
Abstract
The variable coding sequence (VCS) multigene family encodes diverse salivary proteins, such as the SMR1 prohormone and the PR-VB1 proline-rich protein in the rat. In situ hybridization was used to study the cell-specific expression of two new mouse VCS genes, Vcs1 and Vcs2. We show that the Vcs1 transcripts, which code for a proline-rich protein, MSG1, are highly abundant in male and female parotid glands, in which they are specifically detected in acinar cells. No expression was seen in the submandibular or sublingual glands. In contrast, Vcs2 transcripts were found only in the acinar cells of the submandibular glands (SMGs) of male mice, in which they are expressed in response to androgens. Expression was found to be heterogeneous within acinar structures. No Vcs2 transcripts were detected in the SMGs of females or castrated males by Northern blot, RNase protection, or in situ hybridization. Androgen administration to females or castrated males induced expression at a level comparable to that of intact males. The Vcs2 gene is the first example of a mouse androgen-regulated gene that is expressed in SMG acinar cells. This result, in addition to our previous observation on SMR1 expression in rats, demonstrates that both acinar cells and granular convoluted tubule (GCT) cells are target cells for androgen action in rodent SMG.
Keywords
S
The VCS (variable coding sequence) family was recently described in rats (Rosinski-Chupin and Rougeon 1990; Courty et al. 1994,1996). Members of this family have also been found in mice and humans (Isemura and Saitoh 1994; Tronik-Le Roux et al. 1994) and are mainly expressed in the salivary glands. A major characteristic of the genes of this family is the presence of a hypervariable region inside the coding sequence. Interestingly, in the VCS genes, the 3′-untranslated region of the mRNA and most of the coding sequence, except that for the signal peptide, correspond to a single exon (exon III). In intraspecies pairwise comparisons, a higher level of sequence divergence is observed in the hypervariable region than in the adjacent exonic or intronic sequences. Furthermore, most of the mutations at the nucleotide level lead to amino acid substitutions (Courty et al. 1994,1996). As a consequence, the VCS family encodes proteins that are very diverse in amino acid content, structure, and probably function. The first gene of this family to be described was the rat VCSA1 gene, which is expressed mainly in SMG and encodes the SMR1 protein (Rosinski-Chupin et al. 1988). SMR1 is a prohormone protein, whose maturation generates three related peptides, which are released into the saliva and bloodstream (Rougeot et al. 1994). The target organs for the final maturation product, a pentapeptide, were recently shown to be primarily kidney and the bone, suggesting that this peptide might be involved in the modulation of mineral balance (Rougeot et al. 1997). The synthesis and secretion of the mature peptides are under neuronal and hormonal control, in particular androgen regulation. The release of the peptides from the SMG into the bloodstream, by a yet unknown mechanism, is reminiscent of what is seen for renin, NGF and EGF in mice and some kallikreins in rats (Bing and Poulsen 1979; Aloe et al. 1986; Berg et al. 1990). Similarly to these proteins, SMR1 and its maturation peptides are synthesized at a higher level in male than in female rats and therefore might have a function linked to male physiology. However, in contrast with those proteins, which are all produced in the granular convoluted tubules of the gland (reviewed in Gresik 1980), VCSA1 transcripts are found only in acinar cells (Rosinski-Chupin et al. 1993).
Recently, we began the characterization of the VCS family in the mouse. We cloned two cDNAs from SMGs, corresponding to the MSG1 and MSG2α proteins (Tronik-Le Roux et al. 1994). Based on the sequence of the cDNA, MSG1 was predicted to belong to a new group of salivary proline-rich proteins encompassing the PR-VB1 protein of rat (Courty et al. 1994) and the precursor of the salivary P-B peptide of humans (Isemura et al. 1979; Isemura and Saitoh 1994), which are also both encoded by members of the VCS family. The gene encoding the MSG1 protein was named Vcs1.
The Vcs2 gene differs in its structure from other VCS genes; this gene results from the juxtaposition in the same transcription unit, by a complex DNA rearrangement, of sequences from three different VCS genes (Señorale-Pose and Rougeon, 1997) resulting in a composite structure consisting of six exons (exons I, II, IIa, III, II′, and III′). Alternative splicing of the primary messenger generates at least five different transcripts. Putative proteins encoded by these transcripts have only a low level of sequence similarity. One of these, the MSG2α protein, is predicted to be secreted and matured at dibasic sites to generate, like SMR1, a five or six amino acid-long peptide.
Here we study the expression of Vcs1 and Vcs2 mouse genes in the salivary glands, in particular the cellular distribution of the transcripts, by in situ hybridization. We show that the Vcs1 gene, encoding the proline-rich protein MSG1, is mainly expressed in the acinar cells of the parotid glands. In contrast, the Vcs2 gene, encoding MSG2α, appears to be expressed only in the submandibular gland acinar cells of male mice.
