
Abstract
Human ATPase (hASNA-I) is a novel human gene recently cloned on the basis of homology to the arsA gene of bacteria. Its protein product is an ATPase that is free in the cytoplasm and bound in the perinuclear area and nucleolus in human cells. We prepared the hASNA-I-specific 5G8 monoclonal antibody and used it to investigate the expression of hASNA-I in normal human tissues and breast cancers. hASNA-I was detected immunohistochemically only in the epithelial cells of the liver, kidney, and stomach wall, in the adrenal medulla, in the islet cells of the pancreas, in the red pulp of the spleen, and in cardiac and skeletal muscle. No staining was observed in the uterus, testis, lung, thyroid, cerebellum, and large intestine. Although no staining was also observed in normal breast tissue, all four cases of breast fibroadenomas and all 15 cases of either primary or metastatic breast carcinoma demonstrated increased staining. No embryological or functional common denominator is readily apparent. However, the increased expression in malignant breast cells is of particular interest with respect to the use of this antibody for screening of cytological specimens.
Keywords
R
The ArsA protein is a member of a superfamily of ATP binding proteins with a nucleotide binding motif distinct from that of other ATPases (Koonin 1993). In the course of our studies of resistance to toxic metal in human cells, we cloned the human arsenite-stimulated ATPase (hASNA-I) cDNA based on DNA sequence homology to the distinct nucleotide binding motif (Kurdi-Haidar et al. 1996). The human hASNA-I is an ATPase. However, although it is stimulated by arsenite, unlike its bacterial counterpart it has significant basal ATPase activity even in the absence of oxyanions (Kurdi-Haidar et al. unpublished observations). Structural and biochemical characteristics of hASNA-I suggest that it is functionally different from the ArsA protein of E. coli. To investigate the role of hASNA-I in the physiology of human cells, we produced the specific anti-hASNA-I mouse monoclonal antibody 5G8. Immunocytochemical and Western blot analysis of subcellular fractions identified a soluble pool of hASNA-I in the cytoplasm and bound pools at the nuclear membrane and in the nucleolus in human cells (Kurdi-Haidar et al. in press). This distribution suggested that hASNA-I plays a role in nucleocytoplasmic transport in human cells. We report here an immunohistochemical study of the expression of hASNA-I in normal human tissues and the discovery that hASNA-I levels are markedly increased in breast fibroadenomas and carcinomas.
