
Abstract
Ectonucleoside triphosphate diphosphohydrolases (NTPDases) are membrane-bound ectoenzymes that hydrolyze extracellular nucleotides. We investigated the distribution of NTPDase1 and NTPDase2 in murine salivary gland and pancreas. Histochemistry and immunostaining (by both light and electron microscopy), combined with functional assays, were used to describe the localization patterns and enzyme activities in the organs of wild-type and NTPDase1/cd39-null mice. Pancreatic acinar cells and salivary gland acinar/myoepithelial cells were positive for NTPDase1 and NTPDase2. Ecto-ATPase activity was slightly higher in salivary glands. Ductal epithelial cells expressed ecto-ATPase activity but NTPDase1 and NTPDase2 expression were weak at best. ATPase activity was found in blood vessels of both tissues and its localization pattern overlapped with NTPDase1 staining. In these structures, NTPDase2 antibodies stained the basolateral aspect of endothelial cells and the supporting cells. Biochemical assays and histochemical staining showed relatively high levels of ATPase activity in both glands of cd39−/− mice. Our data therefore support a physiological role for NTPDase2 and other ectonucleotidases in the pancreas and salivary glands. Because NTPDase1 is expressed in non-vascular cell types, this finding suggests that NTPDase1 may have functions in the gastrointestinal tract that differ from those demonstrated in the vascular system.
S
Ectonucleotidases that catalyze the hydrolysis of extracellular nucleotides could play a strategic role in modulating these nucleotide-mediated processes. Ectonucleoside triphosphate diphosphohydrolase-1 (NTPDase1), previously identified as ATP diphosphohydrolase (ATPDase; EC 3.6.1.5) or CD39, is an important ectonucleotidase responsible for the sequential hydrolysis of β-and 7-phosphates of tri- and di-phosphonucleosides. It therefore modulates the concentrations of extracellular nucleotides in a variety of physiological systems. For example, NTPDase1 has important functions in the control of blood hemostasis and thrombosis (Kaczmarek et al. 1996; Marcus et al. 1997; Enjyoji et al. 1999). NTPDase2, also named ecto-ATPase or CD39L1 has been shown in vitro to promote platelet aggregation indirectly by converting ATP to ADP, which is a specific agonist of P2Y1 and P2Y12 receptors (Sevigny et al. 2002).
An ATPDase has been purified and characterized from the zymogen granule membrane of pig pancreas (LeBel et al. 1980; Laliberte et al. 1982; Sevigny et al. 1995). Immunolocalization was later performed in the porcine digestive system (Sevigny et al. 1998) with an antiserum directed against the apyrase conserved region 4 (ACR-4) of NTPDases (Kaczmarek et al. 1996; Sevigny et al. 1997a). However, on the basis of these results it is difficult to define which NTPDase is in fact expressed. More recently, we have demonstrated expression of NTPDase1/CD39 in normal and transformed human pancreas by immunolocalization techniques (Kittel et al. 2002).
Similar enzymes have been extensively studied in the salivary glands of blood-feeding arthropods, where a role in preventing blood coagulation has been ascribed to them (Ribeiro et al. 1984, 1990; Valenzuela et al. 2001). Interestingly, several of these soluble apyrases have been cloned and shown to lack the ACR in the protein sequence (Valenzuela et al. 1998; Valenzuela et al. 2001) and were therefore separated into a new family of nucleotidases. The localization and role of such enzymes in mammals have not been determined.
Here we describe the distribution of NTPDase1 and NTPDase2 expression and the localization of ATP/ADPase activities in the pancreas and salivary glands of mice. The distribution and cellular localization of NTPDase1 and NTPDase2 in mouse pancreas and salivary gland provides new information regarding their roles in the regulation of nucleotide signaling pathways in the gastrointestinal tract.
Materials and Methods
This study was carried out in accordance with the guidelines of the Institutional Ethical Committee for Experimental Animals.
All chemicals were purchased from Sigma Chemical (St Louis, MO) unless otherwise indicated. Rabbit polyclonal antibody C9F was raised against mouse NTPDase1 (Braun et al. 2000; Heine et al. 2001) and BZ34F was raised against rat NTPDase2 and has been previously described (Dranoff et al. 2002; Sevigny et al. 2002; Vlajkovic et al. 2002; Braun et al. 2003)
Male mice were used for all experiments. CD39-null (Enjyoji et al. 1999) and wild-type mice were originally from the background 129 SVJ × C57 BL/6 backcrossed seven generations onto C57 BL/6 and were compared with wild-type mice.
Brightfield Techniques
Preparation of Tissues for Cryosections. Animals were sacrificed by CO2 inhalation, organs (pancreas and submandibular salivary glands) were removed, embedded in OCT (Miles Diagnostic Division; Elkhart, IN), and immediately frozen in isopentane cooled by a mixture of ethanol and dry ice. Six-μm cryosections were cut, dried, and fixed in ice-cold acetone for 10 min.
