
Abstract
Enzyme activity that represents ouabain-insensitive, potassium-dependent p-nitrophenylphosphatase (p-NPPase) was assessed in rat atrial myocytes by biochemical and cytochemical procedures. No activity was detected in parallel experiments with ventricular myocytes. Fixed tissues were incubated in a reaction medium containing Tricine buffer, p-nitrophenylphosphate (p-NPP), KCl, MgCl2, CaCl2, CeCl3, Triton X-100, levamisole, and ouabain. Final pH was adjusted to 7.5. Biochemical studies showed that accumulation of p-nitrophenol in the medium was increased proportionally in accordance with the amount of incubated tissue. This activity was optimal with incubation at pH 7.5 and in the presence of KCl. Approximately 70% of the enzyme was inhibited by 2 mM CeCl3. Electron microscopic observations revealed reaction product (RP) at sites of ouabain-insensitive, potassium-dependent p-NPPase activity as electron-dense precipitate localized at the inner surface of the plasma membrane and at the T-tubules of atrial myocytes. Control experiments indicated that the activity was strongly inhibited by sodium orthovanadate and was repressed by omeprazole and 1,3-dicyclohexylcarbodiimide. X-ray microanalysis confirmed the presence of cerium within the cytochemical RP. The ouabain-insensitive, K-dependent p-NPPase activity detected in the present study is considered to be an isoform of a P-type, H-transporting, K-dependent adenosine triphosphatase (H,K-ATPase).
Keywords
A
Renewed interest in phosphatase transporting systems has recently arisen from the invention of cerium-based cytochemical methods. These methods provide a new aspect into the ATPase cytochemistry because they are highly reproducible for enzyme demonstration, generate very small primary cerium reaction product precipitates, and are highly sensitive for enzyme determinations (Halbhuber et al. 1994). The cerium-based methodology has been also employed for both Na,K-ATPase and H,K-ATPase (Kobayashi and Seguchi 1990; Kobayashi et al. 1987).
H,K-ATPase is identified in many tissues and cells (Wingo and Smolka 1995; Miyabo-Takeguchi 1994; Takeguchi et al. 1990; Wingo et al. 1990; Fain et al. 1988). However, very little is known about this enzyme in atrial myocytes. As has been previously reported cytochemically, myocytes from the right atrium exhibit K-dependent p-NPPase activity that is resistant to ouabain (Zinchuk 1992). To elucidate this activity more precisely, we undertook a study employing biochemical and cerium-based cytochemical methodologies. Different controls were utilized to exclude possible interference from other transporting ATPases. Our observations have led to the assumption that the specific function of atrial myocytes forms a basis for the existence of the enzymatic system that is closely related to transporting H,K-ATPase and may be responsible for coordination of secretory and contraction mechanisms.
Materials and Methods
Chemicals
Tricine, p-nitrophenylphosphate (p-NPP) (Mg salt), levamisole, furosemide, and Triton X-100 were purchased from Sigma Chemical (St Louis, MO). 1,3-Dicyclohexylcarbodiimide (DCCD) was from Aldrich Chemical (Milwaukee, WI). Cerium chloride and ouabain were from Nacalai Tesque (Kyoto, Japan). The other chemicals were purchased from Wako Pure Chemical Industries (Osaka, Japan). Omeprazole (5-methoxy-2-(((4-methoxy-3,5,-dimethyl-2-pyridinyl)-methyl)-sulfinyl)-1H-benzimidazole) from Hässle's Chemistry Department (Mölndal, Sweden) was a donation from Fujisawa Astra (Osaka, Japan). All reagents were of the highest grade of purity available.
Animals
Male Sprague–Dawley rats weighing 250–300 g were used. Animals were housed according to standard conditions and had free access to food and water.
Preparation of Tissues. Rats were sacrificed under ether anesthesia. The hearts were quickly removed, and the atria and ventricles were cut into pieces less than 1 mm thick and fixed with a freshly prepared fixative containing 2% paraformaldehyde (PFA) and 0.2% glutaraldehyde (GLA) in 0.1 M cacodylate buffer, pH 7.4, for 40 min at 0-4C. Then the tissues were washed overnight in 0.1 M cacodylate buffer, pH 7.4, cut into 70-90-μm slices with a Microslicer (DTK-2000; Dosaka EM, Kyoto, Japan), and rinsed with 50 mM Tricine buffer, pH 7.5, containing 50 mM KCl and 20 mM MgCl2. Tissue slices from atria and ventricles were examined in parallel biochemical and cytochemical experiments and served as control of the efficiency of Na,K-ATPase inhibition throughout this study. Tissues were preincubated with 10 mM ouabain in a medium containing 50 mM Tricine buffer, pH 7.5, 50 mM KCl, and 20 mM MgCl2, for 20 min at room temperature to inhibit Na,K-ATPase activity. Preincubation with ouabain facilitates its binding to Na,K-ATPase from cardiac tissue (Matsui and Schwartz 1968).
Biochemical Procedure
Fixed tissue slices of 20 mg total wet weight were incubated under different conditions in 10 ml reaction medium containing 50 mM Tricine buffer, pH 7.5, 2 mM p-NPP (Mg salt), 50 mM KCl, 20 mM MgCl2, 10 mM CaCl2, 5 mM levamisole, and 10 mM ouabain. Incubation lasted for 30 min at 37C. After incubation, the specimens were removed and the medium was employed for biochemical analysis. The absorbance of p-nitrophenol produced by hydrolysis of p-NPP was measured at 410 nm and compared with p-nitrophenol standards in a Hitachi 220A spectrophotometer (Hitachi; Tokyo, Japan). The following effects on the enzyme activity were tested spectrophotometrically: (a) concentrations of GLA in the fixative mixture; (b) the amount of incubated tissue; (c) pH of the medium; (d) K concentrations; (e) Na concentrations; (f) cerium concentrations; and (g) inhibitors. To compare the intensity of p-nitrophenol liberation generated by H,K-ATPase to that by Na,K-ATPase, several parallel biochemical experiments were also performed to evaluate Na,K-ATPase activity in atrial and/or ventricular muscle. In those controls, the concentration of GLA in the fixative mixture was 0.05%, no preincubation with ouabain was employed, and ouabain was excluded from the reaction medium.
Different inhibitors were used to test the validity of the enzyme activity demonstrated in this experiment. Ouabain, levamisole, and CaCl2 were added directly to the incubation medium. At neutral pH of the medium, the complex ouabain enzyme is formed more efficiently (Harris et al. 1973). Levamisole was used as inhibitor of nonspecific alkaline phosphatase because this enzyme can cleave p-NPP (Van Belle 1972). Levamisole neither affects substrate p-NPP nor inhibits p-NPPase activity. It is also essential for maximal ouabain inhibition of Na,K-ATPase activity (Stahl and Broderson 1976). Because Ca-ATPase can also hydrolyze p-NPP (Caride et al. 1982), we used 10 mM CaCl2 as a component of the medium to inhibit this enzyme. To prevent turbidity of the medium, sodium orthovanadate was added only to the same preincubation medium as for ouabain control for 20 min. For the same reason, furosemide was added only to the preincubation medium containing 50 mM Tricine buffer, pH 7.5, 100 mM NaCl, and 20 mM MgCl2 for 20 min. DCCD was used in methanol-dissolved form. Omeprazole was used in ethanol-dissolved form. To facilitate penetration of omeprazole into the incubated material, tissues were preincubated in 45 mM Tris-acetate buffer, pH 7.4, containing 1 mM omeprazole and 10% ethanol, for 20 min. Because it reacts with sulfhydryl groups of the enzyme under acidic conditions (Im et al. 1985), we also employed preincubation of the tissue in Tris-acetate buffer, pH 5.0, with the same composition for another 20 min at room temperature before incubation in the reaction medium.
Cytochemical Procedure
Fixed tissue slices were incubated in a reaction medium under the conditions with the biochemically determined optimum described above, with the addition of 2 mM CeCl3 and 0.0002% Triton X-100. The reaction was terminated by washing with 50 mM Tricine buffer, pH 7.5, for 10 min and with 0.1 M cacodylate buffer, pH 7.4, for another 10 min.
As controls, tissues were incubated in the presence of (a) 10 mM sodium orthovanadate, (b) 1 mM omeprazole, (c) 1 mM DCCD, (d) in the absence of CaCl2, or (e) in a substrate-free medium. In addition, the effects of an increase in the concentration of KCl, KCl replacement by NaCl, and the presence of 2 mM furosemide were also examined.