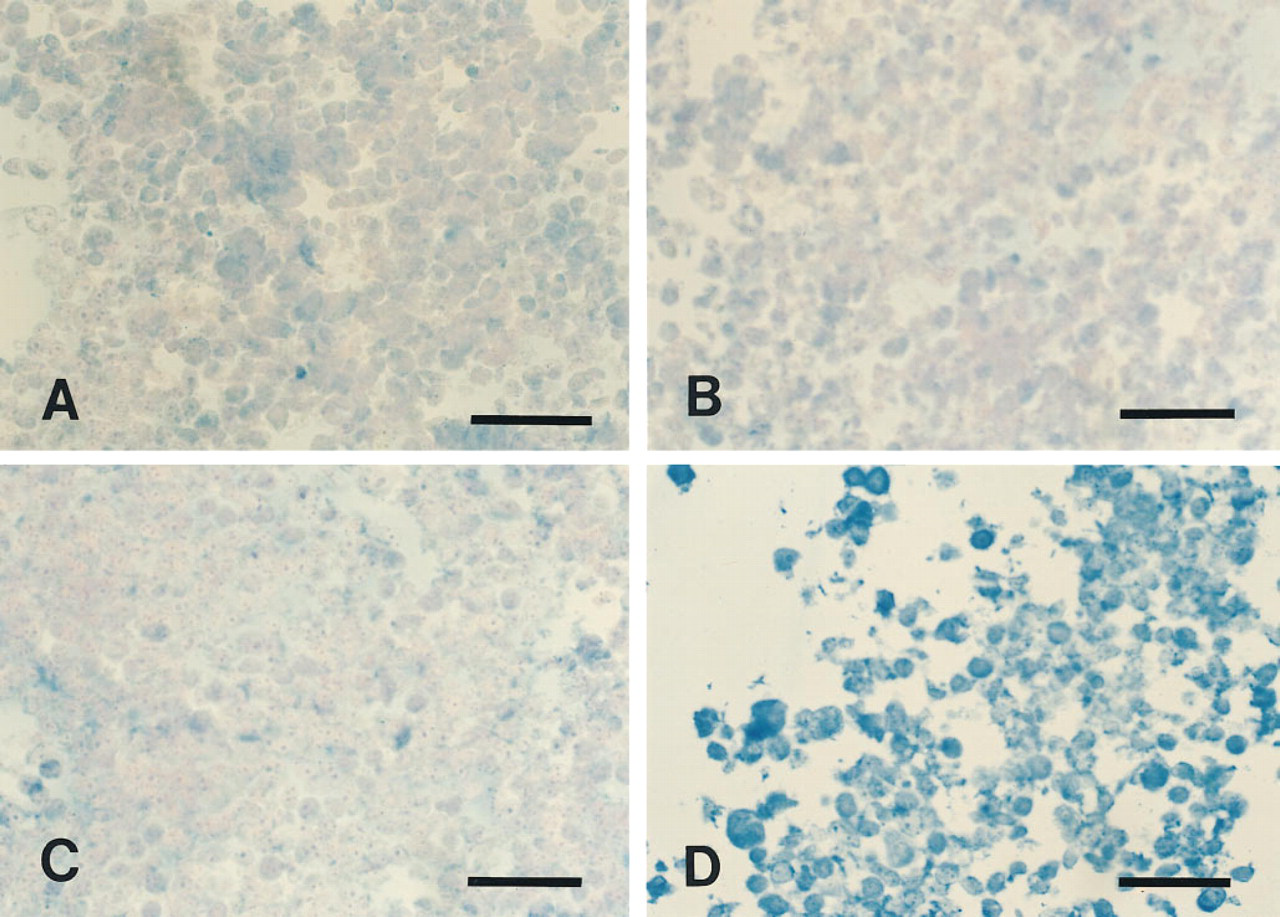
Immunohistochemical detection of overexpression of hASNA-I in human cells. Control T289/E cells stained with purified mouse IgM (
Materials and Methods
Immunohistochemistry
A multitissue block, containing a variety of normal formalin-fixed and paraffin-embedded tissue sections containing five normal sections of each tissue type, was purchased from Dako (Carpinteria, CA). A tissue block containing samples of normal breast, breast fibroadenomas, and primary and metastatic infiltrating ductal carcinomas was obtained from H. Batifora (City of Hope, CA). In addition, sections from four normal breasts and four infiltrating ductal breast carcinomas were obtained from the Tissue Collection and Distribution core laboratory at the UCSD Cancer Center. This study was conducted using 5G8, an IgM anti-hASNA-I mouse monoclonal antibody that was obtained from hybridoma tissue culture medium. This antibody was previously shown to detect a single protein band in human cell lysates by Western blotting, and its further characterization has been reported (Kurdi-Haidar et al. in press). The immunohistochemical staining of the formalin-fixed, paraffin-embedded sections was enhanced by heating in a microwave oven for 5 min on high power in 10 mM citrate, pH 6.0 (Shi et al. 1991) to unmask the hASNA-I epitope recognized by the monoclonal 5G8 antibody. Sections were washed and nonspecific binding was blocked with 10% horse serum in Tris-buffered saline (TBS) (50 mM Tris-HCl, 0.9% NaCl, pH 8.0) for 20 min, then incubated overnight at 4C with the primary 5G8 antibody at a 1:100 dilution in the blocking solution, followed by three 10-min washes in TBS. The mouse monoclonal 5G8 antibody was then localized using the labeled strepavidin-biotin system provided in the alkaline phosphatase Dako LSAB2 Kit. Incubation with the anti-mouse biotinylated “link” antibody was carried out for 10 min and was followed by three 5-min washes in TBS. A second 10-min incubation in alkaline phosphatase-conjugated strepavidin was also followed by three 5-min washes in TBS. Alkaline phosphatase was detected using reagents of the alkaline phosphatase substrate Kit III purchased from Vector Laboratories (Burlingame, CA), in the presence of levamisole, an inhibitor of most forms of alkaline phosphatase other than the conjugate intestinal isoenzyme (Vector Laboratories). Nuclei were counterstained with nuclear Fast Red, purchased from Vector Laboratories, and slides were mounted in Hemo-D Cytoseal 60 mount medium obtained from VWR (San Diego, CA).