Materials and Methods
Animals and Tissue Preparation
Male and female Balb/c mice were obtained from Iffa Credo (Lyon, France). To evaluate the effects of androgens on VCS gene regulation, male, female and castrated mice were purchased at 5 weeks and kept in house for 4 weeks. The 9-week-old mice were then injected
Submandibular, sublingual, and parotid glands were rapidly removed and processed as previously described (Rosinski-Chupin et al. 1993) for fixation and paraffin embedding. After embedding, 5-μm thick sections were obtained using a microtome and mounted on gelatin-coated slides. For RNA preparation, SMGs were rapidly removed and frozen at −80C until RNA extraction.
Northern Blot Analysis
RNA from submandibular glands of 8-week-old male (n = 5) and female (n = 5) Balb/c mice were prepared according to Chomczynski and Sacchi (1987). Glyoxal-denatured RNA (100 μg) was resolved by electrophoresis in a 1.1% agarose gel (Gründemann and Koepsell 1994) and blotted on Hybond N membrane (Amersham; Arlington Heights, IL) as described (Chomczynski 1992). For analysis of androgen regulation, RNAs were prepared and analyzed as previously described (Rosinski-Chupin et al. 1988). For each group of animals, five individual preparations of RNA were pooled and analyzed. The membrane was used successively with different probes, after dehybridization according to manufacturer's instructions. The different probes, corresponding to exon III (Ava II-Eco RI fragment of Vcs2α cDNA, GenBank accession number U 82380), exon III′ (227-
Plasmids for In Vitro Transcription
Two sequences of the Vcs2 gene were used to generate plasmids for in vitro transcription. The first corresponds to a 364-
RNase Protection
Three RNA probes were generated by in vitro transcription in the presence of [α-32P]UTP, according to manufacturer's instructions (Stratagene; La Jolla, CA) of (a) pcV2a from the SP6 promoter after linearization by Xba I, (b) pcV2b from T7 promoter after linearization by Bam HI, and (c) pAct from SP6 promoter after Xba I linearization.
RNase protection assay was performed using RNase I (Promega; Madison, WI) according to the manufacturer's instructions. Briefly, 20 βg of total RNA was hybridized with a mixture of the three probes (200,000 cpm of each; specific activity of each probe 160 Ci/mmol) for 22 hr at 45C in 30 μl of hybridization buffer (80% deionized formamide, 40 mM PIPES, pH 6.4, 0.4 M sodium acetate, pH 7, 1 mM EDTA). After hybridization the samples were digested by 5 U of RNase I for 1 hr at 37C in 300 μl of 10 mM Tris-HCl, pH 7.5, 5 mM EDTA, 200 mM sodium acetate, pH 7). Digestions were stopped by addition of SDS (final concentration 0.07%) and tRNA (40 μg/ml final). After ethanol precipitation, the samples were resolved in a 6% polyacrylamide sequencing gel. For each group of animals, five individual preparations of RNA were separately analyzed.
In Situ Hybridization
35S-Labeled RNA probes were synthesized in a 25-μl volume, using 500 ng of linearized plasmid and 50 μCi of [α-35S]-UTP (1000 Ci/mmol) in the presence of 40 mM Tris-HCl, pH 7.5, 2 mM spermidine, 6 mM MgCl2, 10 mM NaCl, 10 mM dithiotreitol, 0.6 mM each of ATP, GTP, CTP, and 40 U of T7 or SP6 RNA polymerase. No hydrolysis of the probes was performed. Slides were postfixed by immersion in a 4% paraformaldehyde solution in PBS, pretreated with 1 μg/ml proteinase K at 37C in 20 mM Tris-HCl, pH 7.5, 20 mM CaCl2 for 25 min, and acetylated before hybridization. Hybridization was performed overnight at 52C as previously described, using 50,000 cpm/μl of probe in 50% formamide, 0.3 M NaCl, 20 mM Tris-HCl, pH 7.4, 5 mM EDTA, 10 mM NaH2PO4, pH 8.0, 10% dextran sulfate, 1 X Den-hardt's solution, and 0.5 mg/ml yeast RNA. Washings were as previously described (Rosinski-Chupin et al. 1993). After dehydration, the slides were dipped in NTB2 emulsion (Kodak; Chalon-S-Saone, France) diluted V/V in 0.6 M ammonium acetate, exposed for 1–7 days at 4C, developed in D19 (Kodak) for 5 min, fixed in Unifix (Kodak), and counter-stained with hematoxylin (Rhone-Poulenc; Vilers St Paul, France).
In situ hybridization experiments with Vcs1 digoxigenin-labeled RNA probes were performed as previously described (Rosinski-Chupin et al. 1993). After incubation with anti-digoxigenin F(ab) fragments labeled with alkaline phosphatase (Boehringer Mannheim; Meylan, France), alkaline phosphatase activity was detected by reaction with nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolylphos-phate.