Electron Microscopy
Tissues were postfixed with 0.1 M cacodylate-buffered 1% OsO4, pH 7.4, dehydrated in graded ethanols and propylene oxide, and embedded in Spurr's epoxy resin (Spurr 1969). Ultrathin sections were cut on a diamond knife with a Reichert Ultracut E ultramicrotome (Reichert; Vienna, Austria) and examined unstained or stained with uranyl acetate in a JEM 100 S (JEOL; Tokyo, Japan) at an accelerating voltage of 80 kV.
For X-ray spectroscopic microanalysis, unstained samples were collected either on uncoated copper or on colloidin-coated carbon grids. The product of the cytochemical reaction was analyzed in a JEM 2010 HC (JEOL) equipped with an ISIS energy dispersive spectrum analyzing system (EDS; Oxford Instruments, Oxford, UK) for 100 sec real time at accelerating voltage of 200 kV.
Results
Biochemical Assays
Biochemical experiments showed a linear increase of ouabain-insensitive p-NPPase (H,K-ATPase) activity from fixed (0.2% GLA) and further incubated (reaction medium) atrial tissue slices in proportion to the increase with the amount of incubated tissues (Figure 1). Increase of the concentration of GLA in the fixative mixture from 0.05% to 0.2% inhibited 17% of H,K-ATPase activity (Table 1). In parallel control experiments with ventricular myocytes, the liberation of p-nitrophenol when tissues were fixed with 0.05% GLA was presumably generated by surviving Na,K-ATPase. In response to 0.2% GLA, the amount of p-nitrophenol liberation in ventricular myocytes drastically decreased and constituted only 13% of that detected in atrial myocytes. This residual amount of p-nitrophenol is so negligible (1.4 ± 0.2 nmoles/mg/30 min) that it is unable to form either properties of the biochemical reaction or the cytochemical RP (see below). This shows that H,K-ATPase activity is present only in atrial myocytes, because under the same experimental conditions a considerable substrate hydrolysis (12.7 ± 1.0 nmoles/mg/30 min) occurred only in atrial samples. Because we did not observe differences in Na,K-ATPase activity from atrial and ventricular samples, this indicates that when heart tissue of the rat is treated with the fixative mixture used in this study (containing 0.2% GLA) with further preincubation of the tissue with 10 mM ouabain and incubation in a reaction medium containing 10 mM ouabain, the activity of Na,K-ATPase is eliminated in both atrial and ventricular myocytes and therefore does not influence the results of this study.
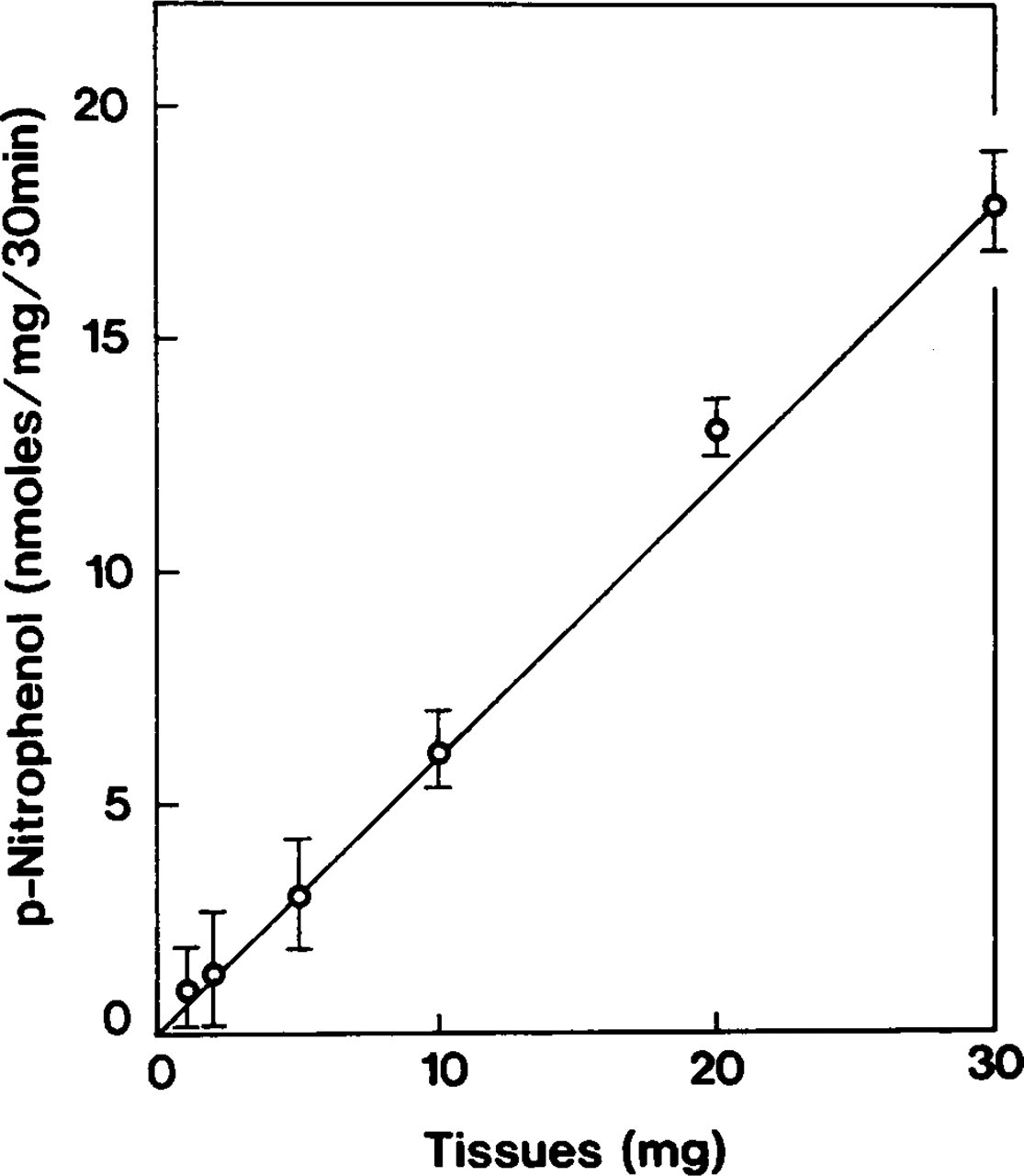
Relation of accumulation of p-nitrophenol to the amount of fixed and incubated atrial tissue slices. Tissues were fixed with a mixture of 2% PFA and 0.2% GLA and further incubated with a reaction medium to study ouabain-insensitive, K-dependent p-NPPase (H,K-ATPase). The accumulation of p-nitrophenol increases in linear proportion with an appropriate increase in incubated tissue. Symbols represent mean ± SE of measurements performed in three separate experiments.
Amount of p-nitrophenol liberation under different fixation conditions with further incubation in the reaction medium a
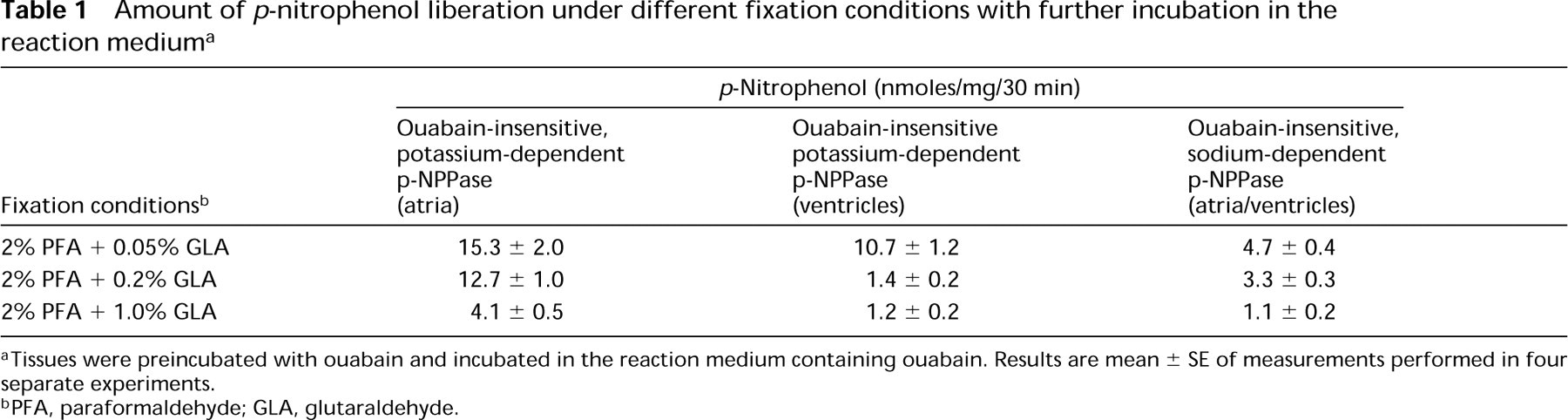
aTissues were preincubated with ouabain and incubated in the reaction medium containing ouabain. Results are mean ± SE of measurements performed in four separate experiments.
bPFA, paraformaldehyde; GLA, glutaraldehyde.
