
Abstract
Cortactin is a tyrosine kinase substrate that binds to filamentous actin. It represents a highly conserved family of perimembrane signaling proteins. The human homologue of cortactin is encoded by gene EMS1, which is amplified in some human breast, head, and neck carcinomas. This study shows that cortactin is preferentially localized to the apical surfaces of the polarized epithelium, specifically, to the terminal web of small bowel epithelium and to apical surfaces of the proximal renal tubules, thyroid follicles, and bronchiolar epithelium. Cortactin is also seen in cell and tissue types with actin-based contractile capacities, including smooth and striated muscle and myoepithelium.
Keywords
C
This study aimed at a systematic characterization of the expression pattern of cortactin in normal human tissues. Immunohistochemical studies were performed on formalin-fixed, paraffin-embedded normal adult and fetal tissues. Staining was carried out using manual capillary action technology as previously described (Montone et al. 1997). Briefly, after deparaffinization and rehydration, the tissue sections were treated with pepsin (2.5 mg/ml) (Research Genetics; Huntsville, AL) at 50C for 5 min. A specific anti-cortactin mouse monoclonal antibody (MAb) 4F11, which recognizes cortactin proteins in multiple species including human (Wu et al. 1991), was used at a concentration of 20-30μg/ml. A chicken-specific anti-cortactin MAb 1H3 was used at the same concentration as an isotype control. The primary antibody was incubated for 30 min at 40C. The primary antibody was detected with a universal biotinylated secondary antibody (Research Genetics) for 5 min at 45C, followed by incubation with streptavidin-horseradish peroxidase (Research Genetics) at 50C for 5 min, and the signal was generated using diaminobenzidine. The slides were conterstained with hematoxylin, coverslipped, and examined by light microscopy.
Anti-cortactin MAb 4F11 exhibited specific staining patterns in a variety of normal adult and fetal human tissues. In the small bowel, anti-cortactin MAb showed strong staining of the brush border. The staining was especially concentrated at the terminal web of the brush border microvilli (Figure 1A). In contrast, the cell body and the basal and lateral cell surfaces showed no significant staining. In the proximal renal tubules, anti-cortactin MAb also stained the brush border. Interestingly, in the collecting ducts, cortactin was detected only in occasional enlarged cells that were morphologically consistent with the intercalated cells which have microvilli. A similar apical surface staining was also observed in fetal thyroid follicles, which are known to have many microvilli (LiVolsi 1992). More interestingly, anti-cortactin staining was seen in endothelial cells, which are also known to contain microvilli (Figure 1B) (Palade et al. 1979). The endothelial staining was found in the systemic organs and central nervous system. In the lungs, cortactin expression was seen in the ciliated apical surfaces of the bronchiolar epithelium.
The apical staining pattern in the tissues correlates very well with the localization of cortactin in cultured cells, which show an enrichment of the protein in actin-based surface projections, including the membrane ruffles and filopodia of migrating cells and cell-substratum adhesion sites of adherent cells (Wu et al. 1991; Wu and Parsons 1993). Microvilli are cell surface projections containing a core of actin filaments. The formation and maintenance of this highly organized actin-based structure relies on the interaction of actin with specific actin binding proteins and with the cell membrane. On the basis of studies of the brush border microvilli of polarized enterocytes, two actinbundling proteins, fimbrin and villin, are important for the formation of the actin filament core, and the other major actin binding protein, myosin I, links the actin bundle to the plasma membrane (Fath et al. 1993). Although a microvillous core can be assembled in vitro by mixing actin, myosin I, villin, and fimbrin, the in vivo regulation of this process during enterocyte differentiation is not fully understood. Protein tyrosine kinases, especially pp60 c-src and tyrosine phosphorylated proteins, are believed to be important in the regulation of intestinal cell growth (Fath et al. 1993; Berryman et al. 1995). As a major protein substrate of pp60 src and a specific filamentous actin binding protein, cortactin may be an important element in the generation of the brush border microvilli. Future studies are needed to further elucidate the role of cortactin in the morphogenesis of microvilli.
