
Abstract
The Notch signaling pathway is a vitally important pathway in regulating brain development. To explore the involvement of the Notch pathway in neuronal cells of adult rat gut, we investigated the expression of Notch1 and Jagged2 by in situ hybridization (ISH) and immunohistochemistry (IHC). In the enteric nervous system, Notch1 and Jagged2 were expressed in ganglia of the submucosal and myenteric plexus. Notch1 was preferentially expressed in cholinergic neurons lacking calretinin or nitric oxide synthase (NOS), whereas Jagged2 was present in most neuron subtypes. We propose that Notch1 and Jagged2 have a continuing role in the maintenance and function of neuronal cells in the adult enteric nervous system.
N
Four-month-old female Sprague–Dawley rats were sacrificed and tissue segments from the stomach (fundic and body regions), esophagus (proximal and distal), and duodenum, jejunum, ileum, cecum, and colon (proximal, middle, and distal for each) were removed, fixed, and processed for in situ hybridization (ISH) (Powell et al. 1998). All experiments were approved by the Animal Ethics Committee of the Womens and Childrens Hospital, Adelaide, South Australia.
The Notch1 probe was a 444-bp EcoR1/HindIII fragment from clone SN6-7 which includes the intracellular domain from the transmembrane region to part of the ankyrin repeats (Weinmaster et al. 1992). The Notch2 probe was a 2462-bp fragment from clone H10-6, including some EGF repeats, the transmembrane region and some of the ankyrin repeats (Weinmaster et al. 1992). The Jagged1 probe was a 527-bp BglII fragment spanning the cysteine-rich domain of the rat clone (Lindsell et al. 1996). The Jagged2 probe was a 1076-bp fragment including several EGF-like repeats and part of the cysteine-rich domain (Powell et al. 1998). All probes were gifts of G. Weinmaster (University of California). The rat β-actin RNA probe was synthesized from pTri-Bactin-125 (Ambion; Austin, TX).
Antisense and sense [α33P]-rUTP radiolabeled ribo-probes were hybridized to tissue sections (Powell et al. 1998). Hybridized sections were washed at a final stringency of 0.1 × SSPE at 65C for 30 min and exposed for 10 days at 4C. Tissues were stained with hematoxylin and images captured under brightfield and darkfield illumination with an Olympus BH2 microscope fitted with an Olympus darkfield condenser (model # U-DCW) and a Sony digital video camera (model #SSC-DC50P), and were analyzed with Image Pro Plus analysis software (Media Cybernetics; Carlsbad, CA).
Whole-mount preparations of myenteric and submucous plexuses were prepared by sharp dissection (Brookes et al. 1992,1995). The following antisera were used for immunohistochemistry: rabbit anti-Notch 1 (Santa Cruz Biotechnology, Santa Cruz, CA; cat# H131, 1:400), rabbit anti-Jagged 2 (Santa Cruz Biotech; cat# C-17, 1:200), sheep anti-nitric oxide synthase (NOS) (gift of P. Emson, Cambridge UK; 1:1000), goat anti-calretinin (SWANT, Bellinzona, Switzerland; cat# CG1, 1:500), rabbit anti-ChAT (gift of M. Schemann, Hannover, Germany; 1:2000), and sheep anti-ChAT (Chemicon, Temecula, CA; cat# AB1582, 1:2000). Secondary antisera were raised in donkey and used as follows: anti-sheep FITC (Amersham, Poole, UK; cat# N1034, 1:50), anti-rabbit CY3 (Jackson Immunoresearch Laboratories, West Grove, PA; cat# 711-165-152, 1:400), anti-sheep FITC (Jackson; cat# 713-095-147, 1:200), anti-sheep CY3 (Jackson; cat# 713-165-147, 1:200), anti-goat FITC (Jackson; cat# 705-095-147, 1:200), and anti-goat CY3 (Jackson; cat# 705-165-147, 1:400). For double labeling, two primary antibodies from different species were applied simultaneously and visualized with a mixture of two species-specific, differently labeled secondary antibodies. No specific expression was detected when primary antibodies were omitted. In addition, all labeling was abolished by pre-incubation of the antiserum with a Jagged2 blocking peptide (Santa Cruz Biotech; cat# SC-8157). Specimens were incubated overnight in primary antisera, rinsed in PBS, incubated in secondary antisera for 2–4 hr, rinsed in PBS, and mounted in bicarbonate-buffered glycerol (pH 8.6). Specimens were viewed on an Olympus AX70 microscope using the appropriate filters. These experiments were approved by the Animal Ethics Committee of Flinders University (Adelaide, South Australia).