Enzyme Histochemistry. Lead precipitation from an enzyme histochemical technique was used for demonstration of ecto-ATPase or ADPase activity (Kittel et al. 2002). After fixation and several rinses in 0.07 M Tris-maleate buffer (pH 7.4), the sections were incubated in a medium containing ATP or ADP (1 mM) as substrate, 2 mM Pb(NO3)2 (capturing agent for the liberated phosphate), 1 mM levamisole (inhibitor of alkaline phosphatases; Amersham, Poole, UK), 1 mM ouabain, (Na+, K+-ATPase inhibitor; Merck, Darmstadt, Germany), 50 μM α,β-methylene ADP (5′-nucleotidase inhibitor), and KCl (5 mM) in Tris-maleate buffer (70 mM, pH 7.4) for 30 min at room temperature (RT). Incubation was followed by three rinses in Tris-maleate buffer. The precipitate was converted to PbS with 1% (NH4)2S (1-min incubation). After rinses in distilled water and haematoxylin staining, the sections were mounted in Aquatex (Merck) and images were obtained on an Olympus CH-30 microscope with an Olympus Camedia C-4040Z digital camera. Control experiments were performed without substrate.
Immunostaining for NTPDase1 or NTPDase2 Polyclonal Antibodies. Sectioning and fixation were carried out as described above. After rinsing with PBS, nonspecific binding sites were blocked with 7% normal goat serum in PBS for 30 min and the sections incubated overnight with C9F or BZ34F antibody at a 1:1000 dilution, at 4C. The staining was performed with Vectastain ABC elite kit (Vector Laboratories; Burlingame, CA) and 3,3′-diaminobenzidine (DAB) was applied as chromogen (Sigma-Aldrich; Oakville, Canada) according to the manufacturer's instructions. After washing with distilled water, sections were counterstained with Harris haematoxylin (Sigma-Aldrich), dehydrated, cleared in xylene, and mounted in Permount (EMS; Warrington, PA). Negative control experiments were performed using the same protocol in which the primary antibody was replaced by its preimmune serum at the same concentration (1:1000) or in the absence of primary antibody.
Investigations at Electron Microscopic Level
Enzyme Histochemistry. For electron microscopic investigation the cerium precipitation method was used. Ecto-ATPase activity was localized as described previously (Kittel et al. 1999). Animals were deeply anesthetized with sodium pentobarbital, then perfused briefly through the ascending aorta with a 0.9% NaCl solution. Perfusion fixation was performed with a cold fixative (3% paraformaldehyde (Merck), 0.5% glutaraldehyde (Taab; Aldermaston, UK), 2 mM CaCl2, and 0.25 M sucrose in 0.05 M cacodylate buffer, pH 7.4) for 30 min. Organs were removed, the samples were washed three times with cacodylate–sucrose buffer (0.25 M sucrose in 0.05 M cacodylate, pH 7.4), and 70-μm vibratome-sections were cut. The sections were rinsed several times in 0.07 M Tris-maleate buffer (pH 7.4), then incubated in a reaction mixture containing 3 mM CeCl3, 5 mM MnCl2, 2 mM CaCl2, 1 mM levamisol, 1 mM ouabain, 50 μM α,β-methylene ADP, and 1 mM ATP or ADP in 0.07 M Trismaleate buffer (pH 7.4) at 37C for 30 min. After washing with Tris-maleate buffer, the preparations were postfixed in 1% OsO4 for 30 min. After washing with distilled water, the sections were dehydrated in graded ethanol, block-stained with 2% uranyl acetate in 70% ethanol for 1 hr, and embedded in Taab 812. Ultrathin sections were examined in a Hitachi 7100 transmission electron microscope (Hitachi; Tokyo, Japan). Control reactions were performed without substrate ATP or ADP.
Immunohistochemistry. Animals were anesthetized as above. Fixation was carried out by intracardiac perfusion with PBS (137 mM NaCl, 3 mM KCl, 15 mM Na+/K-phosphate buffer, pH 7.4), 3% paraformaldehyde, and 0.5% glutaraldehyde, pH 7.4, at 4C. Vibratome sections were cut (70 μm thick), rinsed with PBS, and incubated in blocking solution (PBS containing 5% normal goat serum, 1 mg/ml bovine serum albumin) for an hour at RT. Sections were incubated with one of the polyclonal NTPDase antibodies (dilution 1:1000 in blocking solution) overnight at 4C. After several washes with PBS at RT, tissue sections were incubated with biotinylated anti-rabbit IgG antibody for 2 hr according to the ABC method and DAB was utilized as chromogen. After washing thoroughly with distilled water, sections were postfixed in 1% OsO4, dehydrated in 70% ethanol, stained with 2% uranyl acetate, and embedded in Taab 812. Negative control experiments were performed using the same protocol but with the NTPDase1 antibody omitted or preimmune serum (1:1000) used instead of the first antibody.
Enzymatic Assays. Murine tissues (see above) were collected and snap-frozen in liquid nitrogen and kept at −80C until used. Protein extracts were prepared as previously described (Sevigny et al. 1998) and tested for enzymatic activity the same day. Enzyme assays were carried out at 37C in 1 ml of 5 mM CaCl2, 80 mM Tris-HCl (pH 7.4) as described previously (Sevigny et al. 1997b). Reactions were started by the addition of 350 μM of the substrate (ATP or ADP) and then stopped with 0.25 ml of malachite green reagent. Inorganic phosphate was estimated by the malachite green technique according to Baykov et al. (1988). Enzyme activity was expressed as nmoles of inorganic phosphate released per min per mg of protein. Protein concentrations were estimated by the method of Bradford using bovine serum albumin as a standard (Bradford 1976).
Results
ATPase and ADPase Biochemical Activities
As shown in Table 1, the ADPase activity was dramatically decreased both in pancreas and submandibular salivary gland in cd39−/−-deficient mice compared with wild-type mice. The ATPase activity was also significantly diminished in cd39−/− pancreas compared with wild-type tissue, albeit to a lesser extent than for the ADPase activity. The ATPase activities were similar in the salivary glands of both wild-type and null mice, indicating that enzymes other than NTPDase1 are responsible for the bulk of the ATPase activity in the salivary glands.
