
Abstract
Ion-transporting Na,K-ATPase plays an essential role in nerve conduction. To clarify the cytochemical effects of reserpine on transport Na,K-ATPase activity, the localization of ouabain-sensitive, K+-dependent p-nitrophenylphosphatase (K-NPPase) activity was investigated in the facial nerves of normal and reserpinized guinea pigs using a cerium-based method. In the normal facial nerve, the reaction product of K-NPPase activity was observed on the internodal axolemma and Schmidt-Lanterman incisures. In the Ranvier nodes, enzyme activity was localized to the paranodal and nodal axolemma. In the reserpinized nerves, reaction product was detectable on the nodal axolemma but was undetectable on the other parts of the axolemma. Nodal K-NPPase was not affected by reserpine treatment. Therefore, the transport Na,K-ATPase on the nodal axolemma might differ from that on the other parts of the axolemma. Allowing reserpinized animals to survive. Two different ouabain-sensitive K-NPPase reactivities, “reserpine-sensitive” and “reserpine-resistant,” might be present in the facial nerve of guinea pigs.
Keywords
Materials and Methods
The intratemporal portion of bilateral facial nerves was obtained from 13 Hartley guinea pigs weighing 400–500 g with a normal Prayer's reflex. The experimental animals were divided into the following three groups. The first group was sacrificed at 3 days after one-shot reserpine (10 mg/kg
Preparation of Tissues
Under deep ketamine hydrochloride (
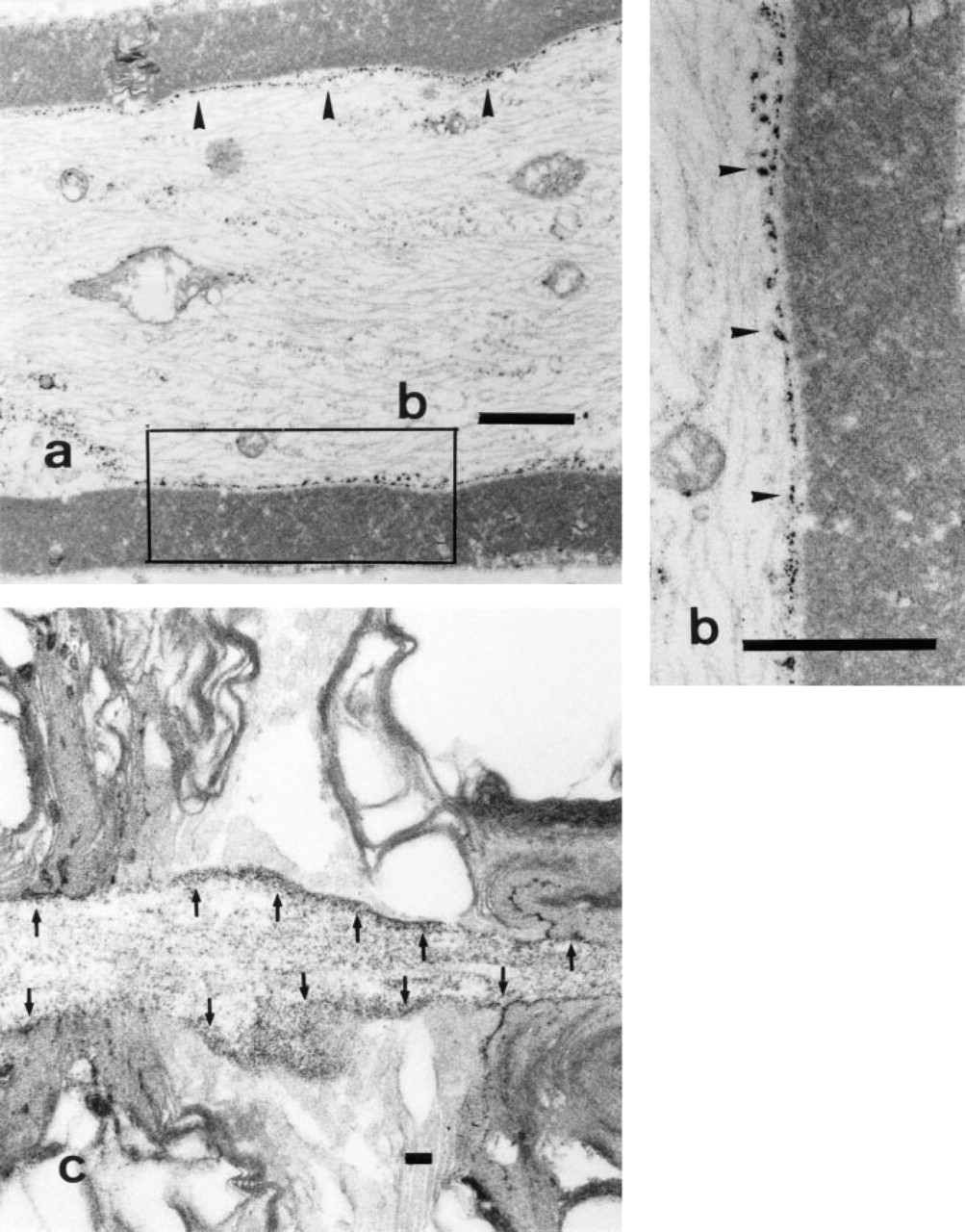
Normal facial nerve.