Notch1 and Jagged2 but not Notch2 or Jagged1 (data not shown) mRNA transcripts were expressed in ganglia distinguished by large nuclei in the myenteric and submucosal plexus throughout the GI tract (Figure 1). Because expression was the same in ganglia throughout the GI tract, only the duodenum is shown.
To more precisely determine the types of enteric neurons expressing Notch1 and Jagged2, IHC localisation was carried out on whole-mount preparations of the duodenum. Notch1 immunofluorescence was visible in ganglia of the myenteric plexus, in nerve fibers in the secondary branches in the circular muscle layer, and in branches of the tertiary plexus in the longitudinal muscle layer (Figure 2). There was general fine punctate staining of myenteric nerve cell bodies. Immunofluorescence was localized to the cytoplasm and was not present in the nucleus nor visibly associated with the cell membrane. To test whether the differential Notch1 labeling was present in all neuronal subtypes, we double labeled preparations with anti-sera that distinguish major subtypes of neurons in the rat enteric nervous system. NOS immunoreactivity was present in about one quarter of myenteric nerve cells, all of which appeared to have a single axon. These are likely to belong to inhibitory motor neurons of the muscularis externa and to descending interneurons (Brookes 2001). There was no overlap between the strongly Notch1-immunoreactive nerve cell bodies and NOS immunoreactivity (Figures 2E and 2F). The calcium-binding protein calretinin was localized in about one third of nerve cell bodies in the myenteric ganglia that showed the typical characteristics of Dogiel type II neurons, with large, smooth somata and several processes arising from the cell body. Again, none of the Notch1-intense neurons was immunoreactive for calretinin in the myenteric plexus (Figures 2C and 2D). Thus, the intensely labeled Notch1-immunoreactive neurons were not randomly distributed among the different classes of enteric neurons but were restricted to a distinct subset of cells. Choline acetyltransferase, a marker of cholinergic neurons, can be detected in the great majority of non-NOS-immunoreactive nerve cell bodies, including the calretinin-immunoreactive neurons. Notch1-immunoreactive nerve fibers in the tertiary plexus and secondary branches in the circular muscle frequently co-localized with ChAT (Figures 2A and 2B). This confirms that Notch1 is present in many neurons in the myenteric plexus but is preferentially expressed in particular subtypes that lack either calretinin or NOS but which are probably cholinergic.