ATPase and ADPase activities in the pancreas and submandibular salivary gland of wild-type and cd39−/− mouse a
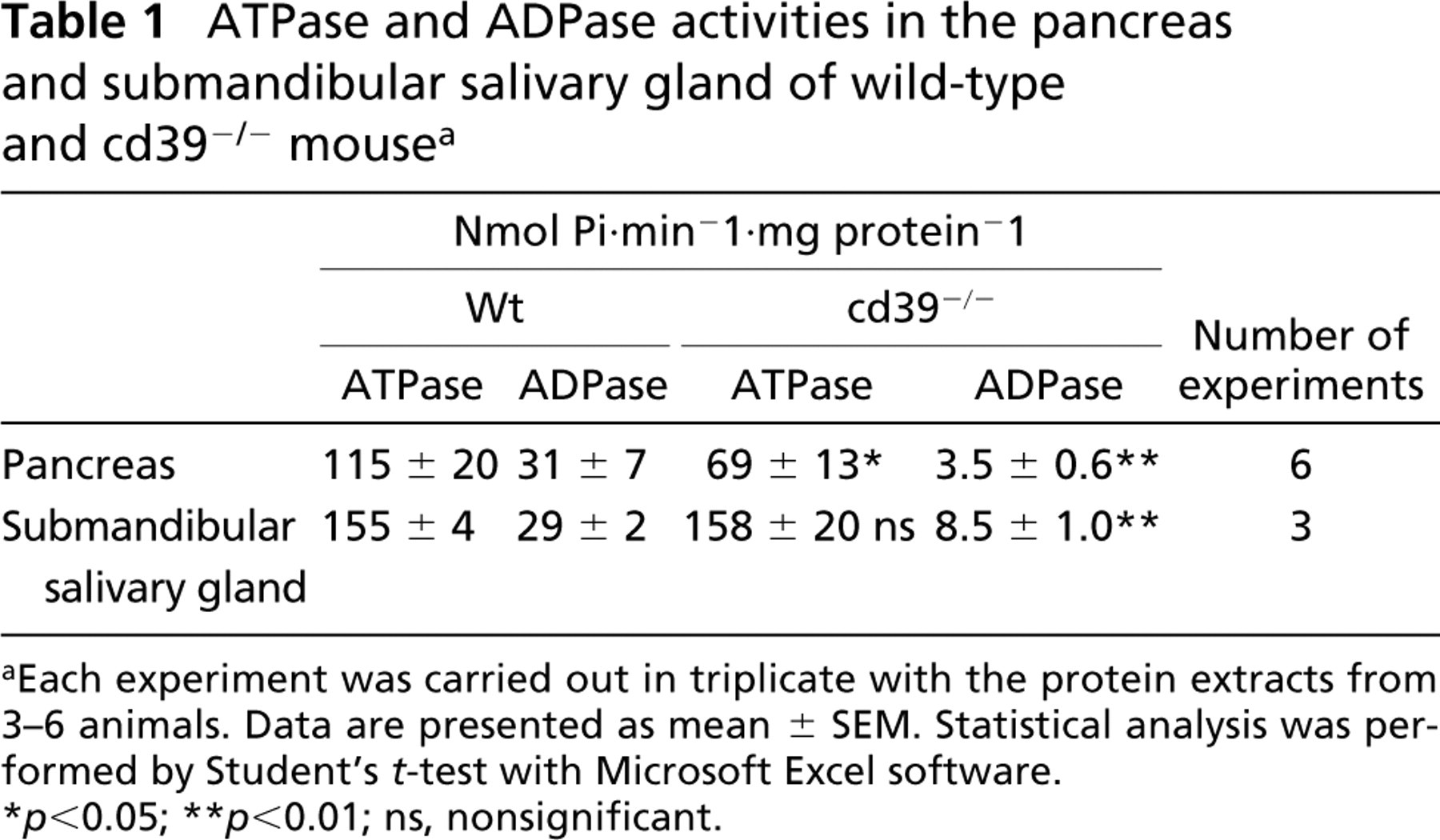
Each experiment was carried out in triplicate with the protein extracts from 3–6 animals. Data are presented as mean ± SEM. Statistical analysis was performed by Student's t-test with Microsoft Excel software.
p<0.05
p<0.01; ns, nonsignificant.
Localization of NTPDase1 in Mouse Pancreas
Immunostaining at the light microscopic level with the anti-mouse NTPDase1 antibody C9F indicated a positive reaction on the endothelial cells of blood vessels of mouse pancreas, while the duct cells appeared negative (Figures 1A and 1B). Positive immunoreactivity was also observed with the NTPDase1 antibody in the acinar lumen (Figure 1C). Under higher magnification, strong staining was present in the plasma membranes of endothelial cells on both basolateral and luminal sides and some immunoreactivity could be seen on the basal side of duct epithelial cells (Figure 1D). With electron microscopic evaluation, some precipitate indicating immunoreactivity could be observed on the basolateral membranes of the acini and in the membranes of some granules (Figure 1E). However, not every acinus contained stained granules (Figure 1F).
