
Abstract
This report describes the subcellular distribution of 5′-nucleotidase (5′-NT) in rat photoreceptor cells and pigment epithelial cells processed by rapid-freeze enzyme cytochemistry. There was a striking difference in the ultrastructural localization of 5′-NT activity between rod outer segments after freeze-substitution fixation and conventional fixation. By rapid-freezing enzyme cytochemistry, 5′-NT activity was localized in the extradiscal space of intact nonvacuolated discs, whereas by conventional cytochemistry it was shown in the intradiscal space of artifactual vacuolated discs. In the freeze-substituted retinal cells, an appreciable difference in functional 5′-NT molecules was also found. The soluble 5′-NT on the cytoplasmic side of the disc membrane was vital in the rod outer segments, whereas the membrane-bound ecto-5′-NT on the exoplasmic (external) surface of the apical process was active in the pigment epithelial cells. Rapid-freezing enzyme cytochemistry should be worth employing as a method to reveal the fine localization of enzyme activity at the level of cell ultrastructures, which are poorly preserved by conventional fixation, and should provide information approximate to that in living cells.
Keywords
E
In retinal photoreceptor cells, the enzyme 5′-nucleotidase (5′-NT) is one of the enzymes in the cGMP metabolism cascade of the vertebrate visual transduction system. Although some researchers have studied the enzyme cytochemical localization of 5′-NT in retinal rod cells processed by conventional fixation (Kreutzberg and Hussain 1984; Hussain and Baydoun 1985; Saito 1991), they have not been in good agreement on its localization. One of the reasons is that rod cells are one of the most delicate cells to treat by conventional fixation and morphological modifications during fixation are often encountered. Yamada (1982) showed the fine structure of intact discs in the rod outer segment processed by rapid-freezing and subsequent freezesubstitution fixation. Therefore, it would be interesting to detect 5′-NT activity in freeze-substituted photoreceptor cells. This article presents the ultrastructural localization of 5′-NT in rat photoreceptor cells and pigment epithelial cells prepared by rapid-freezing enzyme cytochemistry. The results of this study show that cytochemical detection of enzyme activity and preservation of cell ultrastructure are affected by fixation procedures and, consequently, rapid-freezing enzyme cytochemistry should be a suitable method in retinal cytochemistry. In addition, a striking difference in functional 5′-NTs between rod cells and pigment epithelial cells was also found.
Materials and Methods
Reagents
The following chemicals were purchased from Sigma (St Louis, MO): adenosine 5′-monophosphate (5′-AMP), guanosine 5′-monophosphate (5′-GMP), levamisole, and adenosine 5′-[α,β-methylene]diphosphate (AOPCP). Glutaraldehyde (25% and 70% aqueous) and lead nitrate were obtained from Nacalai Tesque (Kyoto, Japan). All other chemicals were of the highest grade available.
Rapid-freezing Enzyme Cytochemistry
Enzyme cytochemical studies of 5′-NT in the retina were performed primarily on Sprague-Dawley rats. Additional experiments performed on the frog (Xenopus laevis) retina gave basically similar results. This report therefore focuses on results obtained in the rat.
Thirty-six adult male Sprague-Dawley rats (200–300 g) were used. The eye of the rat was removed under sodium pertobarbital anesthesia and then dissected along the equator in Hanks' solution. Subsequently, the neural retina, which is composed of the layers from the rod outer segment to the optic nerve fibers, was gently separated from the pigment epithelium. Within a minute after removal from the animal, the neural retina or the pigment epithelium was rapidly frozen on a liquid nitrogen-cooled copper block in a Polaron E7200 Slammer quick-freezing apparatus. After rapid-freezing, the frozen samples were freeze-substituted as described previously (Saito and Takizawa 1991). Briefly, the samples were freeze-substituted in acetone containing 2% glutaraldehyde or in a mixture of 2% glutaraldehyde and 6% tannic acid for 3 days at −80C. The samples were then processed at −20C for 1 hr and at 4C for 1–12 hr and subsequently transferred to acetone solution without fixatives.
