET had this deep granular labeling at its maximum of intensity. In the MET, fewer granules were observed (Figure 3C) and these were uniformly spread, with an intermediate intensity between the labeling of normal epidermis (Figure 3A) and that of adjacent neoepidermis (Figures 3D and 3E). Controls (for the four Hsps studied) of normal and regenerating mouse epidermis (with normal rabbit serum) showed minor background (Figures 3B and 3F).
Hsp90
Normal mouse epidermis had a high suprabasal pattern of expression, basal and low suprabasal cells having very light labeling and nuclei being negative (Figure 4A). During healing, Hsp90 preserved a suprabasal pattern in the TE (Figures 4B and 4C) and the intensity was comparable to that observed in normal epidermis (Figure 4A). The labeling was located in the very high suprabasal cells and the stratum corneum progressively lost this labeling as the cells were reaching the surface. Basal and low suprabasal keratinocytes were slightly more labeled in the thickened neoepidermis (Figures 4B and 4C) than in the normal epidermis (Figure 4A) or in the TE above dermis (Figure 4B). Behind the MET (Figure 4D), the neoepidermis preserved the same suprabasal pattern of expression, with a reduction in the intensity of both basal and suprabasal labeling. The MET showed faint and uniform labeling (Figure 4E).
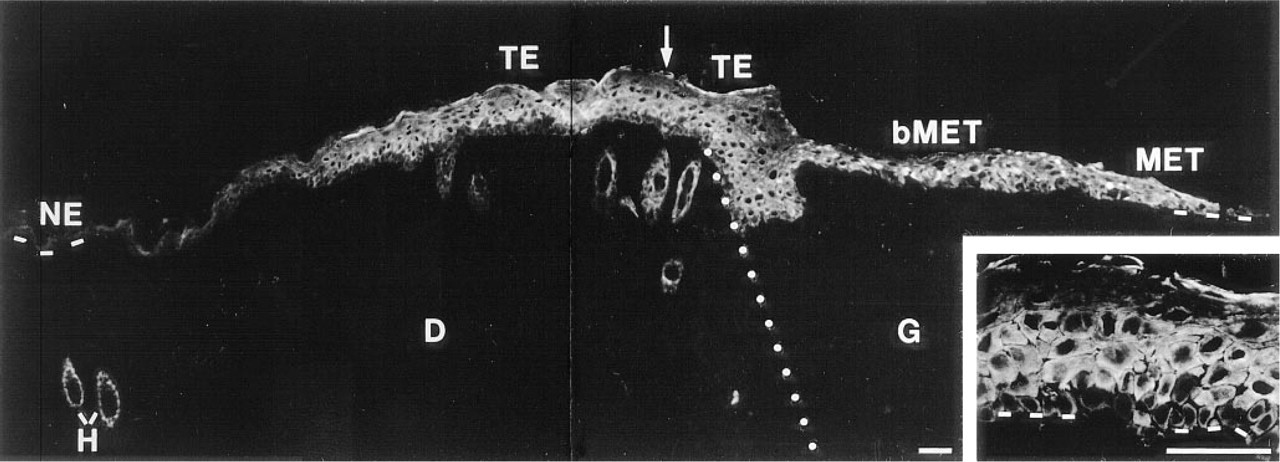
Indirect immunofluorescence study of keratin 6 expression in normal and healing mouse skin. Normal skin is at left and wound healing tissue is at right; dotted line represents the location of the wound margin (as in other figures). No labeling is present in normal epidermis (NE). In the TE, the labeling is located mainly in the low suprabasal keratinocytes, whereas basal and high suprabasal cells are less labeled (see
Hsp27
Normal mouse epidermis had a suprabasal pattern of expression, but the stratum corneum was not labeled (Figure 5A). The labeling was mainly cytoplasmic, with nuclei showing only faint staining. Basal and low suprabasal keratinocytes showed a slight expression of Hsp27. In the wound, the TE above the dermis (at the wound margin) had a pattern and level of expression (Figures 5C and 5D) similar to those of normal epidermis (Figure 5A). In the TE over the granulation tissue (in the wound), the labeling intensity of superficial cells was reduced whereas that of deep cells was lightly increased (Figures 5C and 5D). Instead of being concentrated in a thin suprabasal band, as in the normal epidermis (Figure 5A), the labeling was spread over a wider band (Figures 5B–5D). In some areas, basal keratinocytes showed a higher labeling (Figure 5D), which was more abundant in the TE over dermis (at the wound margin) and decreased towards the MET. Behind the MET, the neoepidermis was characterized by a granular distribution of Hsp27 in the suprabasal keratinocytes (Figure 5B). From the edge of the wound, these granules increased in number and intensity, producing a higher level of labeling. The MET was characterized by a uniform pattern and the intensity of the labeling was comparable to that of adjacent basal or low suprabasal cells (Figure 5E). The expression of Hsp27 by suprabasal keratinocytes in normal and TE (Figures 5A–5D) was seen in lower epidermal layers than that observed for Hsp90 (Figures 4A–4D).