A similar distribution was detected by histochemistry for enzyme activity with ATP or ADP as substrate. Brown lead sulfide precipitates indicating ecto-ATPase and/or ADPase activity (not shown) were found on endothelial cells and in the luminar region of the acini (Figure 1G). Electron microscopy showed strong ecto-ATPase activity on the basal foldings of the duct epithelial cells. Plasma membranes of nerve terminals found in the pancreas also revealed high ecto-ATPase activity (Figure 1H). Control experiments without the substrate ATP did not show any precipitate in any of the cell types in the pancreas (Figure 1I). The acini of cd39−/− pancreas did not show any immunoreactivity with the antibody C9F at either light or electron microscopic level (Figure 1J). This confirmed the specificity of the antibody and the absence of the enzyme in these mice. Some precipitate indicating ecto-ATPase activity was still present in traces in cd39−/− pancreas, suggesting the presence of other ectonucleotidases (Figure 1K).
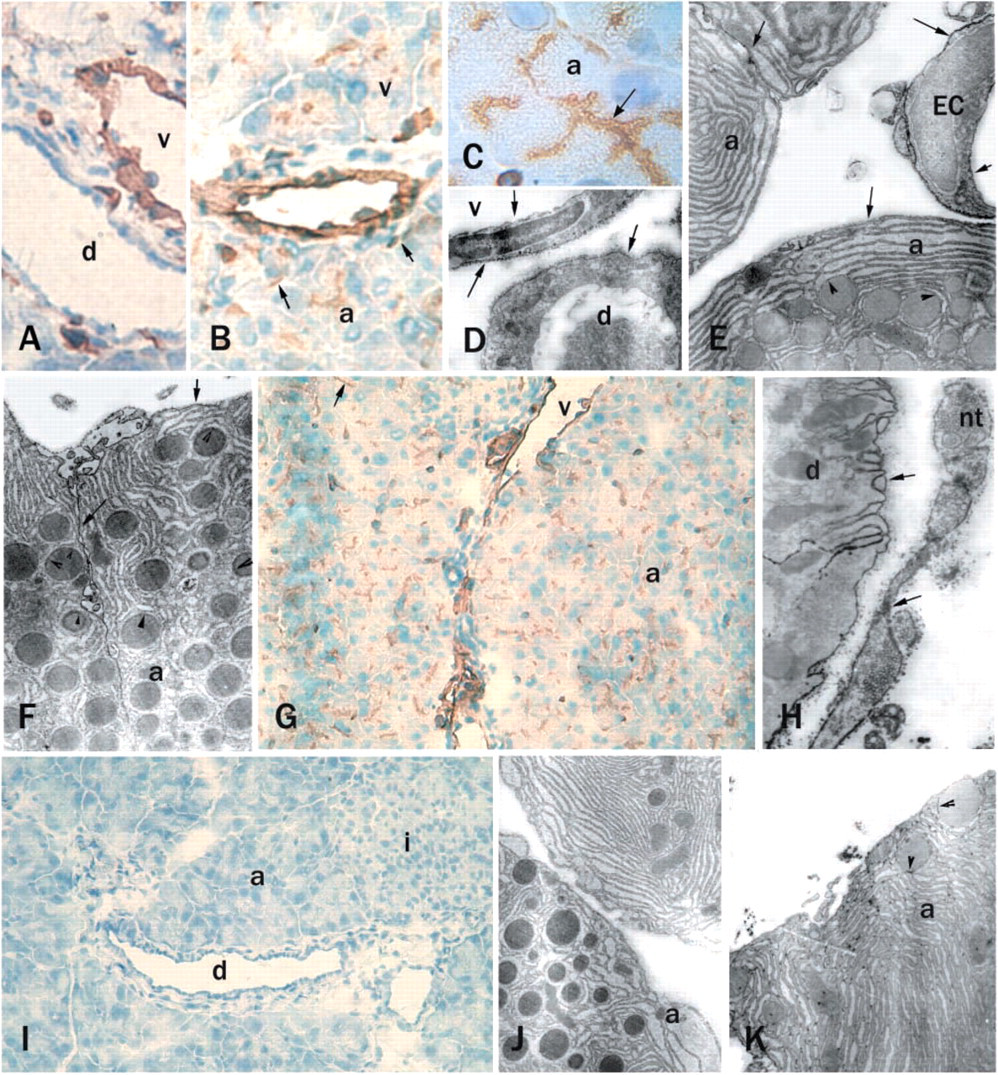
CD39/NTPDase1 immunostaining and enzyme histochemistry for ATPase activity in mouse pancreas. Rusty brown color of DAB (diaminobenzidine) indicates the immunoreactive cells on cryosections (
Localization of NTPDase1 in Mouse Submandibular Salivary Gland
At light microscopic level, similar staining to that seen in pancreas was observed. Weak NTPDase1 staining was detected among mucous acini, whereas duct epithelial cells were negative. Brown deposits showed the immunoreactivity of blood vessels (Figure 2A). Electron microscopy gave a more detailed picture of the localization of this reactivity. Many caveolae containing precipitate were visible on the basolateral aspect of the endothelial cells. Some finely dispersed precipitate was also seen on the basal membrane of the acini (Figure 2B). An acinar cell and its adjacent myoepithelial cell are shown in Figure 2C. This tight contact did not enable us to identify which membrane was stained. However, strong immunoreactivity was found in every case in the caveolae of the myoepithelial cells (Figures 2C and 2D). The plasma membrane of duct epithelial cells also showed some immunoreactivity (Figure 2D).