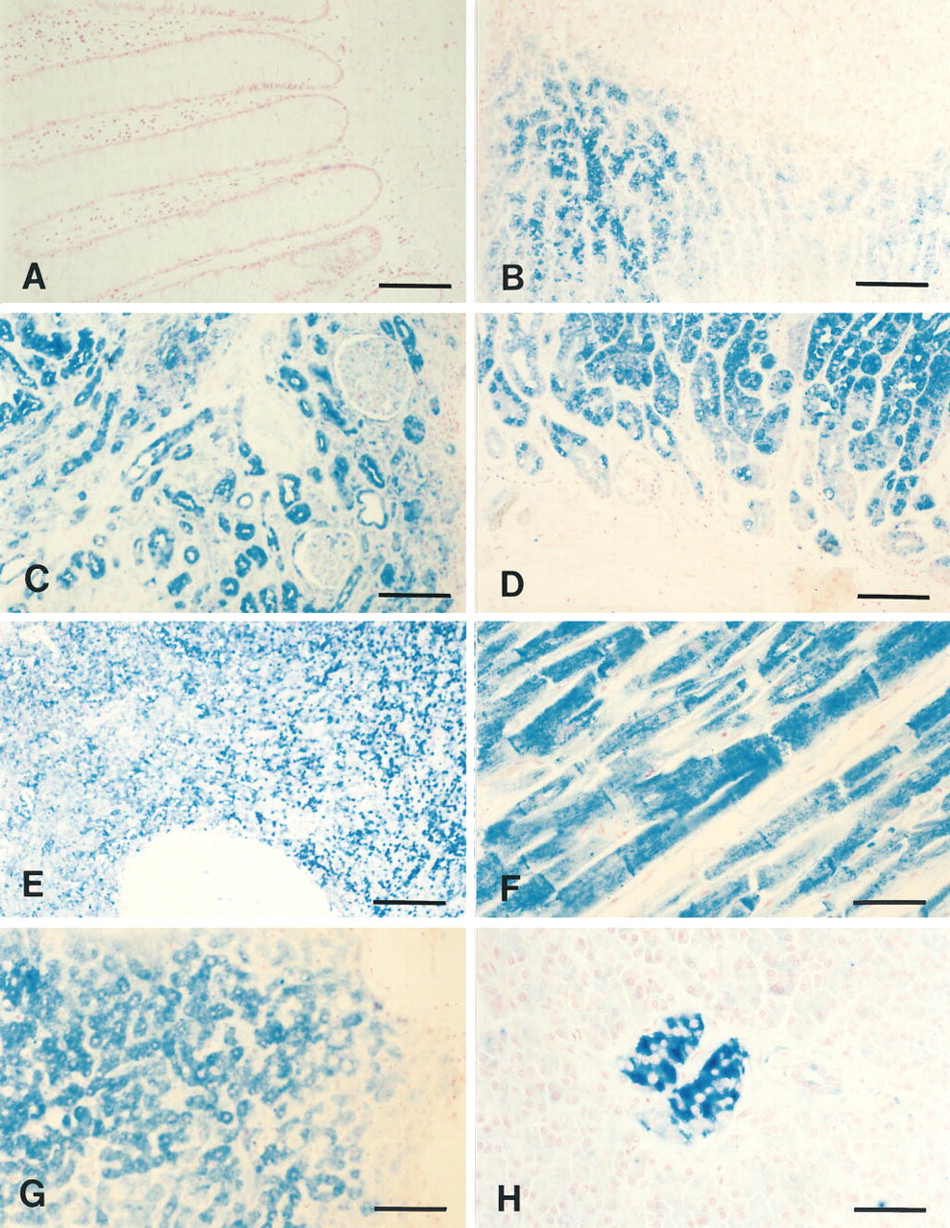
Immunohistochemical detection of hASNA-I in normal human tissues. Immunonegative cells of colon (
Control Cell Lines
A subline of the human malignant melanoma T289 cells (Taetle et al. 1987) overproducing the hASNA-I protein (T289/A) and its empty vector-infected counterpart (T289/E) were generated by retroviral infection using vectors LAPONL (Kurdi-Haidar et al. unpublished results) and LPONL (Yee et al. 1994), respectively. Both sublines were maintained in the presence of 0.4 μg/ml G418 in culture medium (Kurdi-Haidar et al. in press). Tissue culture cell pellets were washed with PBS, fixed in 10% buffered formalin, and paraffin-embedded using standard protocols. Sections were stained either with 5G8 or with control purified mouse IgM (PharMingen; San Diego, CA) at a concentration of 0.05μg/ml.
Results
Immunohistochemical Staining of Cells Overproducing hASNA-I
The 5G8 anti-hASNA-I mouse monoclonal antibody was used to stain formalin-fixed, paraffin-embedded T289/E and T289/A cells (Figure 1). No significant staining was observed in T289/E cells expressing a basal level of hASNA-I with either 5G8 or the control purified mouse IgM (Figures 1A and 1C, respectively). In T289/A cells that were molecularly engineered to overproduce hASNA-I, only background staining was observed with the control IgM antibody (Figure 1B), whereas strong immunostaining was observed with 5G8 (Figure 1D). This result indicated that 5G8 is suitable for immunohistochemical analysis of hASNA-I expression in formalin-fixed, paraffin-embedded tissues.
Immunohistochemical Staining of Normal Human Tissues