Cytochemical Procedure
The tissues were cut into sections 40 μm thick with a Mi-croslicer (Dosaka EM; Kyoto, Japan). The tissue samples were rinsed with 50 mM Tricine buffer, pH 7.5, for 15 min and incubated in the medium according to Kobayashi et al. (1987). The incubation medium contained 50 mM Tricine buffer, pH 7.5, 2 mM cerium chloride as the capture agent, 10 mM magnesium chloride, 50 mM potassium chloride, and 2 mM p-NPP (Mg salt) as the substrate, 5% sucrose, 2.5 mM levamisole (an inhibitor of alkaline phosphatase), and 0.00015% Triton X-100 to facilitate penetration of the incubation medium (Robinson 1985). The medium was filtered through 0.22-μm pore size filters after the pH was adjusted to 7.5. Incubation was performed for 45 min at 37C in a shaking incubator. Tissue samples were washed for 10 min with 0.1 mM Tris-maleate buffer at pH 6.0, then processed for electron microscopy. In control experiments to determine negative enzyme reactivity, tissue samples were incubated in a substrate-free medium, in a medium containing 10 mM ouabain, and in a medium in which K+ had been replaced with Na+.
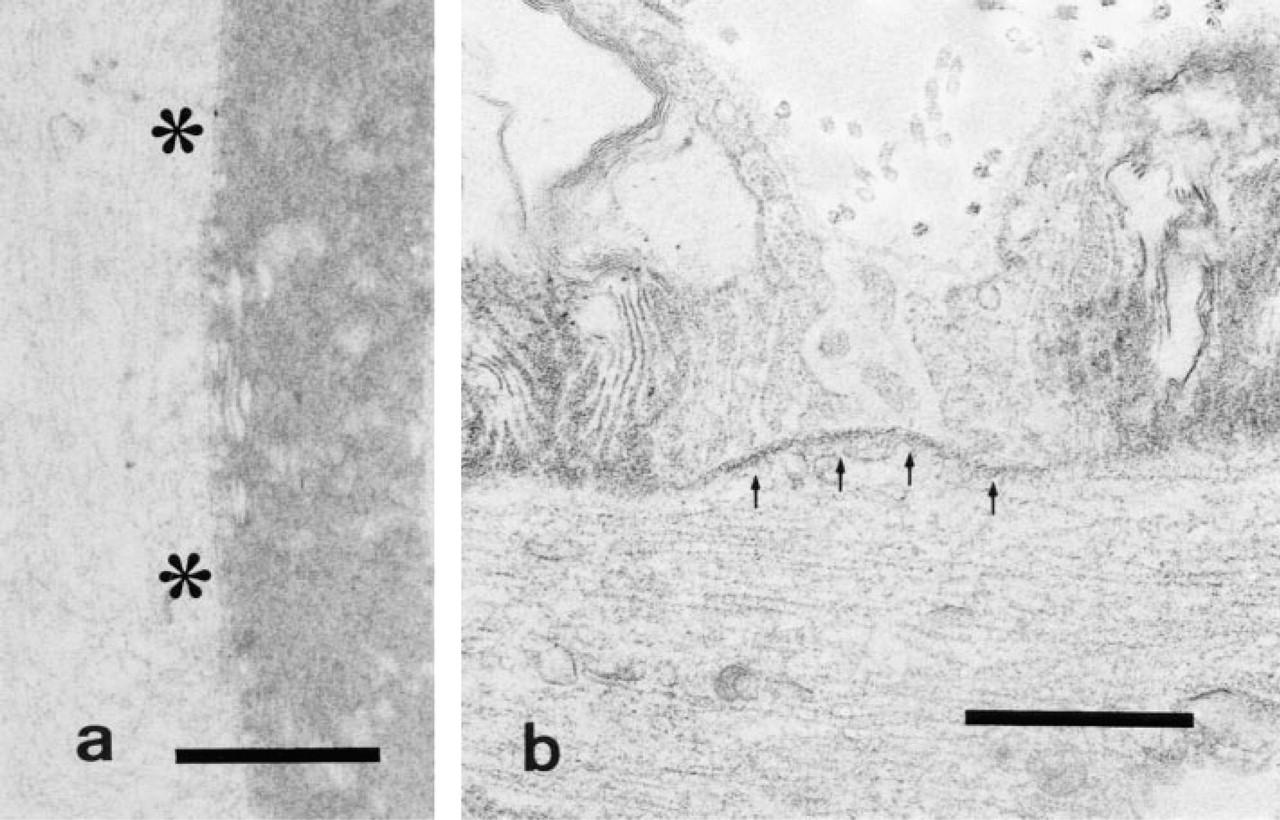
Reserpinized facial nerve.