After freeze-substitution, the samples were rehydrated by gradually adding 0.1 M cacodylate buffer, pH 7.2, with 6% sucrose and then rinsed three times in the same buffer. Forty-μm sections were made with a Dosaka EM DTK-3000 Microslicer (Osaka, Japan) or a Komatsu Electronics MB201 freezing microtome (Tokyo, Japan). The sections were then incubated in a lead-containing reaction medium for detection of 5′-NT activity, a modification of the procedure of Wachstein and Meisel (1957), for 30–60 min at 37C with constant agitation. The 5′-NT medium contained 1.25 mM substrate (5′-AMP or 5′-GMP), 80 mM Tris-maleate (pH 7.2), 200 mM sucrose, 3.6 mM lead nitrate, 10 mM magnesium sulfate, and 2 mM levamisole. Control cytochemical incubation consisted of omission of substrate from the reaction mixture and inclusion of AOPCP, a competitive inhibitor, in the complete reaction mixture. After incubation, the sections were postfixed in Caulfield's solution (pH 7.4) containing 1% osmium tetroxide, 56 mM veronal-acetate, and 150 mM sucrose for 1 hr at 4C, dehydrated in a graded series of ethanol, and embedded in Epon.
Conventional Enzyme Cytochemistry
To compare the effect of freeze-substitution fixation on localization of 5′-NT with that of conventional chemical fixation, immersion fixation was carried out as a parallel experiment. Some samples were fixed in 2% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.4) with 6% sucrose for 30 min at 4C before the cytochemical reaction. In this case, the medium and the incubation parameters (time, temperature, mixing conditions) were identical to those mentioned above.
Results
In the samples processed by immersion fixation, 5′-NT activity was detected both in the rod outer segment of the photoreceptor cells and on the apical processes (microvilli and other protrusions) of the pigment epithelial cells (Figures 1C and 1G). The reaction product of 5′-NT activity was localized in the intradiscal space of the vacuolated discs of the rod cells (Figure 1C) and on the outer surface of the apical processes of the pigment epithelial cells (Figure 1G). Thus, the reaction product indicating 5′-NT activity was associated primarily with the exoplasmic side of the cell membranes of both cells.
In the samples processed by rapid-freezing and subsequent freeze-substitution fixation, cell ultrastructure, especially the rod outer segment, was well preserved. Many discs were compactly packed and uniformly flattened (Figures 1A and 1B). It is noteworthy that the intradiscal space was hardly recognizable in the freeze-substituted samples (Figure 1B), whereas it was easily identified in the conventionally fixed retina (Figure 1C). These observations are consistent with earlier morphological findings suggesting that an intact disc is flat (Yamada 1982). The staining reaction for 5′- NT activity was positive in the retinal cells processed by freeze-substitution fixation as well as by immersion fixation (Figures 1A, 1B, 1E, and 1F). However, in the rod outer segment the subcellular localization of the reaction product on the freeze-substituted disc was distinct from that on the conventionally fixed disc. The former was present on the cytoplasmic side of the disc, whereas in the latter it was located on the exoplasmic side (Figures 1B and 1C). On the other hand, in pigment epithelial cells, the ultrastructural localization of 5′-NT was not affected by fixation procedures (i.e., freeze-substitution fixation and conventional immersion fixation). The activity was associated primarily with the exoplasmic surface of the apical process (Figures 1F and 1G). Addition of AOPCP (2 mM) to the incubation medium completely abolished the staining reaction for 5′-NT (Figures 1D and 1H). Omission of substrate resulted in cells entirely lacking reaction product (data not shown).