Results
Expression of the Mouse Vcs1 Gene in Mouse Parotids
In situ hybridization experiments were performed on mouse salivary glands sections using a Vcs1 cRNA probe into which either a radiolabeled (Figure 1a) or a digoxigenin-labeled (Figure 1b) ribonucleotide was incorporated. This probe, designed to specifically detect Vcs1 transcripts, corresponds to the coding region of Vcs1 exon III. Parotid sections were positive in both male and female mice after a 1-day exposure time. No sex-linked difference in expression was observed (data not shown). The signal was completely abolished by RNase pretreatment and was not detected after hybridization with sense Vcs1 RNA probe (data not shown), demonstrating that this signal is specific for Vcs1 transcripts. Acinar cells were determined as the site of Vcs1 expression in mice parotids. No expression was detected on the parotid ductal cells (Figure 1b). No effect of castration or androgen injection was observed on the intensity of the signal in parotid acinar cells (data not shown).
Surprisingly, although Vcs1 cDNA was cloned from mouse submandibular gland tissue (Tronik-Le Roux et al. 1994), we failed to detect a specific signal (i.e., RNase-sensitive and not detected by a sense probe) in these glands and in sublingual gland sections (Figure 1a). One possible explanation is that, because of the proximity between submandibular and parotid glands, SMG mRNA preparations may have been contaminated by parotid tissue. However, we cannot exclude a very low level of expression, near background level, in this tissue.
Vcs2 mRNAs Are Differentially Expressed in Submandibular Glands of Male and Female Balb/c Mice
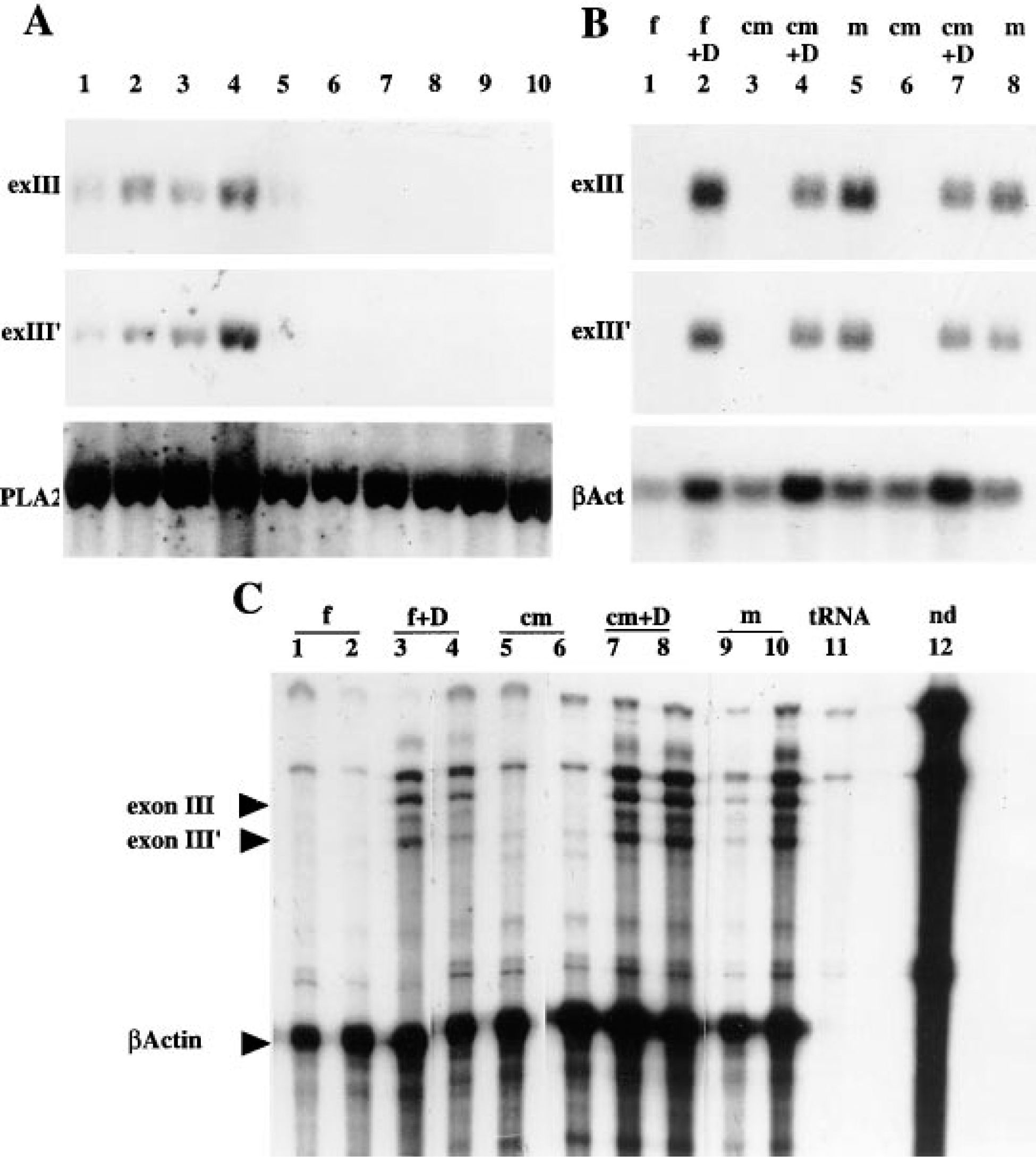
Vcs2 transcripts were previously characterized from SMG of male mice (Señorale-Pose and Rougeon: 1997). To determine if there is a sex-linked difference of Vcs2 expression, Northern blot analysis was performed using two different probes (pIII and pIII′), corresponding to two different exons (exon III and exon III′) of the Vcs2 gene (Figure 2) with sequences divergent to other VCS genes, to avoid cross-hybridization. As shown in Figure 2, these two probes hybridize to different Vcs2 mRNA species and the combination of the two probes enables the detection of all known Vcs2 transcripts. With each probe, transcripts of approximatively 0.7
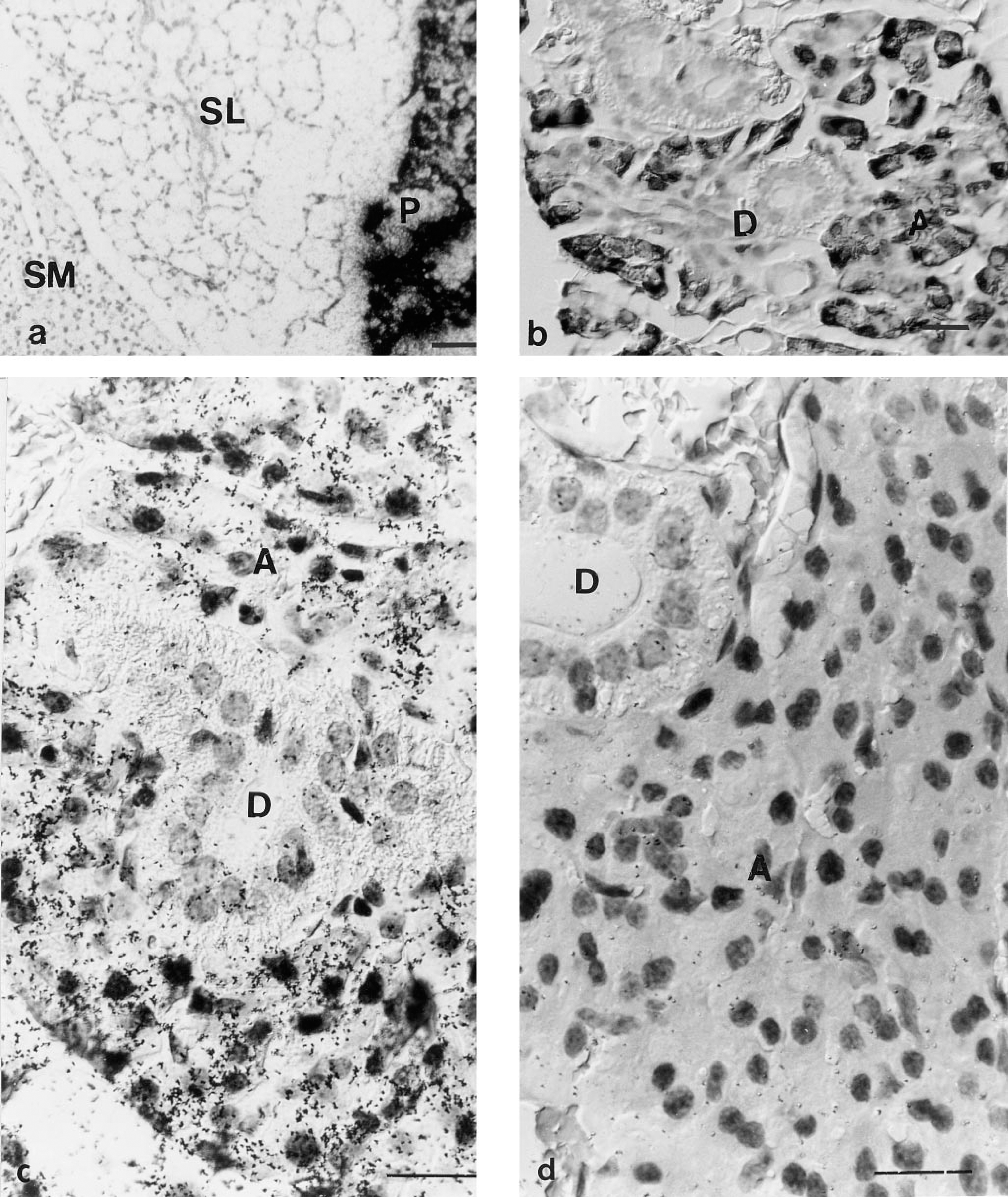
Localization of Vcs1 transcripts by in situ hybridization. (
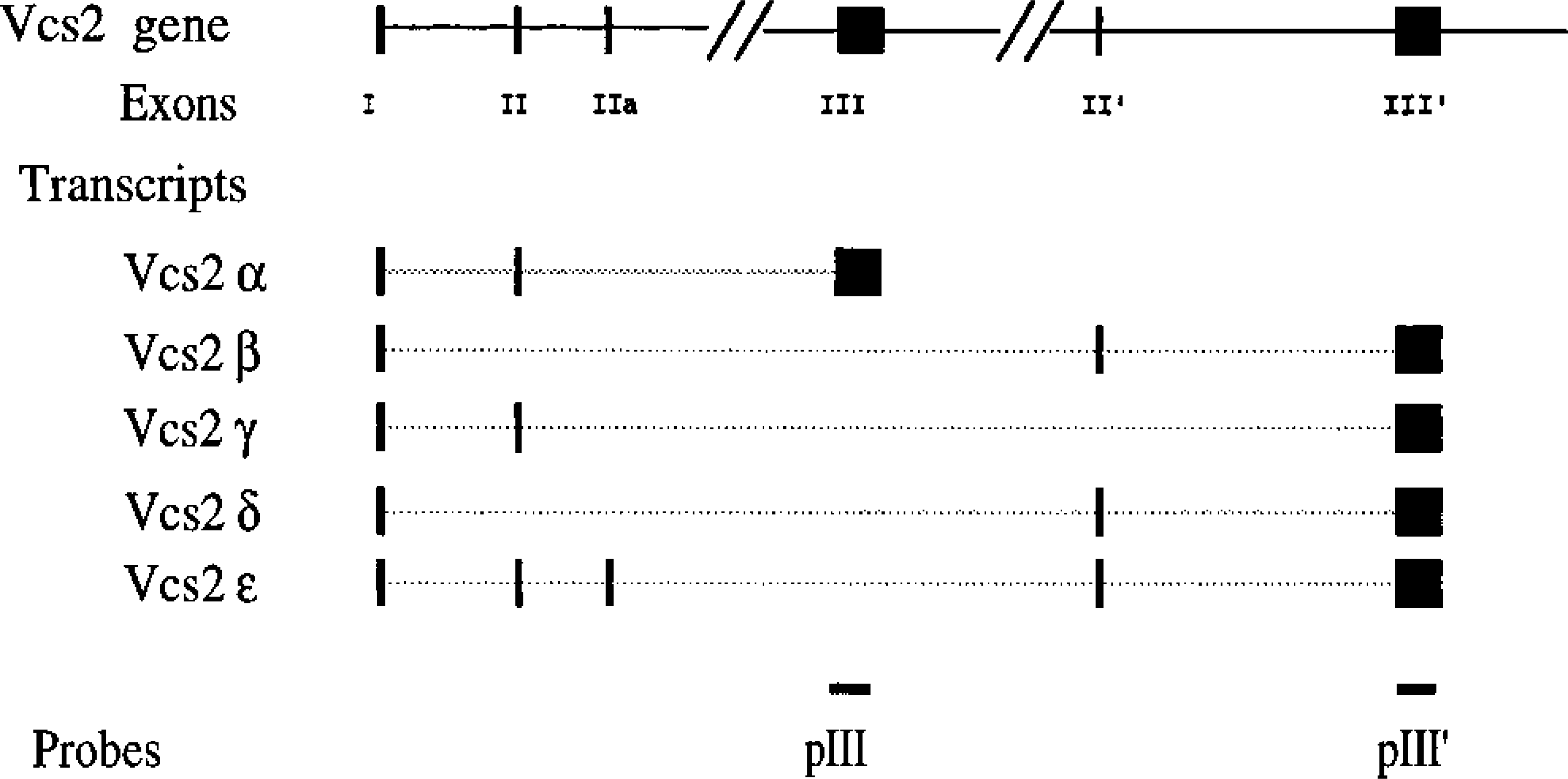
DNA fragments used to generate mouse Vcs2-specific probes. Schematic representation of the structure of the Vcs2 gene. Exons are represented by large bars. The different transcripts obtained from the Vcs2 gene are shown, as well as the two DNA fragments that were subcloned and used as probes in the in situ hybridization experiments and RNase protection assays.
To analyze the role of androgens in this sex-linked difference of expression, we performed castration and androgen injection experiments. SMG RNA preparations were analyzed by Northern blot with both probes. As shown in Figure 3B (Lanes 3, and 6), castration results in the disappearance of SMG-specific Vcs2 transcripts. Androgen injection for 1 week to castrated males (Figure 3B, Lanes 4 and 7) or to females (Figure 3B, Lane 2) results in the induction of expression to a level similar to males. RNase protection experiments confirmed that the signals were specific to Vcs2 transcripts and gave the same results as Northern blot experiments (Figure 3C). Bands specific for both probes were only seen in males (Figure 3C, Lanes 9 and 10), androgen-treated females (Figure 3C, Lanes 3 and 4), or androgen-treated castrated males (Figure 3C, Lanes 7 and 8) and not in females (Figure 3C, Lanes 1 and 2) or castrated males (Figure 3C, Lanes 5 and 6) treated with vehicle alone.