At light microscopic level, enzyme histochemical staining showed ecto-ATPase activity in blood vessels, in cells surrounding the ducts, and in traces around the acini (Figure 2E). Electron microscopic enzyme histochemistry revealed widespread ecto-ATPase activity. Basal membranes of acini and myoepithelial cells and endothelial cell membranes expressed ecto-ATPase activity (Figure 2F). As in the pancreas, plasma membranes of nerve terminals and membranes of nerve fibers in the salivary gland also showed strong ecto-ATPase activity (Figures 2G and 2I). The membranes of the acini, as well as some of their granule membranes, had some ATPase activity but not consistently (Figure 2H). In most cases acinar membranes were heavily stained and their granule membranes were unstained (Figure 2I). The localization pattern of ecto-ADPase activity in the cells of the wild-type mouse salivary gland was similar to that of the ecto-ATPase activity (Figures 2J and 2K).
As expected, cd39−/− mouse salivary gland cells showed no immunoreactivity for NTPDase1 antibody (Figure 2L), but enzyme histochemical staining demonstrated some remaining ATPase activity in the myoepithelial cells and acini (Figure 2L, inset). In the absence of substrates (ATP or ADP) there was no staining (Figure 2M).
Localization of NTPDase2 in Mouse Pancreas and Submandibular Salivary Gland
In cryosections of mouse pancreas, blood vessels exhibited strong immunoreactivity for NTPDase2. Ducts were also stained, and some staining was visible in the luminal area of the acini (Figure 3A). Electron microscopic investigation showed immunoreactivity on the basolateral aspects and in the caveolae of endothelial cells (Figurs 3B and 3E). Staining on the basal side of the acini was of variable intensity (Figures 3B, 3C, and 3E). In contrast to the light microscopic finding, electron microscopy did not show immunoreactivity for NTPDase2 in the luminal region of the acini (Figure 3D).
In cryosections of salivary gland, immunostaining could be seen in blood vessels and in traces around acini (Figure 3F). Electron micrographs show the staining at higher magnification. The myoepithelial cells also had some immunoreactivity. Strong immunoreactivity was found on the plasma membranes of nerve terminals (Figures 3G). The epithelial cells of the ducts were negative for the NTPDase2 antibody (Figure 3H). Two types of acini (mucinous and serous) straddle a capillary in Figure 3I. Only the basal side of the endothelial cells and supporting cells in the vasculature showed some weak immunostaining for NTPDase2 in mouse salivary gland. No staining was detected in the presence of the preimmune serum (Figure 3J).
Table 2 summarizes the enzyme and immunohistochemical staining obtained for selected cell types from pancreas and submandibular salivary gland. In general, the staining for NTPDase1 and NTPDase2 was similar in both tissues. However, whereas NTPDase2 antibodies gave a strong reaction in the pancreatic duct cells, no staining could be detected in the duct epithelial cells of the submandibular salivary gland. Likewise, the acini in the pancreas showed NTPDase2 staining, whereas those in the salivary gland did not.
Discussion
Although the regulatory effect of ectonucleotides on secretion in exocrine glands is undeniable (Hug et al. 1994; Christoffersen et al. 1998), the cellular localization of their hydrolyzing enzymes has not been described thus far. We have therefore investigated the localization pattern of NTPDase1 and NTPDase2 in mouse pancreas and submandibular salivary gland. An enzyme histochemical technique was used to reveal the active enzyme, and immunohistochemistry demonstrated the expression of both active and non-active protein. Biochemical activity measurements showed smaller but still significant ectonucleotidase activities in the pancreas and salivary glands of cd39−/− mice, suggesting the presence of other ectonucleotidases in these organs. Because the ATPase:ADPase ratio was modified in the favor of ATPase activity, the first candidate was a so-called ecto-ATPase, NTPDase2, the presence of which was demonstrated by immunostaining in both organs.