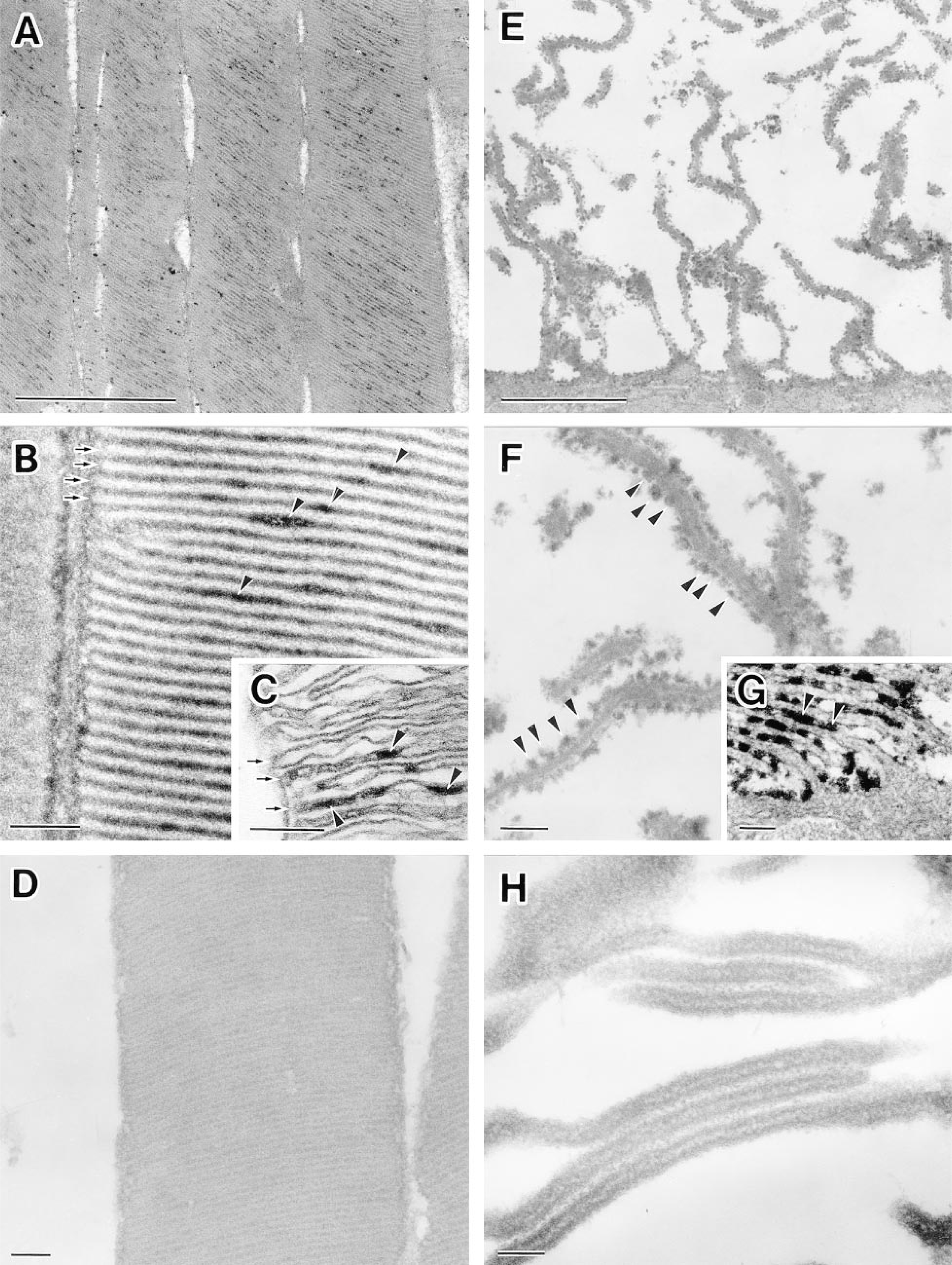
Ultrastructural localization of 5′-nucleotidase (5′-NT) in rat rod outer segments and pigment epithelial cells. (
Discussion
5′-NT is widely distributed in a number of cells and tissues. Animal 5′-NT has been classified into two major forms according to its biochemical properties: a membrane-bound form and a soluble form (for review see Zimmermann 1992). In the outer segment of vertebral photoreceptor cells, the unique properties of 5′-NT have been investigated biochemically. Both membrane-bound and soluble 5′-NTs are present (Fukui and Shichi 1981,1982; Yu and Fager 1983). From biochemical studies, the catalytic site of the soluble 5′-NT is on the extradiscal side of the disk membrane whereas that of the membrane-bound enzyme is on the intradiscal side (Fukui and Shichi 1981,1982; Yu and Fager 1983). The soluble form is believed to be of particular functional importance for the phototransduction system of living photoreceptor cells, whereas it accounts for about 25% of the total 5′-NT activity (Fukui and Shichi 1982). On the other hand, the membrane-bound 5′-NT is considered a nonfunctional enzyme in situ (Yu and Fager 1983). Although the biochemical topology of 5′-NT in the outer segment has been revealed, the cytochemical topology of 5′-NT is controversial. Kreutzberg and Hussain (1984) showed in rat photoreceptor cells that 5′-NT was localized on the cytoplasmic side of discs. On the other hand, Hussain and Baydoun (1985) reported the enzyme in frog photoreceptor cells to be cytochemically undetectable. Saito (1991) noted that 5′-NT activity was demonstrated in the intradiscal space of discs and on the plasma membrane of rat rod cells. These previous studies were carried out with conventional fixation which is often associated with morphological 'odifications, such as a sac-like disc. However, no work has been done on 5′-NT activity by rapid-freezing enzyme cytochemistry, which is one of the most reliable cytochemical techniques for preserving both enzyme activity and cell ultrastructure under more life-like conditions. In this study, 5′-NT in the outer segment of rod cells and in the apical portion of pigment epithelial cells was examined by rapid-freezing enzyme cytochemistry.
A striking difference in the subcellular localization of 5′-NT between the rod outer segment processed by freeze-substitution fixation and that fixed conventionally by immersion is shown in this study (see Figures 1B and 1C). The reason could be that detection of 5′-NT activity and morphological alterations are affected by conventional fixation. The results showing that 5′-NT activity was present on the cytoplasmic side of the freeze-substituted disc membrane and not on the exoplasmic