Amount of p-nitrophenol liberation under different incubation conditions a
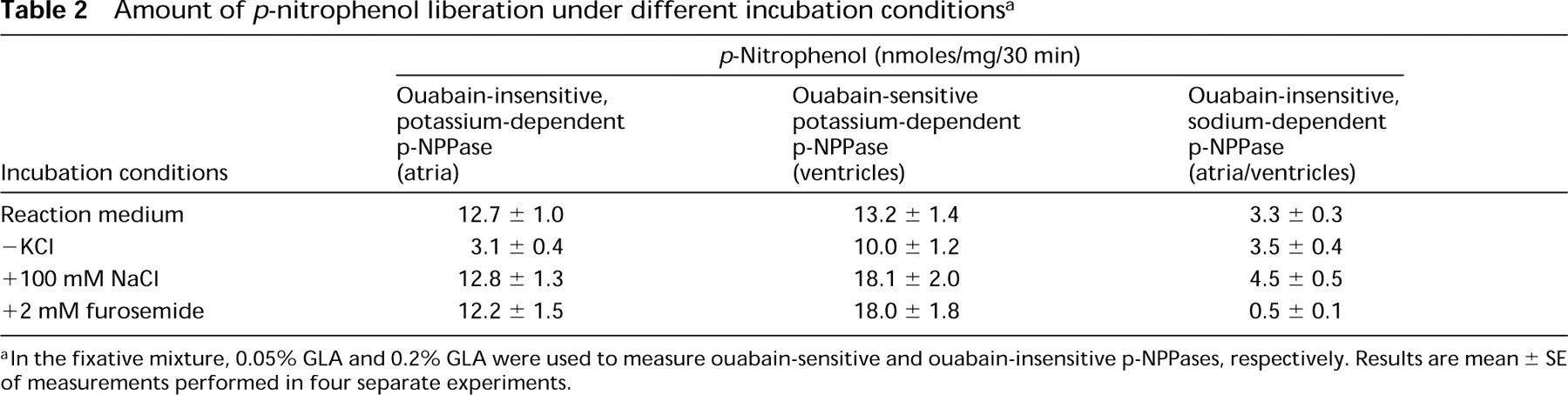
aIn the fixative mixture, 0.05% GLA and 0.2% GLA were used to measure ouabain-sensitive and ouabain-insensitive p-NPPases, respectively. Results are mean ± SE of measurements performed in four separate experiments.
Accumulation of p-nitrophenol was optimal within a pH range of 7.4–7.6 (Figure 2). Ouabain-insensitive p-NPPase activity was dependent on potassium ions (Table 2) and constituted more than half of the detected maximum at a KCl concentration of 10 mM. The enzyme activity increased gradually in the 50–70-mM range and became almost constant thereafter (Figure 3). When tissues were incubated with 100 mM NaCl instead of KCl, the accumulation of p-nitrophenol formed 26% of the total ouabain-insensitive, potassium-dependent p-NPPase (H,K-ATPase) activity (Figure 4). However, this weak ouabain-insensitive, potassium-independent p-NPPase activity was stimulated by an increase in the concentration of sodium ions and revealed strong inhibition when 2 mM furosemide was added (Table 2). Cerium ions inhibited approximately 70% of H,K-ATPase activity. This inhibitory effect was almost constant at concentrations of CeCl3 in the 2–4 mM range (Figure 5).
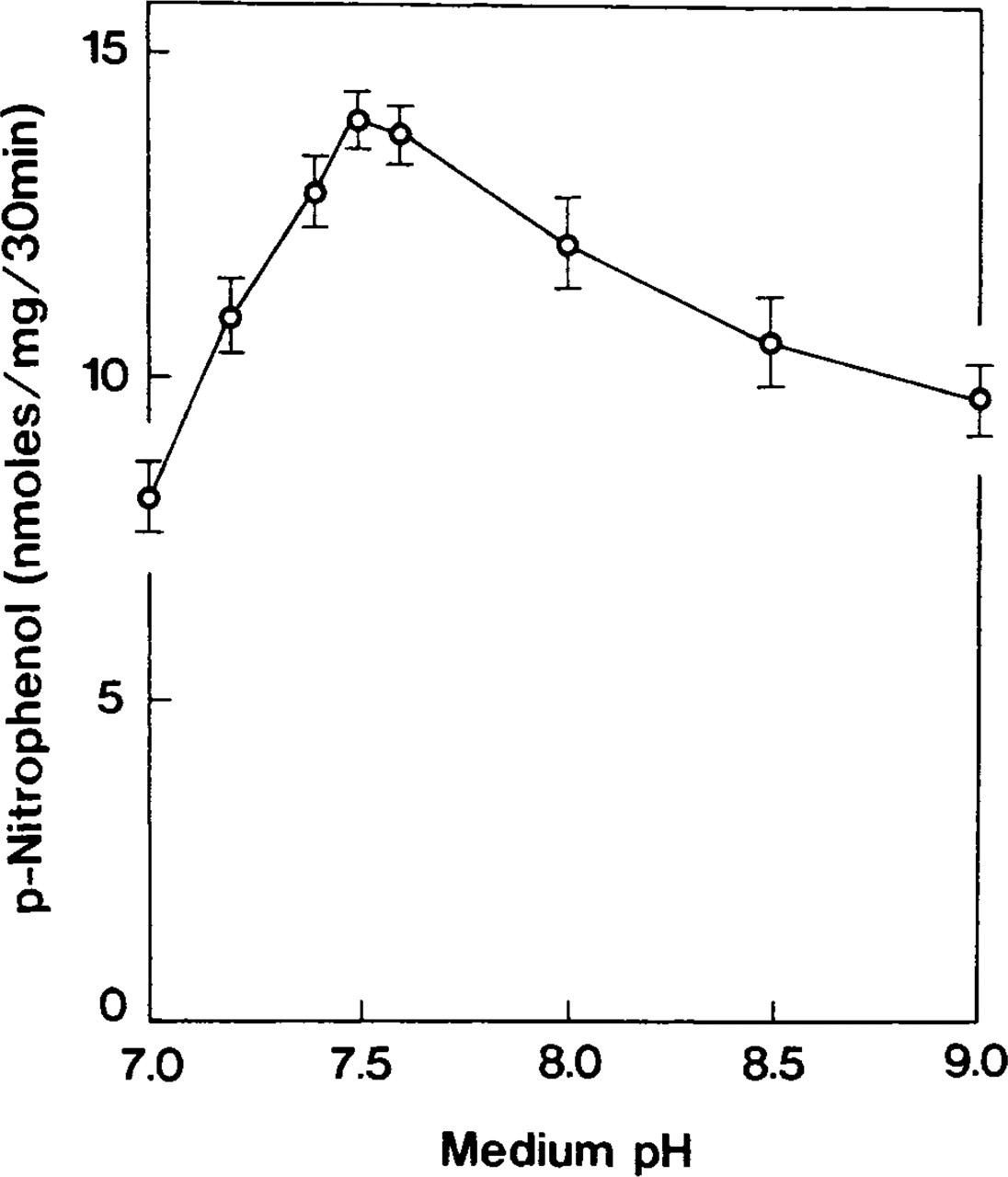
Effect of pH of the incubation medium on ouabain-insensitive p-NPPase activity. Enzyme activity is optimal within neutral pH. Symbols represent the mean ± SE of measurements performed in four separate experiments.