Procedure for Electron Microscopy
After cytochemical incubation, the tissues were postfixed with 2% OsO4 in 0.2 M cacodylate buffer, pH 7.4, for 1 hr at room temperature. The specimens were then dehydrated through a graded series of alcohol solutions and propylene oxide, and then embedded in Spurr's epoxy resin (Spurr 1969). Thin sections were cut on an Ultracut OmU4 ultrami-crotome (C. Reichert; Vienna, Austria) and picked up with copper grids. Uncontrasted ultrathin sections were then observed under a JEM-1200 electron microscope (JEOL; Tokyo, Japan).
The animal use protocol was approved by the Institutional Animal Care and Use Committee of the Hyogo College of Medicine, and all experiments were performed in accordance with the guidelines of the Declaration of Helsinki.
Results
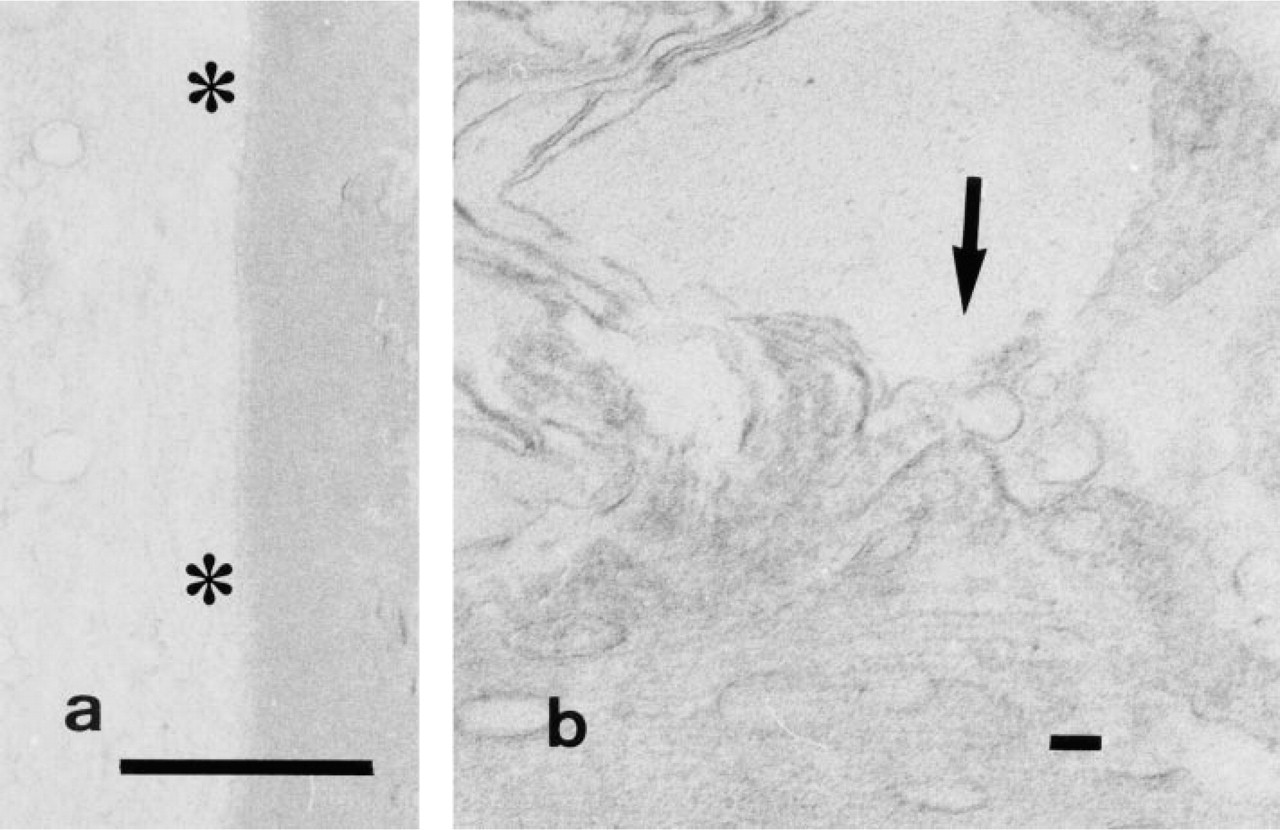
In the axon, cytochemically fine granular reaction product was clearly deposited on the cytoplasmic side of the entire axolemma but was hardly visible on the microtubules in the axoplasm (Figures 1a and 1b). In the nodes of Ranvier, the reaction product was localized to the nodal, paranodal, and internodal portions of the axolemma. Enzyme activity was also localized to the cytoplasmic side of the plasma membrane in the terminal paranodal loop of Schwann cells (Figure 1c).