The 5G8 antibody was used to immunohistochemically detect hASNA-I in a variety of paraffin-embedded human tissues. Expression of hASNA-I was not ubiquitous but instead was limited to specific tissues and to limited types of cells in the tissues. hASNA-Iimmunoreactive tissues included liver, pancreas, stomach, spleen, kidney, heart, and skeletal muscle, as well as the adrenal gland. No detectable hASNA-I immunoreactivity was found in breast, cerebellum, thyroid, uterine epithelium, testis, lung, or in the large or small intestine (data not shown). Figure 2A shows negative staining of colon epithelium. In the adrenal gland (Figure 2B), the chromaffin cells of the medulla were distinctly positive and no staining was observed in any of the cortical layers. In the kidney, the proximal and distal tubules and the epithelial cells of Bowman's capsule in the cortex were hASNA-I positive (Figure 2C). In the stomach, no staining was detected at the luminal edge of the glands but the epithelial cells lining the neck and crypt were positive, with an apparent differential staining between the parietal and chief cells (Figure 2D). In the spleen, only the red pulp stained for hASNA-I. No staining was detected in the white pulp (Figure 2E). The identity of the hASNA-I-immunoreactive cells in the red pulp of the spleen could not be further ascertained using these sections. In cardiac muscle, both the fibers and the intercalated discs were strongly positive (Figure 2F), and so were the fibers of skeletal muscle (data not shown). In the pancreas, staining was limited to the islets of Langerhans, and the β-cells stained more intensely than the α-cells (Figure 2H). All hepatocytes were hASNA-I-positive, although differences in staining intensity were observed among individual cells (Figure 2G). In all tissues in which staining was detected, the hASNA-I was cytoplasmic.