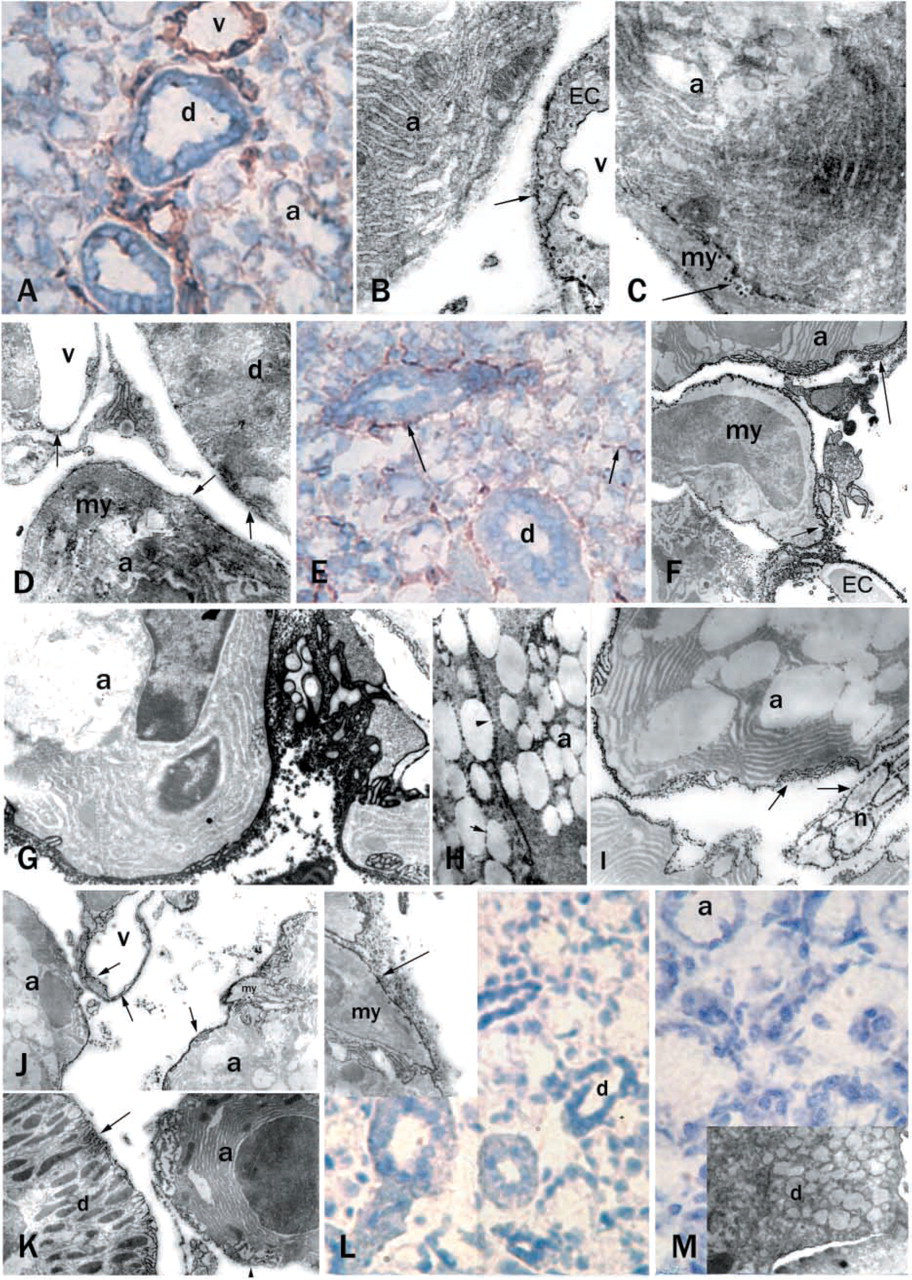
CD39/NTPDase1 immunostaining and enzyme histochemistry for ATPase and ADPase activity in mouse submandibular salivary glands. Rust color of DAB staining indicates the immunoreactive cells on cryosections (
We observed some differences in our LM and EM histochemistry. The differences in enzyme- and immunostaining at light and electron microscopic level in the mouse pancreas could potentially be explained by the high density of proteins in the zymogen granules close to the luminal membranes of the acinar cells. This feature may have masked both the epitopes and the domains responsible for the enzymatic activity. It would be expected that, in vivo, the NTPDases in the zymogen granules would not be active due to the acidic pH and the absence of water. ATPase and ADPase activity and NTPDase1 or NTPDase2 immunostaining may also have been negative because of changes in conformation during fixation. For electron microscopic studies aldehyde fixation is needed, but this treatment aggregates proteins. As a result, NTPDases could be in different conformations and not all epitopes would necessarily be detected by the antibodies.
The heterogeneity of the zymogen granule membranes described in rat pancreas almost two decades ago by Beaudoin and co-workers (1988) could provide another explanation for the occasional NTPDase1 immunostaining of the granules (Figures 1E and 1F). In another case, electron microscopy demonstrated NTPDase2 immunoreactivity at the basal membranes of the acini, while at light microscopic level some staining was also observed in the luminal region after acetone fixation of the crysections. These findings may suggest that this enzyme is expressed at the apical part of the acinar cell but that aldehyde fixation and/or high protein concentration may have prevented binding of the antibody.
Our data and data from previous reports suggest species-dependent variations in the localization of NTPDase1 in pancreas. Like our findings, an immunohistochemical study showed that apical and basolateral membranes of the acinar cells in pig pancreas were immunoreactive for an antibody directed against the common apyrase conserved region 4 (ACR 4) of NTPDase molecules (Sevigny et al. 1998). However, in human pancreatic cryosections, according to electron microscopic staining, acini were devoid of NTPDase1 immunoreactivity or ATPase activity (Kittel et al. 2002). Only the duct cells showed immunoreactivity for NTPDase1. Electron microscopy showed staining was located in the basal foldings of the ductal epithelial cells. Sorensen and colleagues (2003) described similar basolateral staining in the larger ducts of rat pancreas, while CD39/NTPDase1 immunofluorescence staining was localized on the luminal membranes in small intercalated/interlobular ducts. In rat, immunofluorescence showed CD39/NTPDase1 localization on the basolateral membranes of acini and also intracellularly. The differences in the localization of NTPDase1 in duct epithelial cells, depending on the size of the duct, may be of importance. Thus far we have no definitive explanation for this finding, but the localization pattern of NTPDase1 expression on the basal membrane may suggest a close interaction with P2 receptors. Interestingly, the expression level of P2Y1 receptor was found to fluctuate during the development of the pancreas, while the expression levels of some other P2 receptors (e.g., P2 × 7) were constant (Coutinho-Silva et al. 2001).