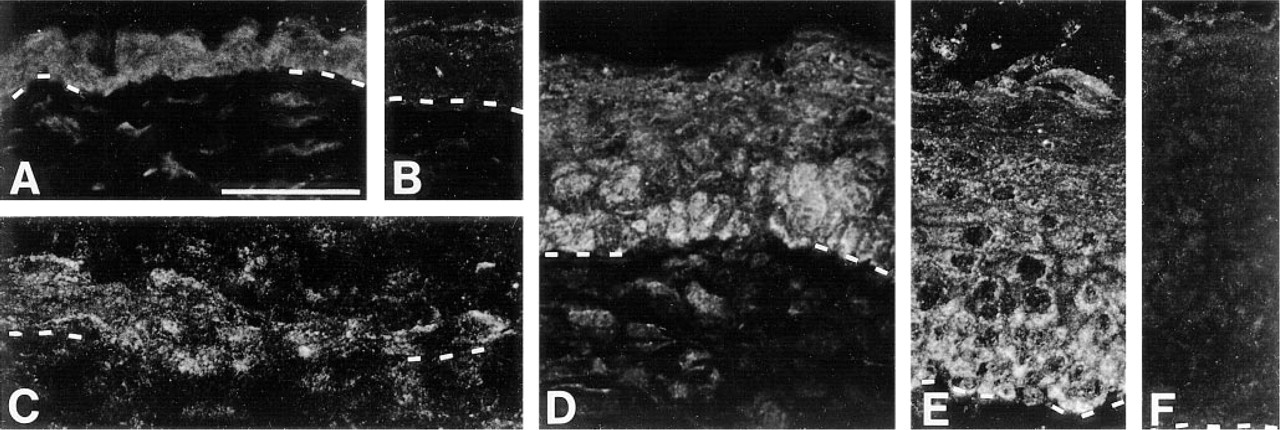
Indirect immunofluorescence study of Hsp60 expression in normal and regenerating mouse epidermis.
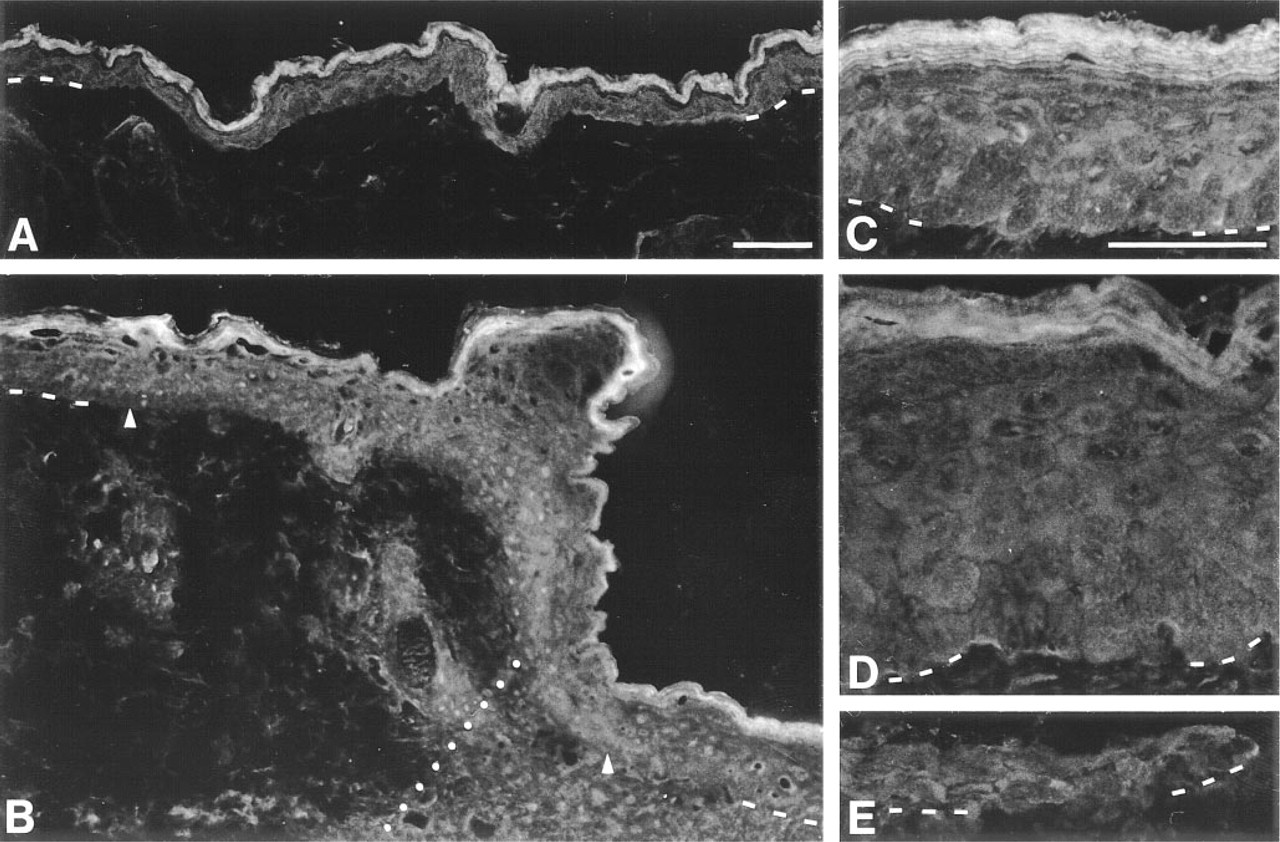
Indirect immunofluorescence study of Hsp90 expression in normal and regenerating mouse epidermis.
Inducible Hsp70
In normal mouse epidermis, the Hsp70 labeling was diffuse rather than granular and was slightly more visible in suprabasal keratinocytes than in basal ones (Figure 6A). The stratum corneum was not labeled. Some nuclei were lightly stained with a granular pattern. In the wound, the neoepidermis (Figures 6B–6E) had a different pattern of expression and a reduced labeling intensity compared to normal epidermis (Figure 6A) or to adjacent TE above dermis (Figure 6D). The thickened neoepidermis showed reduced and uniform Hsp70 labeling (Figure 6D). Many positive nuclei could be observed (Figures 6C and 6D). Distal to the wound margin, basal and low suprabasal keratino-cytes were more labeled than high suprabasal cells, producing a gradient of labeling (Figures 6B and 6C). The pattern of expression was therefore reversed in comparison with normal epidermis (Figures 6B and 6C vs Figure 6A). Fewer positive nuclei were found behind the MET (Figure 6B), and almost none were observed in the MET (Figure 6E). Keratinocytes in the MET had a lower expression of Hsp70 and the gradient of labeling from basal to suprabasal cells was lost (Figure 6E vs Figures 6B and 6C).
Table 1 summarizes the results observed on the expression of Hsp60, Hsp90, Hsp27, and Hsp70 in the normal vs the regenerating epidermis.
Discussion
Indirect immunofluorescence on frozen sections was used to follow the expression of keratin 6 and four Hsps during reepithelialization of mouse skin after wounding. Each Hsp showed a specific pattern of expression in normal epidermis and these patterns were modified in regenerating epidermis, as summarized in Table 1.