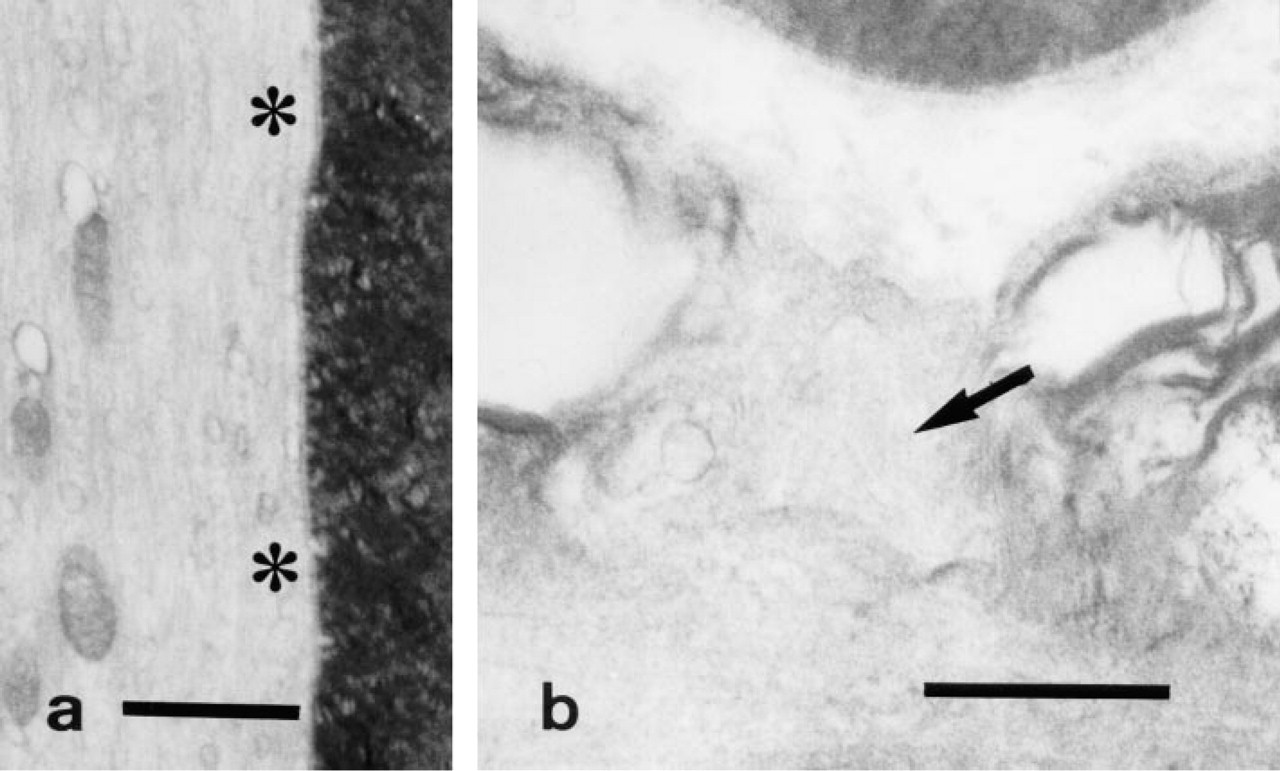
Control study (ouabain treatment).