Hybridization of male Balb/c mouse SMGs with a 35S-radiolabeled RNA probe corresponding to exon III of the Vcs2 gene resulted in a signal localized in acinar cells (Figure 4). Not all the acinar cells were equally labeled, and inside a single acinus there was cell-to-cell variations. No signal above background could be detected either in the ducts of the SMGs or in the adjacent sublingual and parotid glands. The specificity of the signal for SMG acinar cells was assayed by hybridization with a sense probe and no signal was detected under these conditions (Figure 5e).
In situ hybridization of mouse SMGs with an anti-sense RNA probe corresponding to exon III′ of Vcs2 gave similar results as with the exon III probe (Figure 5b). In addition, a similar pattern of positive cells was revealed by hybridizing serial sections of SMGs with either probe, showing that different types of Vcs2 transcripts are found in the same cells (Figures 5a and 5b).
In contrast, no signal was detected in the glands of female mice using either probe (Figure 5c and 5d). Castration resulted in the almost total disappearance of positive cells on sections of male SMGs (Figure 5f). Treatment with dihydrotestosterone of either females (Figure 5g) or castrated males (Figure 5h) led to the induction of expression of Vcs2 transcripts in a relatively high number of acinar cells, but not all acinar cells were positive.
Analysis of Vcs2 expression. (
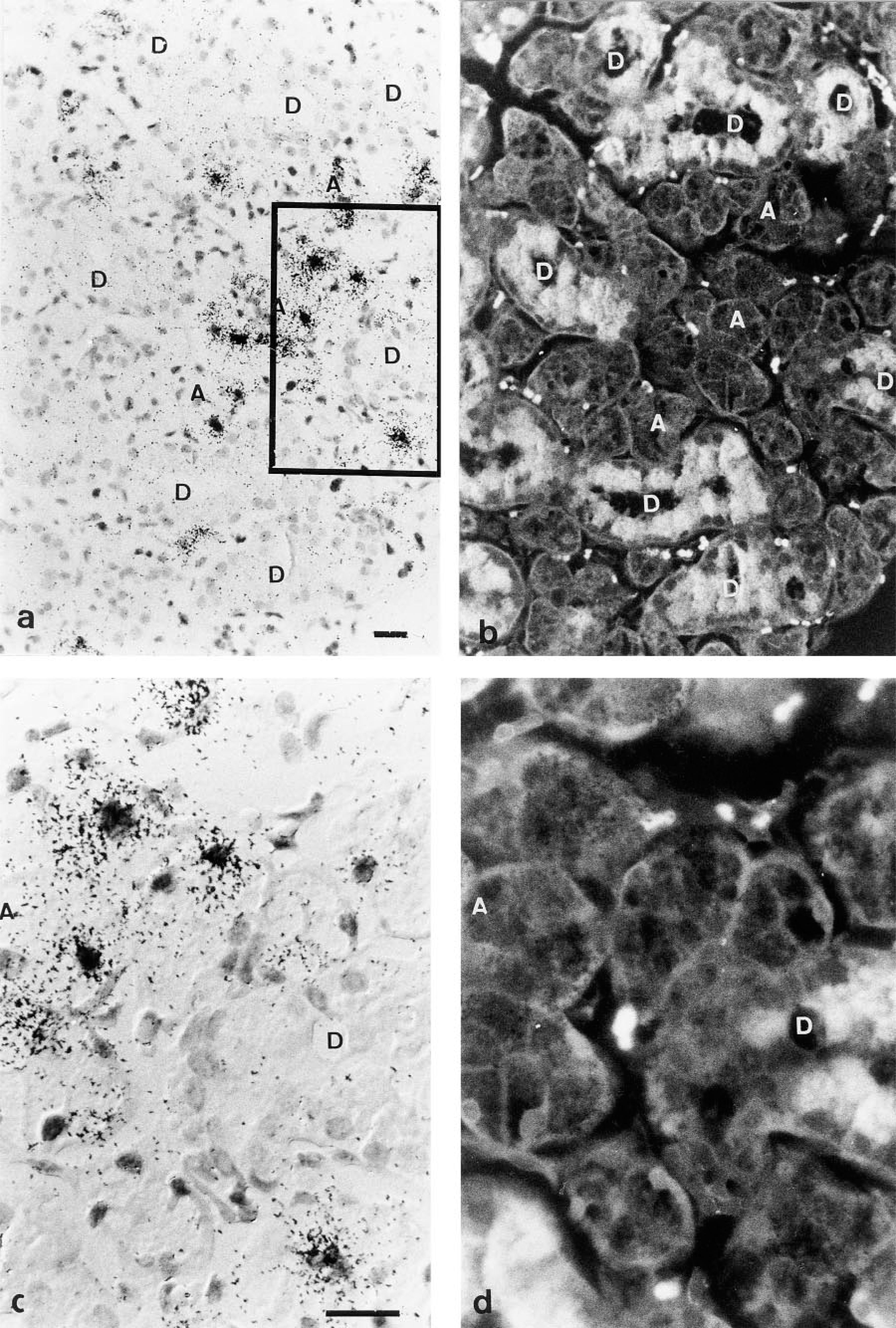
Detection of Vcs2 transcripts on an SMG section of male mouse by in situ hybridization. The probe is a 35S-labeled Vcs2 exon III anti-sense RNA probe. (
Discussion
Our aim was to localize, by in situ hybridization, transcripts of two genes of the mouse VCS family. These two genes encode proteins differing in their sequence and probably in their function. Here we show that these two genes are differentially expressed in the salivary glands. Whereas Vcs1 transcripts are expressed at a high level in the acinar cells of parotids, Vcs2 transcripts are expressed at a low level in the acinar cells of the SMG.