Immunohistochemical Staining of Breast Tissues
As a first step towards determining how hASNA-I is altered by malignant transformation, we examined multiple breast tissue sections from four normal breasts, four breast fibroadenomas, 13 infiltrating duct carcinomas in the breast, and metastatic deposits of two other infiltrating duct carcinomas. No immunostaining for hASNA-I was found in the duct or lobular epithelium of any of the normal breast control sections (Figures 3A and 3B). Detectable hASNA-I immunostaining was observed in all four of the fibroadenomas examined, ein both the epithelial and myoepithelial cells. There was some variability in staining intensity; sections of two representative fibroadenomas are shown in Figures 3C and 3D. All of the carcinomas stained positively for hASNA-I; representative sections are shown in Figures 3E and 3F. The two metastatic carcinoma deposits examined were strongly hASNA-I-immunoreactive (Figures 3G and 3H). Thus, all breast tissue sections containing either adenomatous or carcinomatous cells demonstrated elevated levels of hASNA-I detectable by immunocytochemistry using the antihASNA-I mouse monolonal 5G8 antibody.
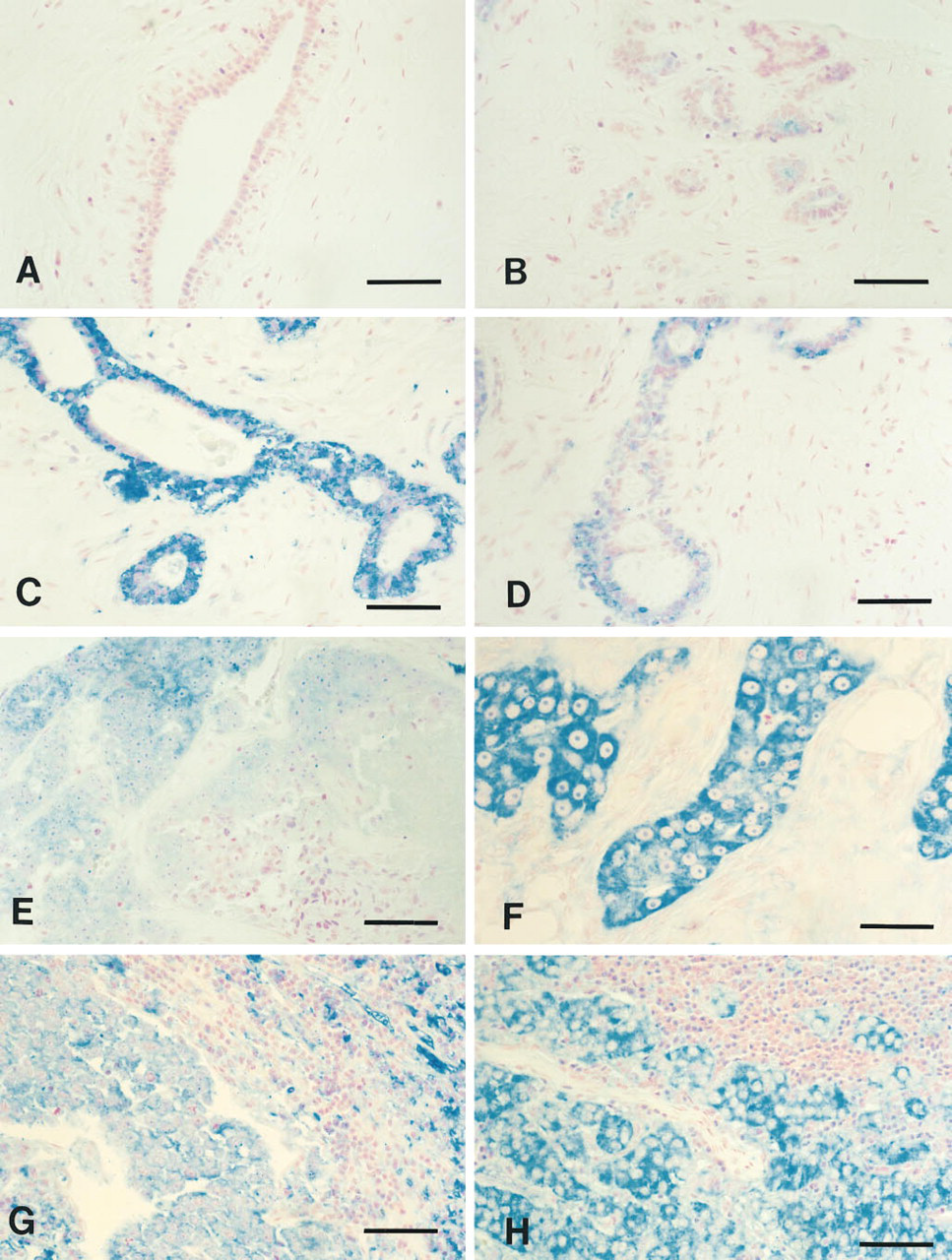
Immunohistochemical detection of hASNA-I in human breast. Immunonegative cells of normal breast (A,B). Immunopositive cells shown are (
Discussion
hASNA-I is a novel human ATPase whose cDNA we isolated as a homologue of the E. coli ArsA, which in bacteria regulates the transport of arsenite and related metalloids. Insight into the role of hASNA-I in the physiology of human cells was derived from a number of findings relating to the ubiquitous expression of its mRNA (Kurdi-Haidar et al. 1996), its distinct biochemical characteristics, and its subcellular localization. We have already established that hASNA-I forms complexes whose size is consistent with tetramers (Kurdi-Haidar et al. unpublished observations), and that it is subcellularly distributed in the cytoplasm, nuclear membrane, and nucleolus (Kurdi-Haidar et al. in press). The fact that it is not found in the plasma membrane suggests that hASNA-I is a paralogue rather than an orthologue of ArsA and that it probably plays a different role in human cells than does the ArsA protein in bacteria.