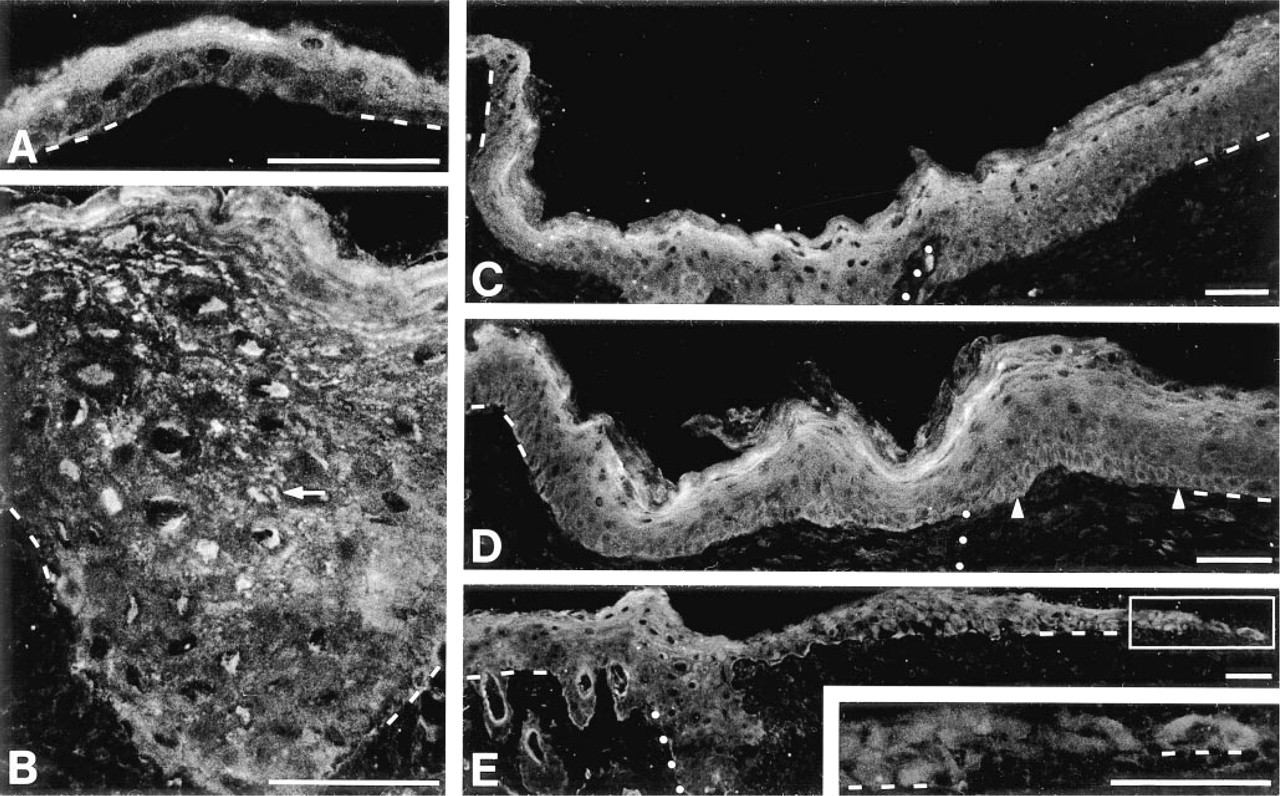
Indirect immunofluorescence study of Hsp27 expression in normal and regenerating mouse epidermis.
In normal skin, the basic keratin 6 is associated with the acidic keratin 16 and both are expressed only by cells in the outer root sheet of hair follicles (Moll et al. 1982; Stark et al. 1987). In the regenerating epidermis, keratin 6 was expressed by keratinocytes in the low suprabasal layers, as already reported by others (Mansbridge and Knapp 1987; Schermer et al. 1989; Paladini et al. 1996). The expression of keratin 6 and 16 was associated with epidermal hyperproliferation (Weiss et al. 1984; Tyner and Fuchs 1986), but recent studies rather associate these keratins with an activated state of keratinocytes (Schermer et al. 1989; Jiang et al. 1993; Paladini et al. 1996). The latter concept correlates better with the observed absence of labeling in basal keratinocytes of the thickened regenerating epidermis and with the expression of keratin 6 in the MET, where cells are known to be migrating rather than proliferating (DiPasquale 1975; Pang et al. 1978; Ortonne et al. 1981; Stenn and Malhotra 1992).
Each Hsp was expressed constitutively with a specific pattern in normal epidermis. However, the patterns and levels of expression were modified during wound healing. Unexpectedly, the changes observed in the regenerating epidermis were not linked to the time after wounding but rather were dependent on the location of keratinocytes in the regenerating epidermis. Each Hsp was expressed by keratinocytes of specific layers in normal and regenerating epidermis. Hsp60 was located in the basal and low suprabasal cells. Hsp90 and Hsp27 were mostly found in the high suprabasal cells, Hsp90 being expressed higher in the differentiation process than Hsp27. Hsp70 could be found throughout the epidermal compartment. Although the pattern of expression of each Hsp was affected differently after wounding, the MET always had a uniform expression of Hsps, the labeling being similar for the basal and suprabasal cells.