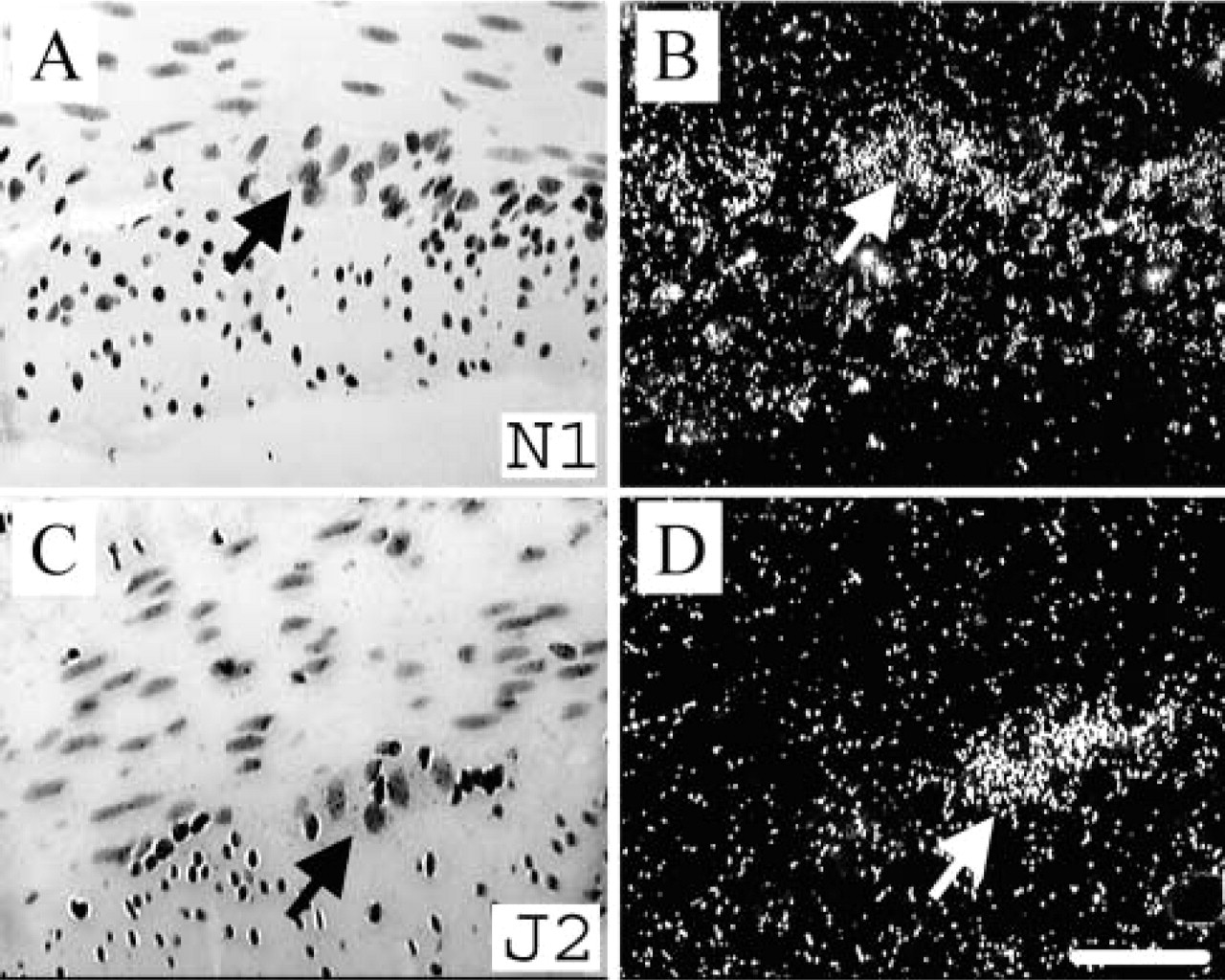
Expression of Notch1 and Jagged2 in the enteric nervous system. Notch1 and Jagged2 were expressed in the myenteric plexus (
Jagged2 immunoreactivity was present in the great majority of nerve cell bodies in the myenteric plexus but was less variable in intensity than Notch1. The labeling was more diffuse than that of Notch1 and was restricted to the cytoplasm. There was no visible labeling of nerve fibers either in the ganglia or in the muscle layers, unlike Notch1. Jagged2 immunoreactivity co-localized with NOS (Figures 2K and 2L), calretinin (Figures 2I and 2J), and ChAT (Figures 2G and 2H), suggesting that it was widely distributed among different functional classes of enteric neurons. This was the case in both myenteric and submucous ganglia.