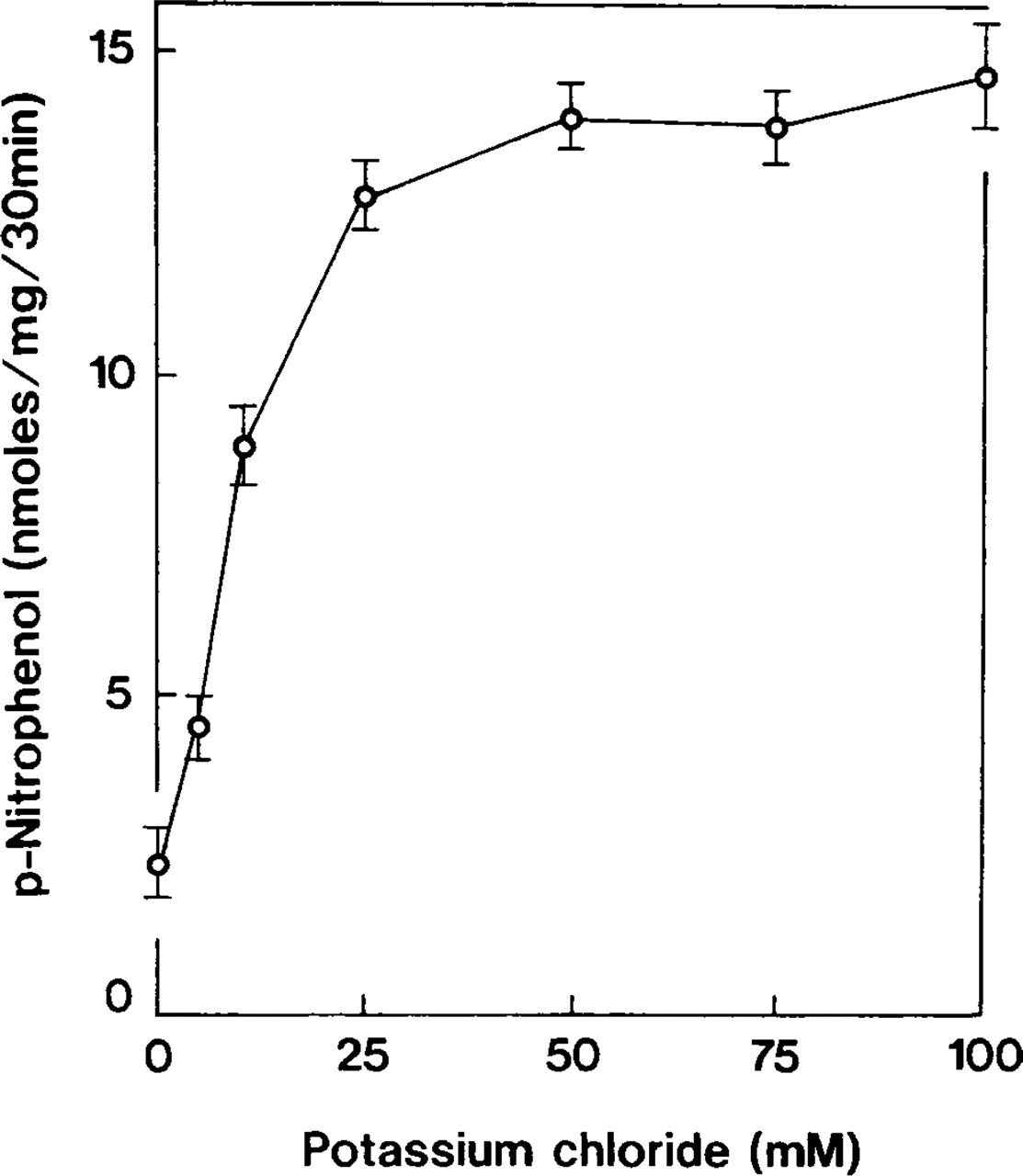
Effect of potassium concentrations on enzyme activity. Ouabain-insensitive p-NPPase is dependent on potassium ions. Symbols represent mean ± SE of measurements performed in four separate experiments.
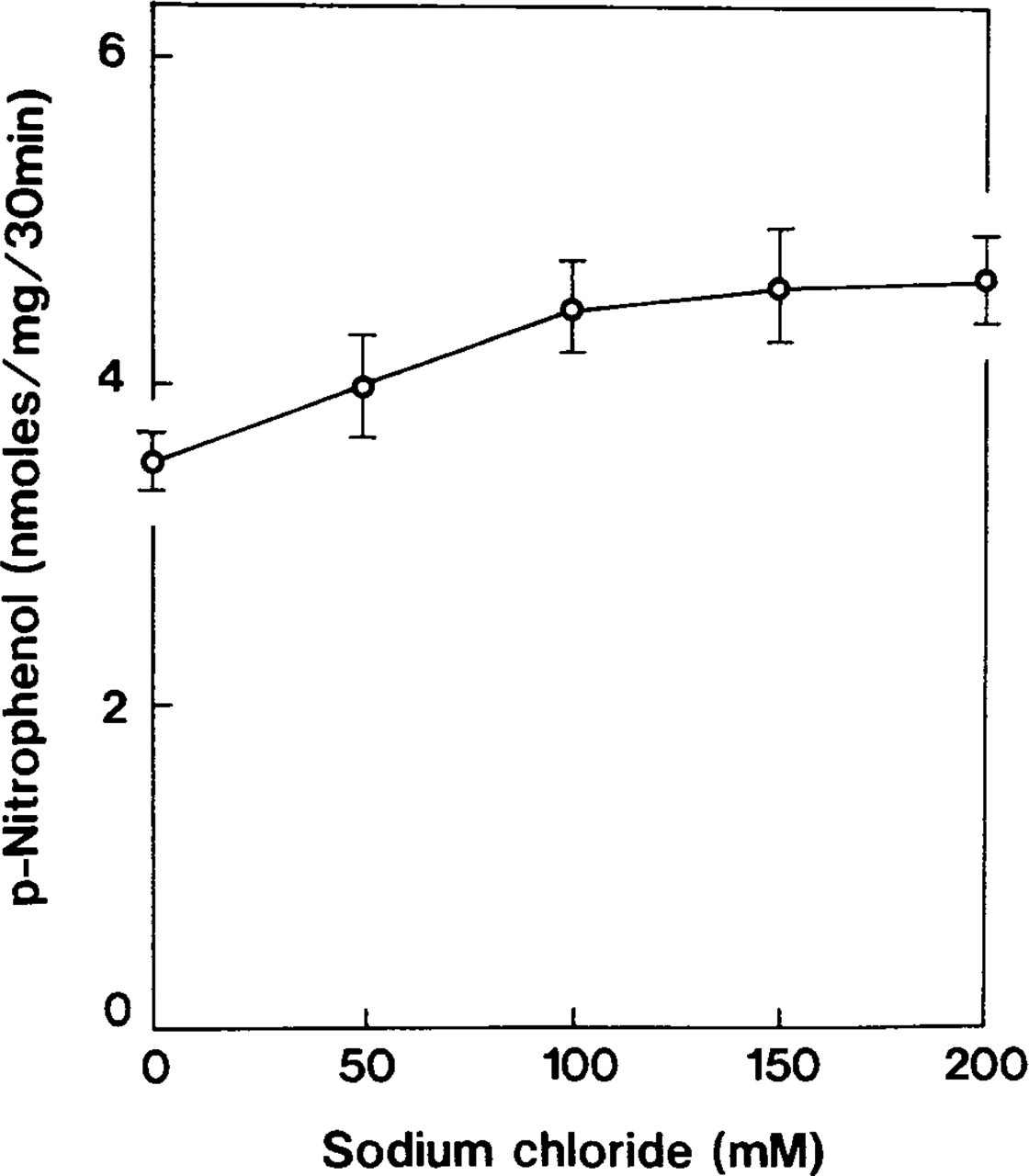
Control study on ouabain-insensitive p-NPPase; incubation in a medium in which potassium ions were excluded by sodium replacement. p-nitrophenol accumulation forms approximately 30% of that obtained from the reaction medium. Symbols represent mean ± SE of measurements performed in four separate experiments.