Keratinocytes located in the regenerating epidermis expressed more Hsp60 than those in normal epidermis. The pattern of expression varied from a light and uniform labeling in normal epidermis to a granular pattern located mainly in the basal and low suprabasal cells of the TE and finally to a uniformly distributed granular pattern in the MET. This granular pattern can be explained by the mitochondrial origin of Hsp60 (Hendrick and Hartl 1993; Horwich and Willison 1993; Craig et al. 1994). It could easily be observed because mitochondria have been shown to be larger and longer in the TE (Odland and Ross 1968). In the MET, the granular labeling was slightly reduced compared to the neoepidermis behind; this result is in agreement with previous observations that show quite normal mitochondria in cells of the MET except for the presence of dense bodies (Croft and Tarin 1970). Keratinocytes are known to be migrating in the MET (Di-Pasquale 1975; Pang et al. 1978; Ortonne et al. 1981; Stenn and Malhotra 1992) and proliferating in deep epidermal layers of the TE (Stenn and Malhotra 1992; Kirsner and Eaglstein 1993). Therefore, the presence of Hsp60 may be associated with folding, activation/inactivation, or conformational changes of mitochondrial proteins that would be required during migration or proliferation (Langer and Neupert 1991; Gething and Sambrook 1992; Hendrick and Hartl 1993; Horwich and Willison 1993). Oral lichen planus (Bramanti et al. 1995) and cholesteatoma (Shinoda and Huang 1996) are two pathologies characterized by the hyperproliferative state of keratinocytes that have also been shown to express more Hsp60 in basal and low suprabasal cells. Therefore, the increased expression of Hsp60 is likely associated with the status of proliferation and migration of keratinocytes that are induced by the wound environment.
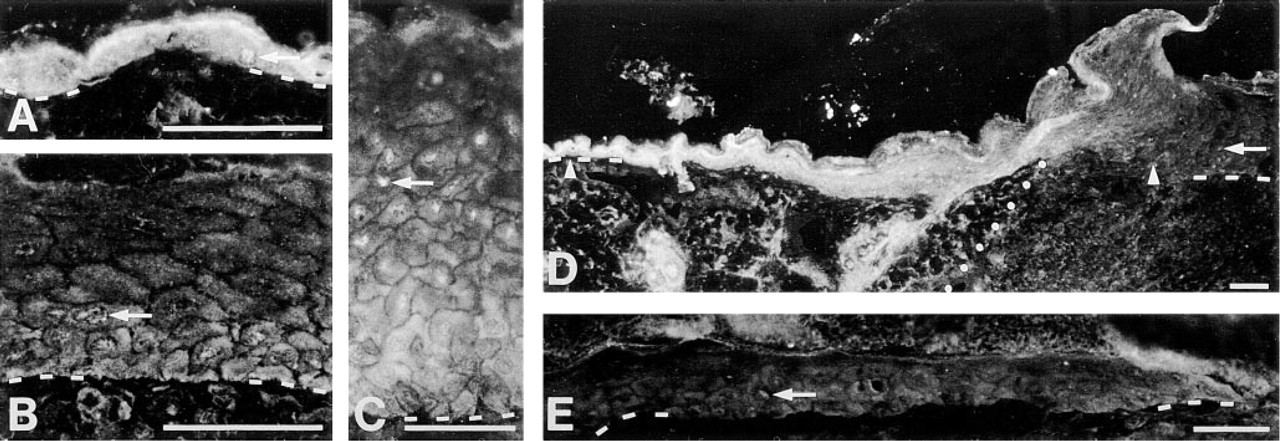
Indirect immunofluorescence study of Hsp70 expression in normal and regenerating mouse epidermis. Arrows indicate positive nuclei.
Hsp60, Hsp90, Hsp27, and Hsp70 in normal and regenerating epidermis

+(+) signifies that the labeling varies from + to ++ and ++ (+) from ++ to +++.