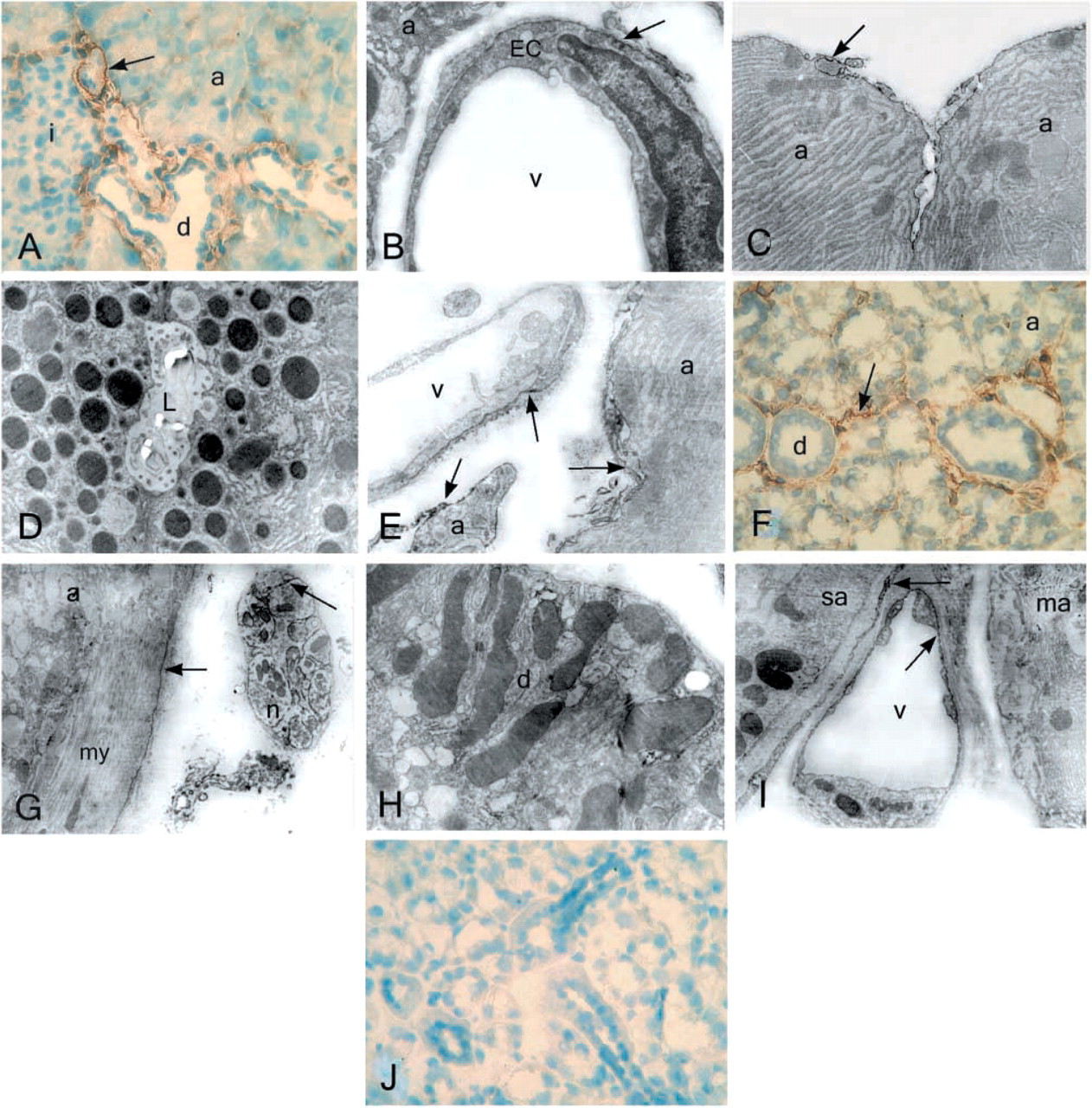
NTPDase2 immunostaining of pancreas (
Summary of enzyme and immunohistochemical staining obtained for selected cell types of pancreas and submandibular salivary gland a
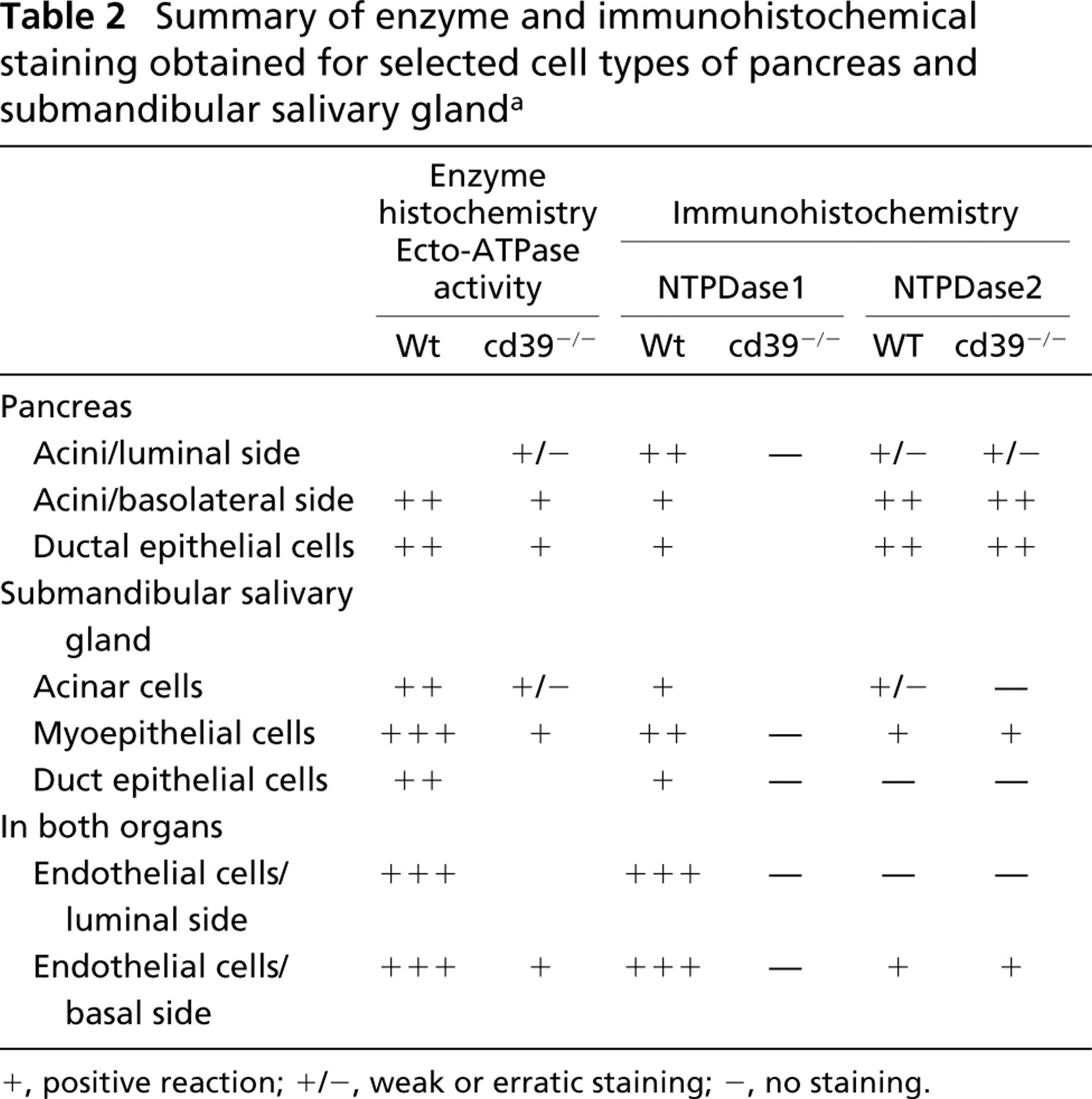
+, positive reaction; +/−, weak or erratic staining; −, no staining.
The effects of ATP on salivary gland cells have been investigated in the past (Thyberg et al. 1982). There is evidence from pharmacological and molecular approaches for the expression of two ligand-gated ion channels, P2 × 4 and P2 × 7, and two G-protein-coupled receptors, P2Y1 and P2Y2, as detected in different salivary epithelial cell lines (Yu and Turner 1991; Gibb et al. 1994; Buell et al. 1996) and in dispersed salivary gland cells (Park et al. 1997; Gibbons et al. 2001). The control of the concentration of the agonists of P2-receptors by NTPDases is an important factor in nucleotide signaling.
Our work is the first to show the expression pattern of NTPDases and ecto-ATPase/ADPase activity in the salivary gland of a mammal. The strongest immunoreactivity and enzyme activity belonged in every case to the membranes of myoepithelial cells and their caveolae. Caveolae are special membrane invaginations abundantly found in endothelial and smooth muscle cells. They have roles in endothelium-dependent relaxation, contractility, and maintenance of myogenic tone, as well as in organizing signaling pathways in the cell. The presence of P2 receptors has also been detected in caveolae (Kittel, unpublished observation). In the present study, acini attached to the myoepithelial cells showed weak immunoreactivity for NTPDase1 antibodies on their basal sides, while ecto-ATPase and ecto-ADPase activity was strong at their basal membrane and infoldings.
In the cd39−/− mouse salivary gland, enzyme histochemical staining demonstrated ATPase activity in the myoepithelial cells and weak activity in the acini. This finding, together with the biochemical activity data, supports the presence of NTPDase2 in this organ. NTPDase2 immunoreactivity was found in the basal membrane of endothelial cells as well as in the supporting cells of the vasculature in cd39−/− salivary glands.