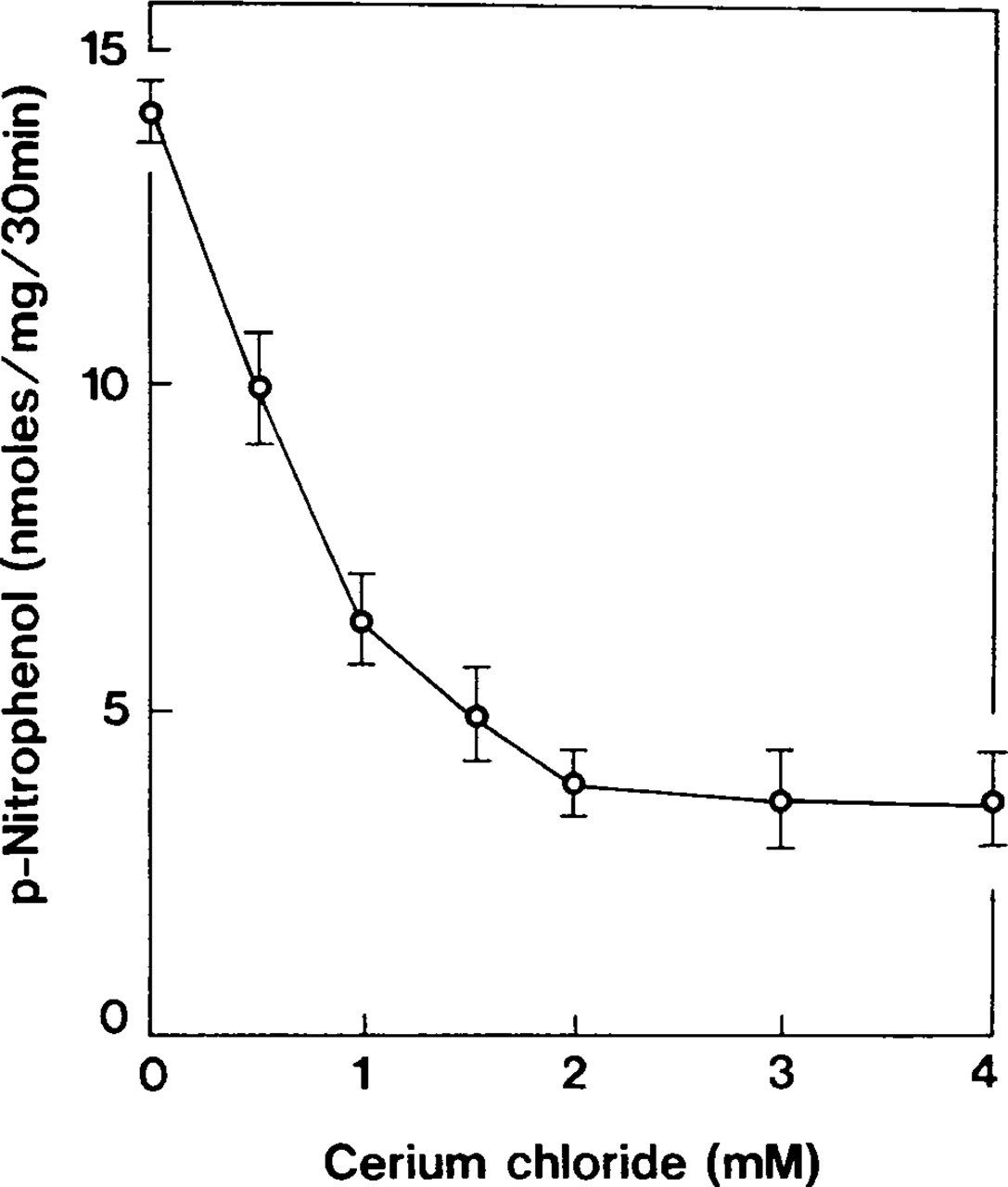
Effect of cerium concentrations on accumulation of p-nitrophenol generated by ouabain-insensitive, K-dependent p-NPPase. The accumulation of p-nitrophenol is decreased with an increase in cerium concentration, but constitutes almost 30% of the total activity. Symbols represent mean ± SE of measurements performed in four separate experiments.
In biochemical controls, the liberation of p-nitrophenol decreased to 18%, 64%, and 60% using 10 mM sodium orthovanadate, 1 mM DCCD, and 1 mM omeprazole, respectively (Table 3).
Cytochemical Assays
Cytochemical experiments revealed RP at sites of ouabain-insensitive, K-dependent p-NPPase (H,K-ATPase) activity as an electron dense precipitate that was detected at the inner surface of the plasma membrane and at the T-tubules (Figures 6a-6c). Distribution of the RP varied in different regions of the plasma membrane. At regions adjacent to neighboring myocytes, RP was localized along membranes (Figure 7a). Regions of the membrane adjacent to pericapillary spaces displayed RP predominantly at the plasma membrane invaginations (Figure 7b). Other portions of the plasma membrane in these regions often did not exhibit RP. No reaction was observed at secretory granules, mitochondria, and vacuoles.
In cytochemical controls, no H,K-ATPase activity was detected in ventricular myocytes (Figure 8a). In atrial myocytes, this activity was significantly inhibited by 10 mM sodium orthovanadate (Figure 8b). Both 1 mM omeprazole and 1 mM DCCD strongly repressed H,K-ATPase activity (Figures 8c and 8d). Exclusion of CaCl2 from the reaction medium stimulated substrate hydrolysis at the sarcoplasmic reticulum (SR) (Figure 8e). Incubation in a substrate-free medium (Figure 8f) or in a heated medium resulted in complete enzyme inhibition. Increase of the KCl concentration from 50 mM to 100 mM did not essentially change the cytochemical reaction. A weak sodium-stimulated and furosemide-inhibited p-NPPase activity detected biochemically was not evaluated by this cytochemical procedure because of the weakness and considered inhibitory effect of the capture agent. We found that 2 mM furosemide did not influence the K-dependent activity.
Amount of p-nitrophenol liberation in control experiments a
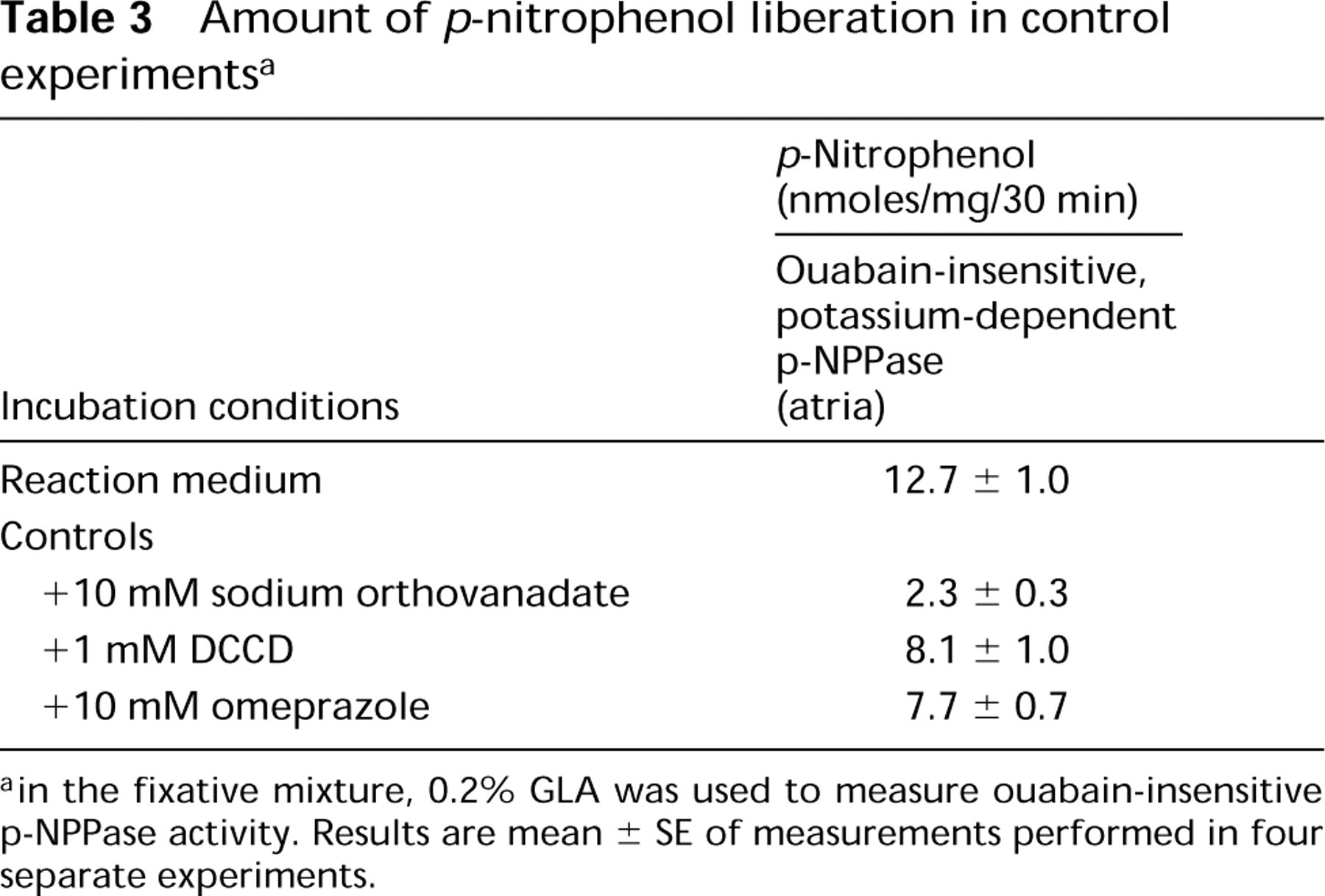
ain the fixative mixture, 0.2% GLA was used to measure ouabain-insensitive p-NPPase activity. Results are mean ± SE of measurements performed in four separate experiments.