Hsp90 had a very high suprabasal pattern of expression in normal and regenerating epidermis (except in the MET). Hsp27 also had a suprabasal pattern of expression in normal epidermis, as already shown in human skin (Gandour Edwards et al. 1994; Trautinger et al. 1995), and this suprabasal pattern was preserved in the regenerating epidermis (except in the MET). These suprabasal patterns of expression for Hsp90 and Hsp27 can be associated with cell differentiation as in the case of other markers such as keratins, filaggrin, involucrin and loricrin (Holbrook and Wolff 1987; Mansbridge and Knapp 1987; Stenn and Malhotra 1992; Fuchs 1993).
In the TE, the labeling pattern of Hsp90 was similar to that of normal epidermis, with maximal expression in the very high suprabasal cells. Although cultured human keratinocytes have been shown to respond to physical and chemical stresses by the induction of a 90-kD protein (Maytin et al. 1990; Edwards et al. 1991; Holland et al. 1993), we observed that the stress of the wound healing conditions induced only a slight expression in basal and low suprabasal cells of the TE. In agreement with our results, no major difference is found in the level of expression of Hsp90 between oral lichen planus and normal mucosa (Bramanti et al. 1995). The reduction in the expression of Hsp90 towards the MET could be explained by the absence of terminally differentiated keratinocytes as shown by alterations in the ultrastructure, expression of keratins, and other differentiation markers (Croft and Tarin 1970; Ortonne et al. 1981; Mansbridge and Knapp 1987; Stenn and Malhotra 1992; Paladini et al. 1996). Moreover, this lowered expression of Hsp90 is consistent with prior observations that show a reduced expression of Hsp90 in vitro under glucose, calcium, or serum (growth factors) deprivation conditions (Jerome et al. 1991; Charveron et al. 1995). A low glucose level has been reported in wound fluids (Hunt and Hussain 1992). The functional significance of Hsp90 in wound healing keratinocytes remains unknown, but the literature is abundant concerning the interaction of Hsp90 with specific proteins, including steroid hormone receptors (Pratt 1993; Hutchison et al. 1994; Maytin 1995), tyrosine kinases (Welch 1992; Craig et al. 1994), and specific DNA sites (Morcillo et al. 1993; Biggiogera et al. 1996).
Hsp27 was mainly expressed by suprabasal keratinocytes in normal and regenerating epidermis (except in the MET). This expression of Hsp27 mainly by keratinocytes in a low proliferative and highly differentiated state is consistent with the data published by others on different cell types and tumors (Ciocca et al. 1993; Minowada and Welch 1995a; Trautinger et al. 1995). Interestingly, basal keratinocytes of the TE, which are in a less differentiated and highly proliferative state (Stenn and Malhotra 1992; Kirsner and Eaglstein 1993), showed some areas of Hsp27 expression. Similar areas of basal labeling were reported in some skin lesions such as psoriasis, dermatitis, and seborrheic keratosis (Gandour Edwards et al. 1994; Trautinger et al. 1995). Mainly in keratinocytes behind the MET, Hsp27 had a granular distribution, as already observed by others in cultured keratinocytes after heat shock or arsenite treatments, as well as in other cell types under various conditions (Ciocca et al. 1993; McClaren and Isseroff 1994). The granular distribution of Hsp27 was different from that seen for Hsp60, the latter being a mitochondrial protein. In the MET, a light and uniform pattern of expression was observed that could be explained by the absence of terminally differentiated keratinocytes, as discussed for Hsp90. One of the well-documented biochemical activities of Hsp27 is to regulate F-actin polymerization (Benndorf et al. 1994; Landry and Huot 1995; Lavoie et al. 1993,1995; Huot et al. 1997). This activity is regulated by phosphorylation of the protein in a stress-activated signaling pathway involving the MAPK SAPK2/p38 and the MAPKAP kinase-2/3 (Rouse et al. 1994; Landry and Huot 1995; McLaughlin et al. 1996; Guay et al. 1997; Huot et al. 1995,1997). In endothelial cells, activation of the SAPK2/p38 pathway appears to be an essential mediator of the activation of cell migration in response to VEGF (Rousseau et al. 1997). A high concentration of Hsp27 has also been shown to stabilize stress fibers during stress (Huot et al. 1995,1997). A regulated expression of Hsp27 in the regenerating tissue may thus serve both a regulatory function in mediating keratinocyte migration and a protective function against the wound-stressful environment.