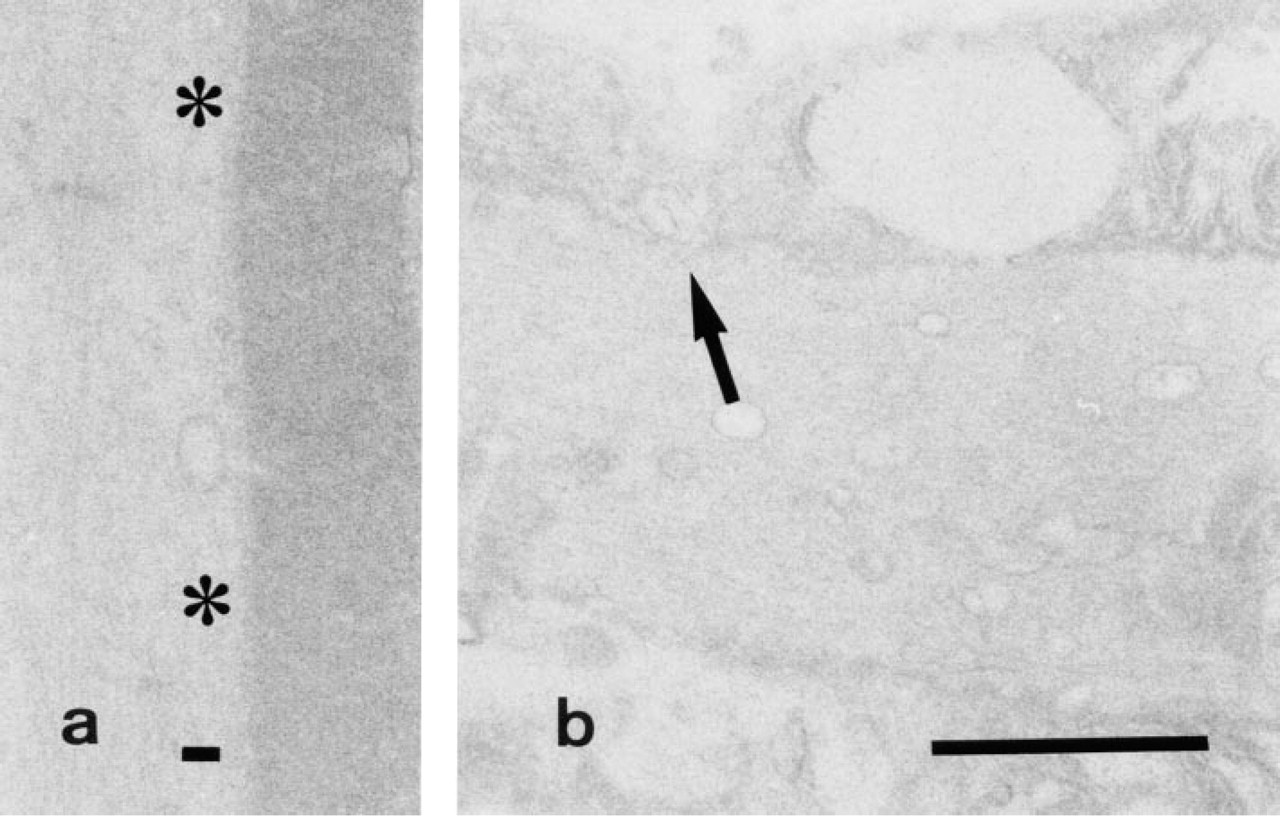
Control study (substrate-free).
In animals sacrificed at 3 days after one-shot reserpine administration, the enzyme reaction was undetectable on the internodal, paranodal axolemma, and Schmidt-Lanterman incisure (Figure 2a), whereas it was detectable on the Ranvier node axolemma (Figure 2b).
In the control samples, formation of reaction product was almost completely inhibited when 10 mM ouabain was included in the medium (Figures 3a and 3b) and reaction product was undetectable in a substrate-free medium (Figures 4a and 4b) and in a medium in which K+ had been replaced with Na+ (Figures 5a and 5b).
Control study (K+ replacement with Na+).