The Vcs1 gene was previously shown to encode a proline-rich protein. Proline-rich proteins are abundant in the saliva, well-characterized in humans, rats and mice (Ann and Carlson 1985; Clements et al. 1985; Carlson 1988; Lin and Ann 1991; Azen 1993; Kim et al. 1993). They contain a repeat segment rich in proline. Many characterized salivary proline-rich proteins belong to the same multigene family. Notable exceptions are the proline-rich proteins encoded by the VCS genes: rat PR-VB1, mouse MSG1, and human hPR-PB proteins) (Isemura et al. 1979; Courty et al. 1994; Isemura and Saitoh 1994). These proteins differ from other proline-rich proteins in the structure of their repeated peptide motif and in their low abundance of Glu/Gln amino acids. Here we show that the MSG1 protein also differs from classical proline-rich proteins in the regulation of its expression. Most pro-line-rich proteins in rodents are expressed at low basal levels in the SMGs and the parotids, and are induced by isoproterenol or tannin-rich diets (Ann et al. 1987; Mehansho et al. 1983,1987). Because of this induced expression and their capacity to bind polyphenols, such as tannins, it was proposed that these proline-rich proteins participate in dietary tannin detoxification. In contrast, the high expression levels of Vcs1 mRNA in the parotids is not dependent on any type of stimulation. In the rat, the VCSB1 gene was also shown to be constitutively expressed at high levels in the parotids (Rosinski-Chupin, unpublished data). This suggests that the VCS-encoded proline-rich proteins may have a function different from that of tannin-induced PRP. Indeed, on the basis of the presence of putative sites for collagenases, it was suggested that the VCS PRP may have a role in the protection of oral surfaces by interfering with bacterial collagenases (Courty et al. 1994). A PB-like polypeptide was also purified from bovine developing enamel (Strawich and Glimcher 1990), leading to the hypothesis that some of these proteins might also have a function in tooth mineralization (Isemura and Saitoh 1994).
No effect of androgens on the expression of the Vcs1 gene in the parotids has been detected. Whether this result is due to the lack of androgen responsive regulatory sequences in Vcs1 gene or to the absence of androgen responsiveness of the parotids is not known. To date, to our knowledge, no sex-linked difference of gene expression in the parotids has been demonstrated, and attempts to demonstrate the presence of the androgen receptor in mouse parotid glands have failed (Morrell et al. 1987).
In contrast, the mouseVcs2 gene is differentially expressed in the SMGs of males and females in response to androgens. Expression in males can be abolished by castration and expression in females can be induced by androgen injection. Therefore, the Vcs2 gene is reminiscent of the rat VCSA1 gene, although both genes differ in their level of expression. Whereas VCSA1 mRNAs represent more than 10% of adult male rat SMG mRNAs, Vcs2 transcripts are low-abundance mRNAs. In females, Vcs2 transcripts were not detected by Northern blot, RNase protection, or in situ hybridization. In contrast, low but detectable levels of VCSA1 mRNAs are present in female rat SMG. We also show that, like the rat VCSA1 gene (Rosinski-Chupin et al. 1993) the Vcs2 gene is expressed in the acinar cells of the SMG. This is in contrast to all other known androgen-regulated genes of mouse SMG, which are expressed only in the convoluted granular tubules of the glands. However, the presence of the androgen receptor in the acinar cells and in duct cells in mice and rats has been reported (Morrell et al. 1987; Sar et al. 1990). Therefore, the Vcs2 gene is the first potential target for regulation by the androgen receptor in mouse SMG acinar cells.
Cell specificity of expression among acinar cells is revealed because the level of Vcs2 transcript accumulation varied among cells. The reasons for such differences are not known, and could include different accessibility of cells to androgens or other hormones, and differences in the state of differentiation or in the cell cycle. The persistence of a perinatal cellular phenotype in some acinar cells of adult rat SMG has been reported (Man et al. 1995). The Vcs2 gene generates, by alternative splicing, at least five different mRNAs which can all be detected by one or the other of our two probes. On the basis of the cellular distribution of signals obtained with both probes, we have no evidence for a differential distribution of the mature transcripts. This may mean that this cellular specificity of expression is controlled at the level of the synthesis of the primary transcript, i.e., at the transcriptional level. An alternative hypothesis is a post-transcriptional regulation involving sequences common to the different transcripts.