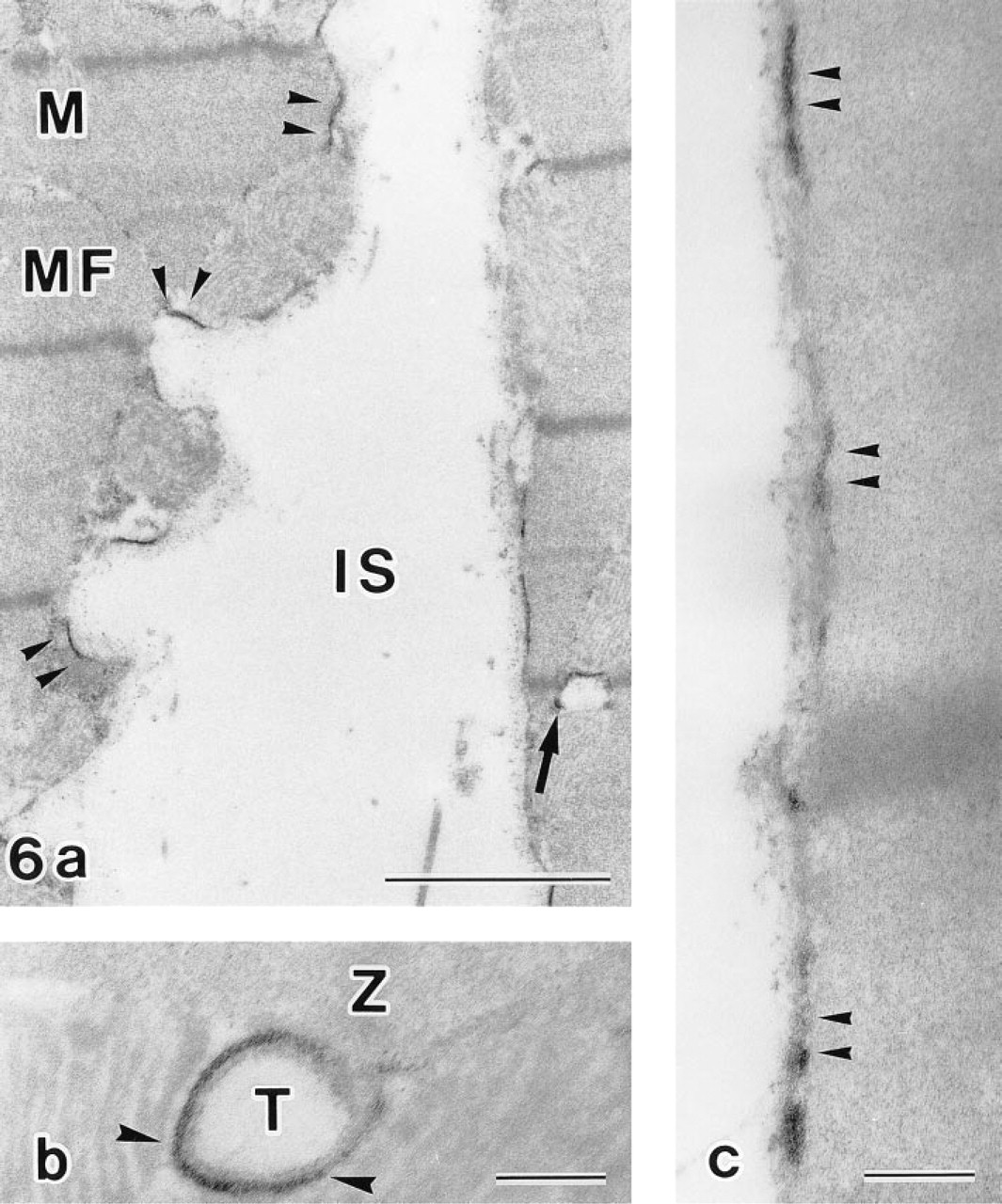
Incubation with a reaction medium for demonstration of ouabain-insensitive, K-dependent p-NPPase. RP is detected at the plasma membrane (arrowheads) and at the T-tubules (arrow) (
X-ray Microanalysis
The presence of cerium within the product of the cytochemical reaction was verified by electron probe X-ray dispersive microanalysis. The X-ray spectrum of the precipitate indicated the presence of cerium as well as phosphorous and oxygen peaks, and confirmed the composition of the RP as cerium phosphate (Figure 9a). Neither cerium nor phosphorous peaks were detected in areas without reaction (Figure 9b).
Discussion
We report here that in the absence of classical ouabainsensitive, K-dependent p-NPPase (Na,K-ATPase), atrial myocytes represent biochemically and cytochemically detectable ouabain-insensitive, K-dependent p-NPPase activity that is similar to that of H,K-ATPase. This may indicate that these excitable cells are involved in proton–potassium exchange.
In this study we employed cardiac muscle from rat because investigations of rat heart are considerably more advanced and the results obtained can be easily interpreted (Sweadner et al. 1994). Rat heart has been demonstrated to possess multiple ouabain binding sites of both high- and low-affinity types (Akera et al. 1986). High-affinity binding sites, however, form a minority and do not contribute to high sensitivity to ouabain of rat cardiac muscle in total (Noel and Godfraind 1984). This view is based on results of biochemical experiments with unfixed tissue, which is poorly suitable for histochemical studies because adequate morphological preservation and therefore reliable demonstration of enzyme reactive sites is hard to achieve. In this study we utilized fixed tissue. A contribution of Na,K-ATPase was fully eliminated by several consistent steps, such as inhibitable fixation, preincubation with ouabain, and inclusion of ouabain as a component of the reaction medium. These conditions are generally agreed to be inhibitory for Na,K-ATPase (Rostgaard and Møller 1980; Ernst and Philpott 1970). Efficiency of the elimination of Na,K-ATPase was confirmed in control experiments with ventricular myocytes. To avoid doubts concerning the contribution of Na,K-ATPase to H,K-ATPase in the course of this study, we also examined the generally accepted highly ouabain-sensitive heart tissue of guinea pigs. In parallel experiments with guinea pigs atrial and ventricular myocytes, similar patterns of ouabain-insensitive, K-dependent p-NPPase (H,K-ATPase) activity in response to increased stringency of fixation conditions were biochemically defined in atrial myocytes. No H,K-ATPase activity was found in ventricular myocytes. Cytochemically, we have also succeeded in detecting this enzyme in guinea pigs atrial myocytes, whereas virtually no reaction was found in ventricular myocytes (data not shown). This enabled us to analyze the biochemical properties and cytochemical localization of H,K-ATPase activity in rat atrial myocytes under the conditions of the present experiment without concern that Na,K-ATPase is implicated, and suggested that the existence of the proton-translocating, potassium-dependent ATPase system in these cells is physiologically determined.