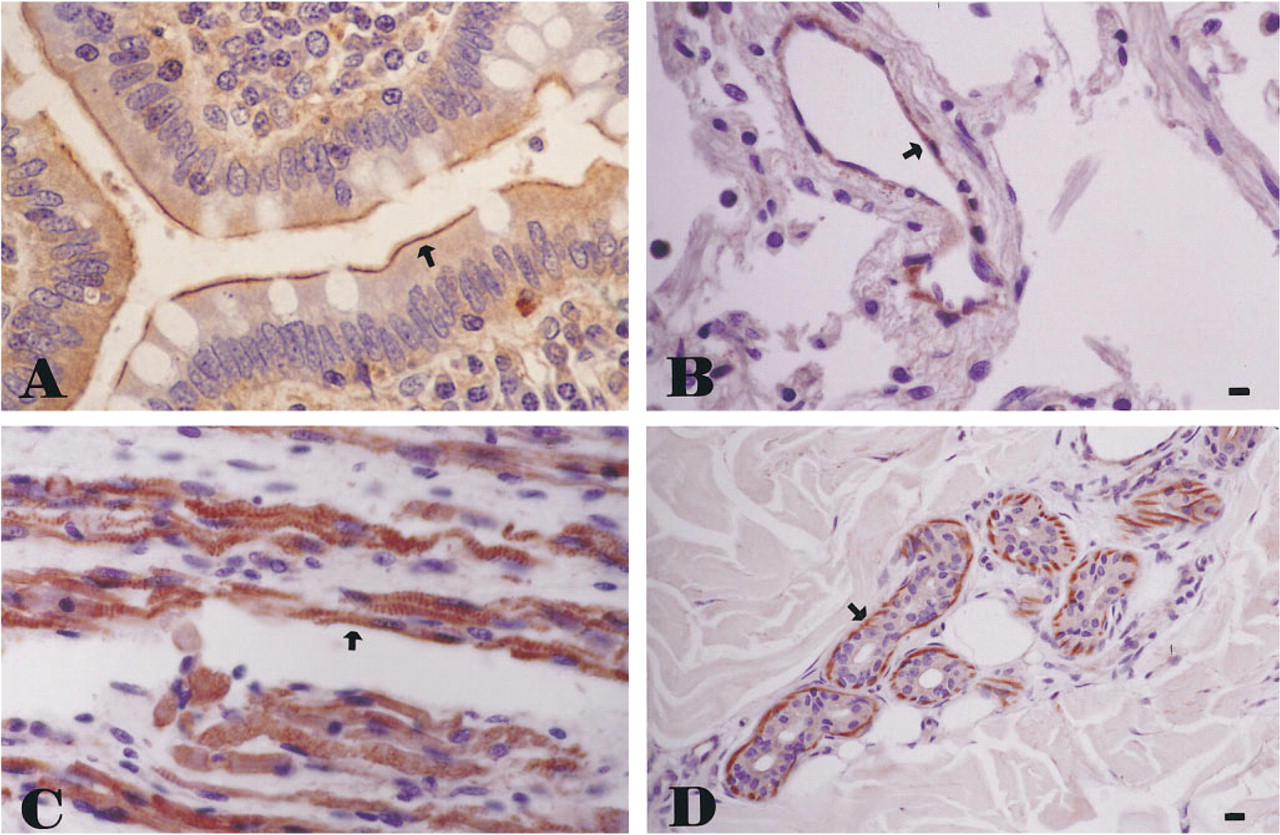
Immunohistochemical stains of normal human tissue using anti-cortactin MAb 4F11 in the small bowel, showing reactivity in the terminal web of brush border microvilli (
Similar to the report by Parham et al. (1995), strong anti-cortactin staining was seen in the smooth muscle of the blood vessels and visceral walls. In addition, cortactin expression was observed in the skeletal and cardiac muscles (Figure 1C). Other cell types with actin-based contractile capacity, including the myoepithelium of the minor salivary glands, sweat glands, and breast lobules, also showed evidence of cortactin expression (Figure 1D). This preferential expression in cells of muscle differentiation may prove to be useful in ancillary testing in a variety of soft-tissue tumors (Parham et al. 1995).
The tissue reactivity of MAb 4F11 seen in the current immunohistochemical study is specific for cortactin. MAb 4F11 shows no crossreactivity with other protein species (Wu et al. 1991). MAb 4F11 recognizes specifically the 80- and 85-kD proteins of cortactin in Western blots using protein extracts from the human kidney, intestine, cerebellum, and hippocampus (data not shown). Moreover, staining of various types of tissue using the isotype control antibody 1H3, which reacts only with cortactin in the chicken (Wu et al. 1991), showed no significant staining.
Footnotes
Acknowledgements
We are grateful to Dr J. Thomas Parsons and his laboratory (Department of Microbiology, University of Virginia) for kindly providing the monoclonal antibodies 4F11 and 1H3. We thank Ms Yunrui Du (University of Virginia) for critical review of the manuscript.