In embryonic brain development, one of the functions of Notch is to enable differentiation of precursor cells into glial cells (Gaiano et al. 2000). With reports of Notch involvement in neurite outgrowth and dendritic branching, a role for Notch in postmitotic cortical neuronal cells is emerging (Berezovska et al. 1999; Redmond et al. 2000). Our finding that Notch1 and Jagged2 are expressed in non-proliferating ganglion cells in the myenteric and submucosal plexus in the adult GI tract extends the known involvement of Notch signaling to postmitotic neuronal cells outside the brain and, for the first time, highlights the involvement of Notch in the enteric nervous system. Notch1 is expressed in specific types of enteric neurons. It is not present in either NOS- or calretinin-immunoreactive neurons, which together make up about one half of all myenteric neurons in the myenteric plexus of rat duodenum. However, it is present in some of the ChAT neurons lacking these markers. It is likely that NOS-immunoreactive neurons included inhibitory motor neurons and a population of interneurons with aborally directed projections. Calretinin-immunoreactive neurons are likely to be primary afferent neurons with local projections that may run to the underlying mucosa (Mann et al. 1997) or aborally (Brookes et al. 1995). In contrast, ChAT-immunoreactive neurons lacking NOS or calretinin are likely to project locally to the longitudinal muscle (Brookes et al. 1992) or orally to the ganglia or circular muscle layer (Brookes 2001). Therefore, Notch1 may be restricted to neurons with particular polarized projections in the enteric nervous system, where it may be involved in their maintenance.
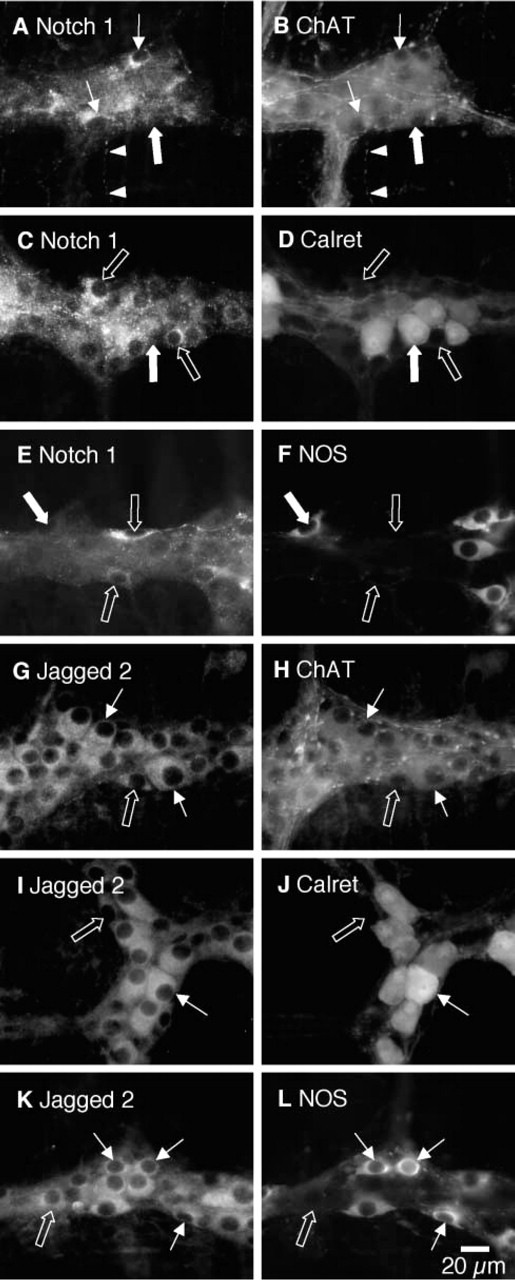
Co-expression of Notch1 and Jagged2 with IHC markers for enteric neurons in the myenteric plexus of rat duodenum. The most intensely labeled Notch1-immunoreactive neurons (arrows,
With the lack of Notch1 expression in neurons expressing Jagged2, it appears that other Notch receptors and possibly ligands not studied in this report may be present in the enteric nervous system.
Footnotes
Acknowledgements
GRS and BCP were funded in part by a Cooperative Research Centre grant from the Australian Government. SJHB was supported by DK56986 of the National Institutes of Health (USA).
We would like to thank Kate Brody for excellent technical assistance with the immunohistochemistry.