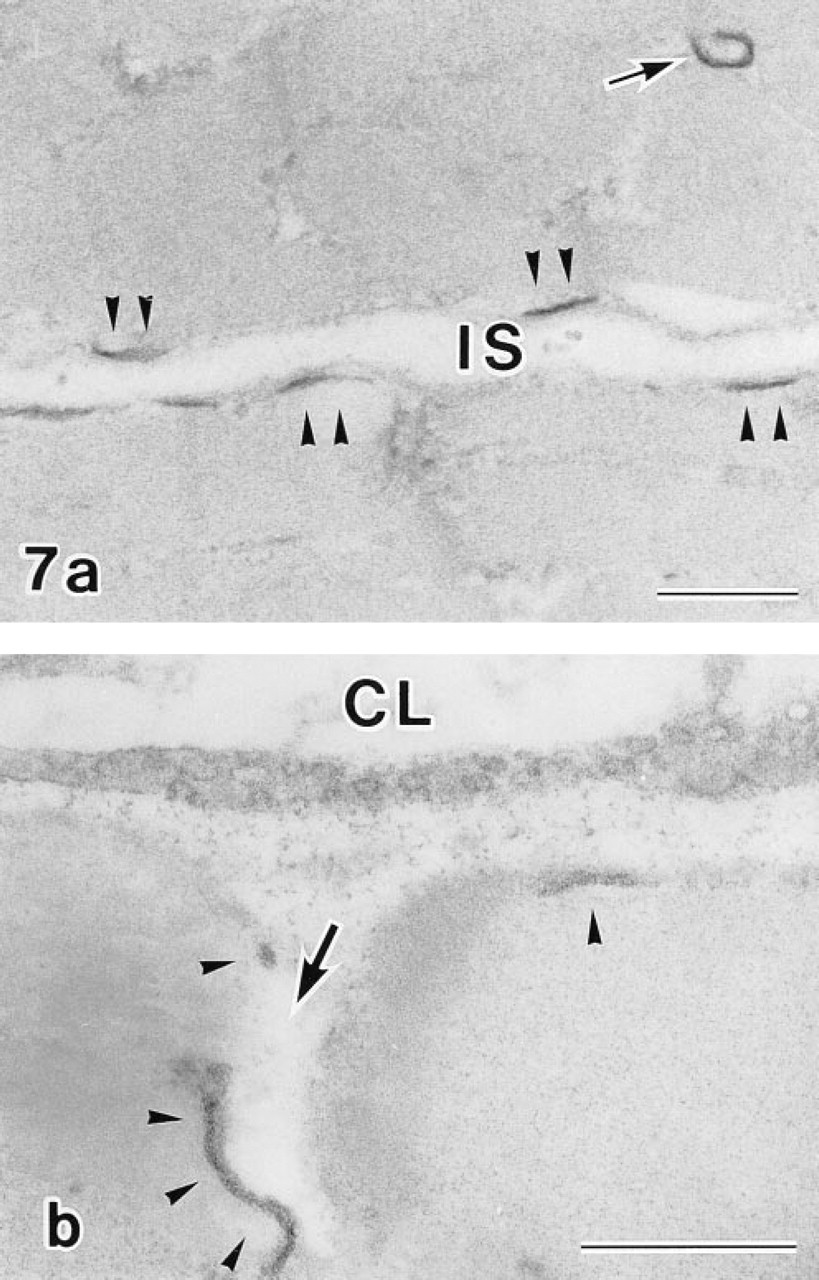
Electron micrographs of two different regions of the plasma membrane adjacent to a neighboring myocyte (
Various controls were employed in this study to determine the properties of the ouabain-insensitive, K-dependent p-NPPase activity in rat atrial myocytes. Ouabain, levamisole, and CaCl2 were used as components of the reaction medium. In addition, sodium orthovanadate, DCCD, NaCl, furosemide, and omeprazole were also tested. A high calcium concentration is believed to have an inhibitory on Ca-ATPase (Rega et al. 1973). In control cytochemical experiments when CaCl2 was omitted from the incubation medium, we observed substrate hydrolysis at the SR and a decrease in H,K-ATPase activity at the plasma membrane. This may be due to the fact that atrial myocytes contain more calcium and share more extensively developed SR than ventricular myocytes (Fukuda 1975). Therefore, under the conditions of this experiment, inhibition of Ca-ATPase should be employed to avoid precipitation that may be related to this enzyme. Sodium orthovanadate, DCCD, and omeprazole are known inhibitors of H,K-ATPase (Lorentzon et al. 1987; Rabon et al. 1985; Sachs et al. 1982). In this study, sodium orthovanadate displayed a very strong inhibitory effect. It abolished 82% of the activity and therefore firmly established the nature of the H,K-ATPase as a P-type enzyme (Rabon et al. 1985). DCCD displayed the least effect of all chemicals tested, possibly because inhibition by DCCD demands stronger fixation. Omeprazole is believed to be an effective inhibitor of H,K-ATPase (Wallmark et al. 1985). Liberation of p-nitrophenol in experiments with omeprazole constituted approximately 60% of the maximum, which was a little less than with DCCD but significantly more than with sodium orthovanadate. Taken together, all these inhibitors demonstrated reliable inhibitory properties. However, the relation between them is not the same as has been reported for gastric cells (Kobayashi and Seguchi 1990). This discrepancy may be due to diversity of H,K-ATPase isoforms in various tissues and to the methodologies employed.
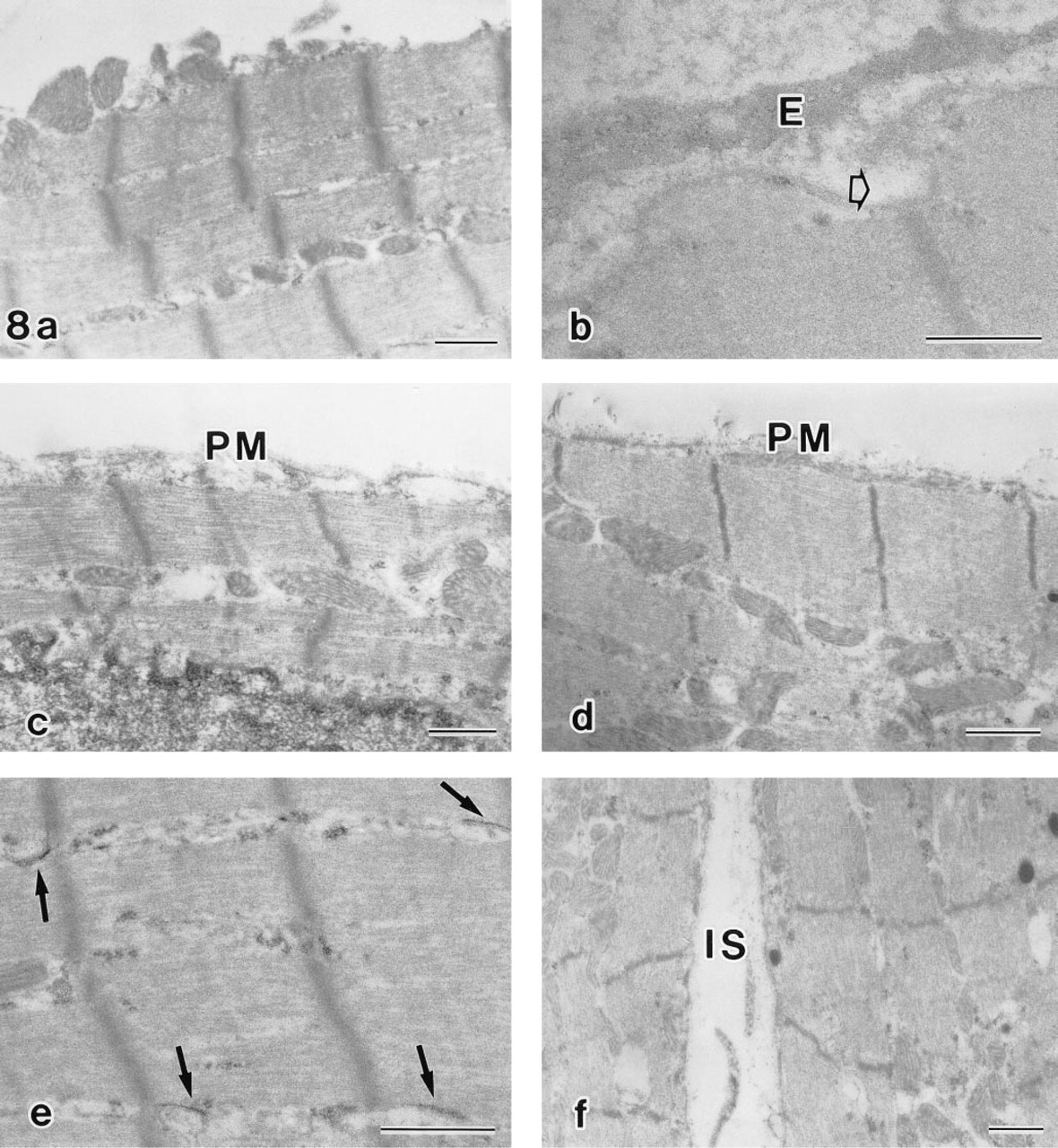
Control cytochemical experiments. No ouabain-insensitive, K-dependent p-NPPase (H,K-ATPase) activity was detected in ventricular myocytes (
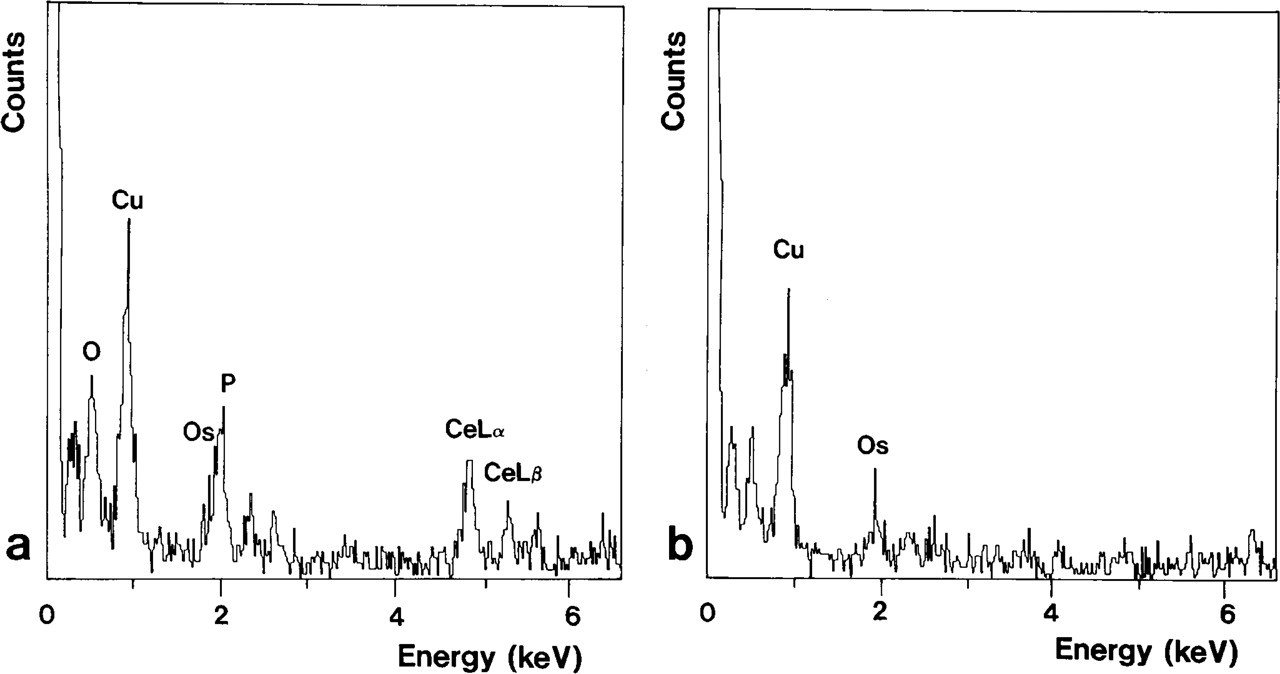
X-ray spectra of areas with (
As a methodological basis, we applied the cerium-based technique of Kobayashi and Seguchi (1990), designed for use at physiological pH, because our biochemical assays have demonstrated the highest H,K-ATPase activity within this range. p-NPP served in our study as a substrate that provides a valuable combination of biochemical and cytochemical experiments that can be precisely quantified and controlled. Cerium has now a deserved reputation as the preferred capture agent in phosphatase cytochemistry. It enables one to investigate unknown aspects of cell metabolism by providing precise localization of the RP and by forming fine primary cerium precipitates (Van Noorden and Frederiks 1993). Production of the final RP is proportional to enzyme activity (Robinson and Karnovsky 1983), so the latter can be quantified in situ. The reason for using this capture agent in heart tissue is justified also by the fact that the cerium-based techniques are efficient probes in pathophysiological cardiovascular research (Shlafer et al. 1990). In this study, it was particularly useful to employ this technique for studying unaffected tissue. The residual ouabain-insensitive, potassium-dependent p-NPPase (H,K-ATPase) activity constituted in our cytochemical experiment approximately 30%, which is markedly higher than that reported for lead-based H,K-ATPase cytochemistry (Fujimoto et al. 1986) and is sufficient for cytochemical demonstration.