side can be explained by assuming that the cytochemically visible 5′-NT activity represents more functional activity of the soluble 5′-NT. In addition, the membrane-bound 5′-NT activity was detected not in the discs but on the apical processes of pigment epithelial cells (see Figures 1B and 1F). The membrane-bound 5′-NT activity in discs may be inactive in situ and different from that on the apical processes of pigment epithelial cells because each disc membrane is not connected directly to the plasma membrane and the lumen of the disc was almost obliterated (Yamada 1982). The results are in good agreement with the biochemical analyses mentioned above and our previous observations that the localization of other cGMP-related enzymes (i.e., guanylate cyclase and cGMP phosphodiesterase) for the phototransduction system is in the extradiscal space (Saito and Takizawa 1991; Takizawa and Saito 1992). The question of cytochemical heterogeneity of 5′-NT in the other portions of photoreceptor cells remains to be answered (Kreutzberg and Hussain 1984). The asymmetric distribution of 5′-NT on the plasma membrane of retinal Müller cells is also an issue of retinal cytochemistry (Kreutzberg and Hussain 1982). Unfortunately, thus far only the outer segment layer and the pigment epithelium can be rapid-frozen by the metal contact method without discernible ice crystal damages. Moor and co-workers (1980) reported that rapidfreezing under high pressure was able to freeze nerve tissues at a thickness of up to 500 μm without ice crystal damage. It may be possible to answer these questions by a combination of the high-pressure freezing technique and enzyme cytochemistry.
This study presents the localization of 5′-NT activity in the outer segment of the rod cells and in the apical portion of the pigment epithelial cells processed by rapid-freezing enzyme cytochemistry. In the rod outer segments, the soluble 5′-NT on the cytoplasmic side of the disk membrane was vital, whereas in the pigment epithelial cells the membrane-bound ecto-5′-NT on the exoplasmic (external) surface of the apical process was active. Rapid-freezing enzyme cytochemistry is a recommended approach for detection of other enzymes associated with subcellular structures that are not well preserved by conventional fixation, and can provide information about cells approximating, as closely as possible, the conditions in living cells.
Footnotes
Acknowledgements
Supported in part by grants from the Ministry of Education, Science, Sports, and Culture of Japan.
I am grateful to Dr Takuma Saito of Jichi Medical School for critical comments on this manuscript. I wish to thank Ms Kiyomi Inose, Ms Chiaki Ishijima, and Ms Megumi Yatabe for excellent technical assistance.