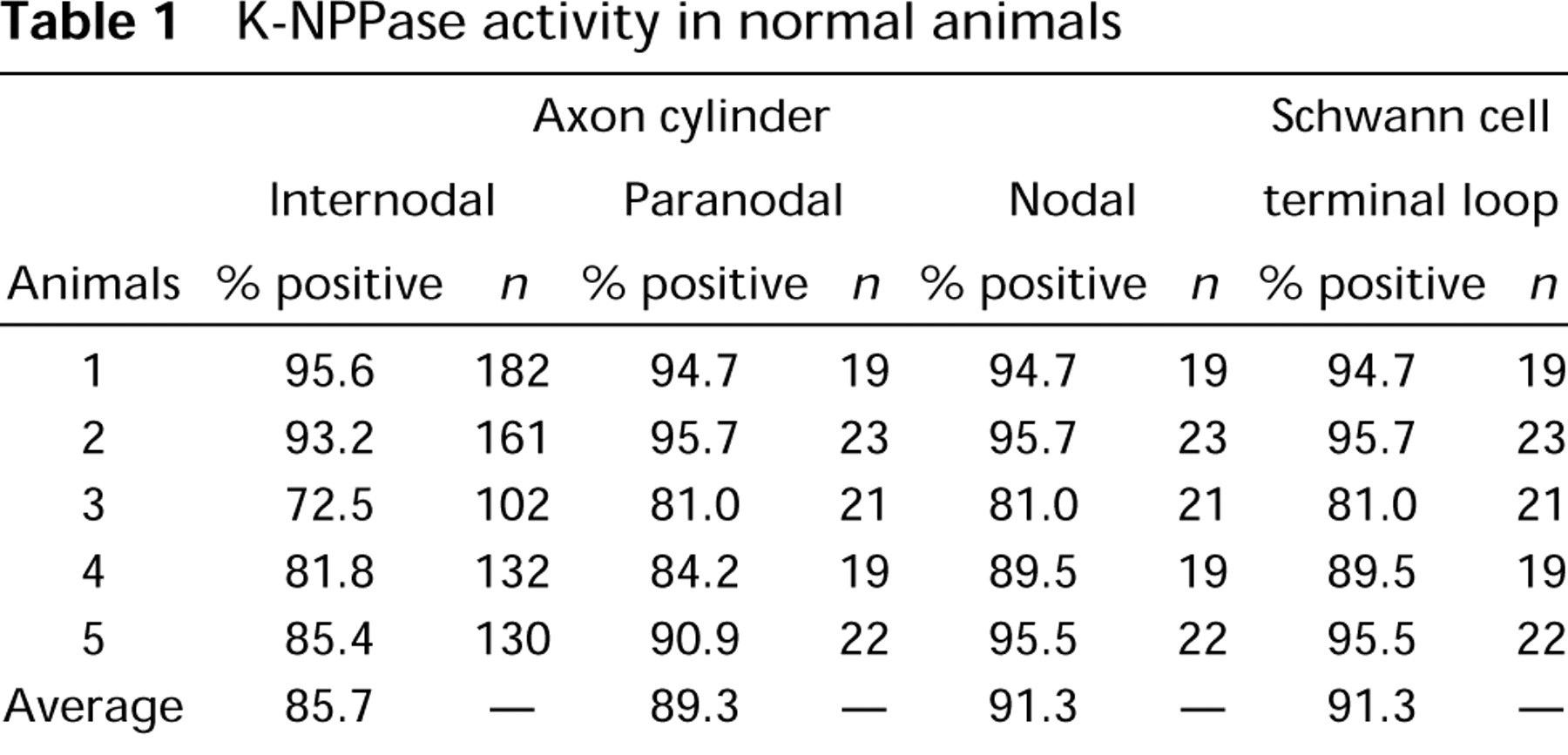
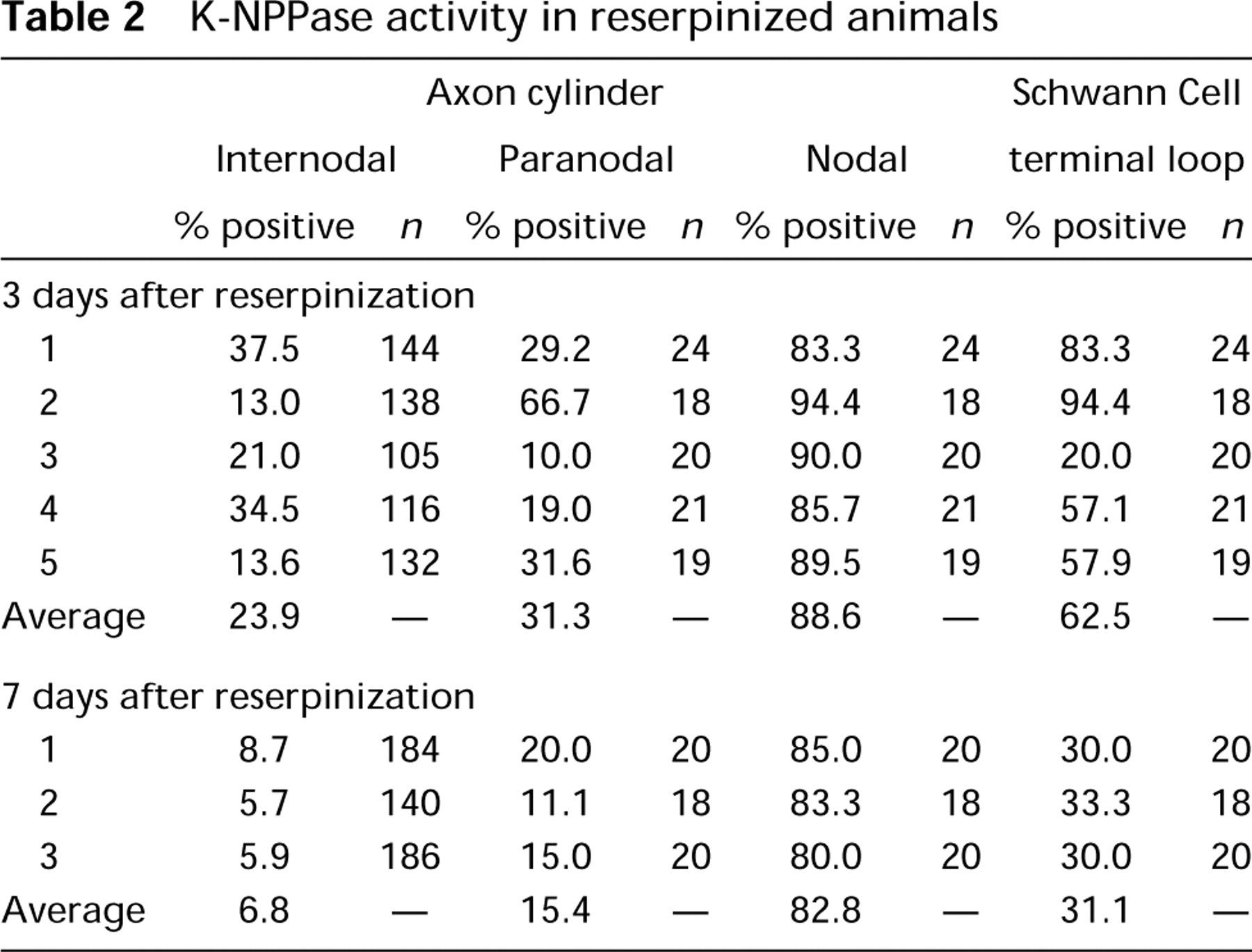
Tables 1 and 2 summarize the results obtained from the five normal and eight reserpinized guinea pigs, respectively. In untreated normal animals, K-NPPase activity was detectable in 617 internodal axolemmae of a total of 707 (87.2%) nerve fibers and was also noted in 95 nodal axolemmae of 104 (91.3%) nodes of Ran-vier. However, in the 3 days after reserpinization, enzyme activity was only detectable in 152 internodal axolemmae of 635 (23.9%) but was clearly detectable in 90 nodal axolemmae of 102 (88.2%) investigated cases. At 7 days after reserpinization, enzyme activity was detectable only in 35 internodal axolemmae of 510 (6.9%) but was clearly detected in 48 nodal axolemmae of the 58 (82.8%) cases investigated. Reaction product was detectable on the nodal axolemma in the facial nerve but was almost undetectable on other parts of the axolemma after reserpine treatment.
K-NPPase activity in normal animals
Discussion
Ion-transporting ATPase is a membrane-bound enzyme that couples ATP hydrolysis to the active transport of sodium and potassium ions and plays a crucial role in nerve conduction (Albers et al. 1989). In the present study, the cerium-based method of Kobayashi et al. (1987) was employed to detect K-NPPase activity. The major advantages of this method are its ability to detect ouabain-sensitive K-NPPase activity at physiological pH and the absence of nonspecific deposits and reaction product diffusion.