Finally, we must note the strong ATPase and ADPase activities of the endothelial cells of blood vessels. Staining of endothelial cells, as expected, showed the same pattern of immunoreactivity and enzyme histochemical staining as in the pancreas and other tissues (Kittel 1997, 1999; Sevigny et al. 1997b; Braun et al. 2000; Sevigny et al. 2002). Interestingly, caveolae were more numerous in the endothelial cells of salivary gland than of pancreas. The caveolae of endothelial cells were labeled with NTPDase1-specific antibodies in both organs. Recent studies have reported the localization of NTPDase2 in pericytes and fibroblasts in the cardiac vasculature and liver (Dranoff et al. 2002; Sevigny et al. 2002). In addition to this previous finding, we have demonstrated here that NTPDase2 immunoreactivity was also observed on the basal side and in the caveolae of endothelial cells. Some of these variations could be due to the heterogeneity in the vascular endothelium, as previously reported (Ponder and Wilkinson 1983; Pino 1986; Steinhoff et al. 1993). Because NTPDase1 and 2 have different expression patterns in endothelial cells, this suggests the possibility of spatial regulation of nucleotide-mediated signaling in the vasculature, as has been proposed previously (Sevigny et al. 2002).
Taken together, the biochemical and histochemical data presented in this work demonstrate the presence both of NTPDase1 and NTPDase2 in mouse pancreas and salivary gland. Our demonstration of their different distributions in the acinar cells, ducts, and vasculature should help to clarify their respective roles in the regulation of P2 signaling pathways.
Footnotes
Acknowledgements
Supported by the Hungarian Scientific Research Fund (OTKA) T 034722 and M36314 (to AK) and by the Canadian Institutes of Health Research (to JS), the NIH (to SCR), and by the Deutsche Forschungsgemeinschaft (SFB 269, A4; to NB).
We thank Mr Gyozo Goda for excellent technical assistance and the EM-TEK Kft. for their generous support.