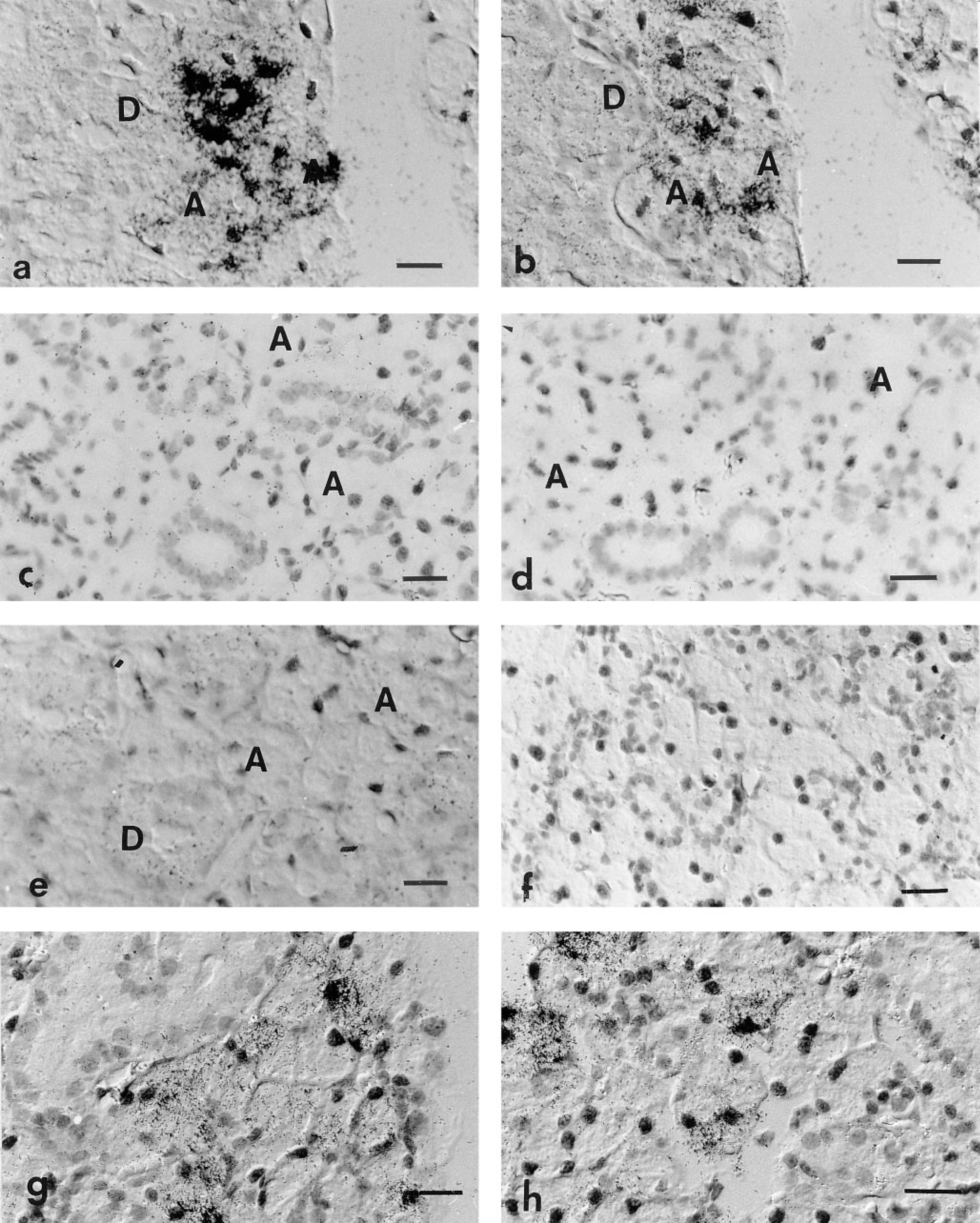
In situ hybridization experiments showing the dependence on androgens of the detection of Vcs2 transcripts. Sections of submandibular glands of male (
In conclusion, our results show that the VCS genes are differentially regulated, suggesting that regulation of expression has evolved together with the diversification of function, as has also been observed in a number of other multigene families (Ohta 1994). In addition, our study reveals for the first time the existence of sex-linked differences in gene expression in the acinar cells of the mouse SMG. This observation extends our previous demonstration that SMG acinar cells are target cells for androgen regulation and suggests that androgen responsiveness is a general characteristic of rodent SMG acinar cells.
Footnotes
Acknowledgements
Supported by grants from the Institut Pasteur, from the GREG (Groupement d'Etudes et de Recherches sur le Genome), and from the DRET (contract 93 113/DRET).
We especially thank Dr B. Laoide for critical reading of this manuscript and Dr C. Rougeot for help and suggestions in experimental care of mice.