In previous cytochemical studies of the nervous system, Na,K-ATPase activity was reported to be localized on the synaptic plasma membrane (Vorbrodt et al. 1982; Inomata et al. 1983; Nasu 1983), the axolemma of axons and dendrites (Stahl and Broderson 1976; Broderson et al. 1978; Vorbrodt et al. 1982; Inomata et al. 1983; Nasu 1983), and the astrocytic foot processes (Mrsulja et al. 1985) in the brain, and on the nodal axolemmae, and the paranodal regions of Schwann cells in peripheral nerves, including the sciatic nerve (Vorbrodt et al. 1982; Mrsulja et al. 1985; Ariyasu and Ellisman 1989). Therefore, there appears to be no significant difference in the localization of Na,K-ATPase activity between the facial nerve (Kanoh et al. 1993b,1994; Kanoh and Kumoi 1994) and other peripheral nerves, such as the sciatic nerve.
Na,K-ATPase contains two polypeptide subunits: the α or catalytic subunit and the β or glycoprotein subunit. In 1979, Sweadner described two forms of the catalytic unit of Na,K-ATPase, the α type, which has a low affinity for ouabain, and the α+ type, which has a high affinity for it. The enzyme detected in the current experimental study was almost completely inhibited by ouabain, and therefore appeared to be the α + form of Na,K-ATPase. Recently, three α-isoforms, α1, α2, and α3, have been sequenced in the rat brain (Shull et al. 1986; Herrera et al. 1987; Orlowski and Lingrel 1988), and three α-polypeptides have also been detected immunohistochemically in the brain (Arystarkhova et al. 1989; Shyjan and Levenson 1989; Urayama et al. 1989). It is now known that α1 is the original α-isoform, and that α2 and α3 are contained in the α+ isoform (Schneider et al. 1988).
Reserpine, an adrenergic neuron blocker, belongs to the family rauwolfia alkaloids and is employed clinically for treatment of hypertension. Pharmacologically, this compound releases biological amines such as norepinephrine, epinephrine, dopamine, and serotonin from storage or binding sites in the central and peripheral nervous system. Therefore, high doses of reserpine induce depletion of these amines, inhibiting reabsorption in the storage site and preventing recombination at the binding sites. The purpose of the present study was to evaluate the cytochemical effects of catecholamine depletion on transport Na,K-ATPase activity in the guinea pig facial nerve. The dose of reserpine used in the present study (10 mg/kg) was believed to be high enough to completely abolish the activity of catecholamines (Wakada 1980). This dose is 500–1000 times higher than that in clinical use. The biological half-lives of reserpine in serum and blood were also reported to be 271 hr and 386 hr, respectively (Maass et al. 1969). Therefore, for evaluating the effects of reserpine on K-NPPase activity, Day 3 and Day 7 after reserpine administration represent the reasonable checkpoints.
K-NPPase activity in reserpinized animals
In the cochlea, K-NPPase activity in the stria vascularis was shown to be almost completely decreased after reserpine administration (Kanoh et al. 1993a, 1994). In a paper to be published in the near future, K-NPPase activity in the kidney was reported to be similarly suppressed after reserpine administration. In the present study, reaction product was detectable on the nodal axolemma but was almost undetectable on other parts of the axolemma and Schmidt-Lanterman incisures after reserpine treatment, i.e., transport Na,K-ATPase localized to the Ranvier nodes was not affected by reserpine administration, whereas transport Na,K-ATPase detected on other parts of the axolemma was decreased by reserpine treatment. Therefore, two different ouabain-sensitive K-NPPase reactivities, “re-serpine-sensitive” and “reserpine-resistant,” might be present in the facial nerve. Even when transport Na,K-ATPase activity on the internodal axolemma was almost completely eliminated, reserpinized animals displayed no facial palsy (Kanoh and Sakagami 1996), and could survive. In 1991, Mata et al. reported two different immunoreactivities in the nervous system. They evaluated immunocytochemical localization in the sciatic and optic nerves of the rat using a polyclonal antiserum raised against the denatured catalytic subunit of brain Na+,K+-ATPase. Immunoreactivity was detected along the internodal axolemma of myelinated fibers in both nerves and was undetectable on the nodal axolemma. Although the mechanism of suppression of K-NPPase activity in the facial nerves requires further consideration, the current results suggest that the nodal and internodal axolemma may contain different enzymes and that, because reserpine induces the depletion of sympathetic substances, transport Na,K-ATPase may be partially maintained by catecholamines. Other explanations at the molecular level will be forthcoming in the near future.
Footnotes
Acknowledgements
Supported by a Grant-in-Aid (08671998) for Science Research from the Ministry of Education, Science and Culture of Japan.
The author thanks Prof Harumichi Seguchi and Associate Prof Teruhiko Okada, Department of Anatomy and Cell Biology, Kochi Medical School, for their valuable comments.