Immunohistochemical staining of normal human tissues was carried out using the specific anti-hASNA-I monoclonal antibody to further elucidate the role it plays in different human organs. This study indicated that different tissues vary in their level of detectable hASNA-I and that staining of immunopositive tissues is cell type-specific. In addition, individual immunopositive cells also differed in their level of hASNA-I.
We have previously reported the presence of hASNA-I mRNA, determined by Northern blot analysis, in a variety of human tissues including heart, brain, lung, liver, skeletal muscle, kidney, and pancreas (Kurdi-Haidar et al. 1996). Concordant distribution was found by immunohistochemistry, with at least some cells that stained positively for hASNA-I with 5G8 in heart, liver, kidney, and pancreas. The apparent lack of concordance in lung is unexplained, although bronchi and vascular structures larger than arterioles were not present in the sections examined in this study. Additional portions of brain will need to be examined immunohistochemically to determine the source of hASNA-I message in this organ.
Immunohistochemical staining showed that hASNA-I is selectively present in specific cell types for which no embryological or functional common denominator is readily apparent. Among endocrine cells, staining was observed in the chromaffin cells of the adrenal medulla and the islet cells of the pancreas. Specific epithelial cells in the glands of the stomach wall and kidney demonstrated strong staining, whereas other parts of the same epithelial structures showed undetectable staining for hASNA-I. Localization to the intercalated disks in cardiac muscle was particularly striking.
The finding that hASNA-I levels are markedly increased in fibroadenomatous and carcinomatous lesions of the breast is of potential interest for the cytological detection of breast cancer. A more thorough examination of the various subtypes of breast malignancies and of nipple and needle aspiration cytology specimens from these cases is needed. It will also be of interest to determine the molecular mechanisms that underlie this overexpression of hASNA-I and how they are linked to the transformation process.
Footnotes
Acknowledgments
Supported in part by Grants CA69004 from the National Institute of Health and 2RB-0125 from the California Breast Cancer Research Program.
We thank M.A. Lawrence for technical assistance.