On the basis of our biochemical, cytochemical, and X-ray analytical data, we conclude that the enzyme activity detected under the conditions of our experiment is an expression of ouabain-insensitive, K-dependent p-NPPase (H,K-ATPase) (E.C. 3.6.1.36). Two distinctively different ATPases are recognized to date as being responsible for proton translocation: plasma membrane (P-type) H,K-ATPase and vacuolar (V-type) H-ATPase (Pedersen and Carafoli 1987a,b). The latter is K-independent, does not form phosphorylated intermediates, and functions mainly for acidification. The presence of this enzyme in rat atrial myocytes has been suggested by Somlyo et al. (1988), because atrial specific granules have a high calcium content, are acidic, and maintain anion gradients. Both biochemical and cytochemical experiments in this study, however, revealed that the detected enzyme is K-dependent and is similar to P-type H,K-ATPase. The cytochemical reaction demonstrated RP localized to the inner surface of the plasma membrane. The site of this localization is in good agreement with biochemical data showing that ATP-binding sites of P-type ATPases are located at the cytoplasmic side of the plasma membrane and represent high amino acid homology (Farley and Faller 1985). We did not observe the RP at secretory granules, probably owing to differences in the methodological approach, such as fixation conditions, neutral pH of the incubation medium, and the presence of various inhibitors.
Physiologically, the H,K-ATPase can be involved in the intracellular regulation of pH and may be associated with the metabolism of specific secretory granules. It is known that atrial and ventricular myocytes differ in their function and sensitivity to inhibitors of ATPases (Grupp et al. 1979). Atrial myocytes contain specific granules and possess secretory function (Needleman et al. 1985). Atrial natriuretic factor (ANF) is released from these granules into the blood and regulates intracardiac blood pressure and electrolyte balance (Inagami 1989; Atlas 1986). Despite the established fact that most effects of ANF are mediated by membrane-bound rather than by cytosolic enzymes (Hamet et al. 1984), the exact metabolic constitution of the plasma membrane of atrial myocytes remains far from being elucidated. In this respect, diversities in the localization of the RP at the plasma membrane may indicate specialized functions of these regions of the membrane. This feature of the cytochemical reaction may show that the H,K-ATPase participates in the physiological pathway that is related to transport into the vascular lumina, being involved in the externalization of atrial secretory peptides. Nevertheless, when we observed the cytochemical reaction at this portion of the plasma membrane, peripheral granules were usually not located there. This, taken together with the known fact that the sequence of the active ANF peptide contains no acidic residues, gives us a basis for concluding that the relation between the activity of H,K-ATPase and the metabolism of atrial secretory granules is rather indirect and is mediated by another regulatory system(s). It can be also construed as strong evidence that the detected RP does not contain molecules that were secreted or/and spilled together with ANF.
Our biochemical and cytochemical findings imply that H,K-ATPase activity may be related to atrial potassium homeostasis. Whether this H,K-ATPase is elec-trogenic or neutral remains to be determined. However, even generation of a membrane potential can not be considered an obvious proof for the electrogenicity of the proton ATPase, since this potential may be produced by a secondary effect of a pH gradient generated by other neutral ATPases (Al-Awqati 1986).
In summary, using a biochemically optimized cytochemical medium with cerium as capture agent, we have studied H,K-ATPase activity in rat atrial myocytes. We suggest that the detected P-type enzyme is a functionally active isoform of H,K-ATPase and may be involved in the proton–potassium exchange in atrial myocytes. Further studies may be required for the demonstration of immunocytochemical parallels between this enzyme and well-established H,K-ATPases that will provide better understanding of its function.
Footnotes
Acknowledgements
We wish to thank Mr Patrick C. Nahirney for help in preparing this manuscript. Supported by Grants-in-Aid for Scientific Research (C) (Nos. 07807101 and 07670021) and for University-to-University Cooperative Research from the Ministry of Education, Science, Sports and Culture of Japan.