In normal epidermis, keratinocytes constitutively express the inducible form of Hsp70, as shown by others (Trautinger et al. 1993; Boehncke et al. 1994; Charveron et al. 1995). A light suprabasal pattern and a distribution in both the cytoplasm and the nuclei of keratinocytes were observed. In comparison, the neoepidermis showed reduced and uniform expression of Hsp70, followed by an induction in basal and low suprabasal cells distal to the wound margin. Other stressful conditions have also been reported to induce the expression of Hsp70 in human keratinocytes in vitro and in situ such as heat, cold, chemicals, and UV radiation (Edwards et al. 1991; Muramatsu et al. 1992; Holland et al. 1993; Maytin 1995). The low expression of Hsp70 by keratinocytes of the neoepidermis and the basal-low suprabasal pattern behind the MET are not unique to wound healing. Similar observations have been made in oral lichen planus (Bramanti et al. 1995) and focally in psoriasis (Boehncke et al. 1994). Over the 21-day period of our study, the induction of Hsp70 by keratinocytes of the deep layers behind the MET and the low expression by keratinocytes of the MET are consistent with previous in vitro results up to 8 hr about wounding of monkey renal epithelial cells, in which the expression of Hsp70 is induced only at the wound edge and not away from the edge (Pawar et al. 1995). In the regenerating epidermis, Hsp70 was also located in the nuclei of keratinocytes, with a decreasing proportion of positive nuclei towards the MET. This keratinocyte nuclear labeling of Hsp70 is also found in cholesteatoma (Shinoda and Huang 1996), under various stressful conditions in vitro and in situ (Muramatsu et al. 1992; Charveron et al. 1995). The literature is abundant regarding the possible chaperone function of Hsp70 (Gething and Sambrook 1992; Hendrick and Hartl 1993; Frydman et al. 1994). The expression of Hsp70 in basal and low suprabasal cells was located behind the MET, where the expression of Hsp60 was at its maximum. Therefore, they could act together as molecular chaperones in response to keratinocyte proliferation.
Several studies have been conducted to understand the migration of epithelial cells over the wound surface (Vaughan and Trinkaus 1966; Krawczyk 1971; Winter 1972; Pang et al. 1978; Buck 1979; Radice 1980; Ortonne et al. 1981; Paladini et al. 1996). Most have not made a clear distinction between the origin of cells at the leading edge (basal or suprabasal origin) and the mechanism of migration (rolling of cells over each other or sliding as a coherent sheet). The observed expression of Hsp60 and Hsp70 by keratinocytes behind the MET vs in the MET reinforces the concept of the suprabasal origin of the MET. Behind the MET, a gradient of labeling was observed between basal cells (high expression) and high suprabasal cells (light expression). In the MET, an intermediate intensity was observed. Our study therefore suggests that keratinocytes of the MET originate from low suprabasal layers behind the MET.
In summary, the present study shows that the wound environment, which is believed to be stressful for cells, affects the expression of Hsps in a more complex manner than expected. Instead of being in direct relation to the time after wounding, the expression of Hsps is dependent on the location of keratinocytes in the regenerating epidermis. However, the variations observed along the neoepidermis might be due to the wound environment as the healing process progresses in the granulation tissue from the wound margin to its center. Each Hsp shows a specific pattern of expression in the regenerating epidermis instead of showing a unique pattern of response to the wound environment. This might be explained by their different functions: Hsp60 and Hsp70 were associated with proliferation in the TE, whereas the expression of Hsp27 and Hsp90 was correlated with the differentiation of keratinocytes. As discussed, the patterns of expression of these Hsps are probably not unique to wound healing because similar observations have been reported in other pathologies. The present data should help in our understanding of the physiological significance of the yet unknown roles of Hsps in living organisms.
Footnotes
Acknowledgments
Supported by grants from the Medical Research Council of Canada (MRC) to FAA, JL, RMT, and LG. AL is a recipient of a MRC studentship and FAA and LG are recipients of scholarships from the Fonds de Recherche en Santé du Québec (FRSQ).
We thank Julie Bergeron and Julie Tremblay for technical support and Dr Pierre A. Coulombe for the anti-keratin 6 antibody. We are grateful to Claude Marin for the photographs.