
Abstract
Mucins are high molecular weight glycoproteins secreted by salivary glands and epithelial cells lining the digestive, respiratory, and reproductive tracts. These glyco-proteins, encoded in at least 13 distinct human genes, can be subdivided into gel-forming and membrane-associated forms. The gel-forming mucin MUC5B is secreted by mucous acinar cells in major and minor salivary glands, but little is known about the expression pattern of membrane-associated mucins. In this study, RT-PCR and Northern blotting demonstrated the presence of transcripts for MUC1 and MUC4 in both parotid and submandibular glands, and in situ hybridization localized these transcripts to epithelial cells lining striated and excretory ducts and in some serous acinar cells. The same cellular distribution was observed by immunohistochemistry. Soluble forms of both mucins were detected in parotid secretion after immunoprecipitation with mucin-specific antibodies. These studies have shown that membrane-associated mucins are produced in both parotid and submandibular glands and that they are expressed in different cell types than gel-forming mucins. Although the function of these mucins in the oral cavity remains to be elucidated, it is possible that they both contribute to the epithelial protective mucin layer and act as receptors initiating one or more intracellular signal transduction pathways.
M
Human submandibular, sublingual, and minor salivary gland secretions contain high and low molecular weight mucins named MG1 (mucous glycoprotein 1) and MG2 (mucous glycoprotein 2), respectively (Loomis et al. 1987). These mucins are important both for maintaining the viscoelastic and rheological properties of saliva and for binding and facilitating clearance of a variety of oral microorganisms (Tabak 1995). Cloning studies have shown that MG1 consists primarily of the MUC5B gene product (Nielsen et al. 1997; Troxler et al. 1997), whereas MG2 is encoded in the MUC7 gene (Bobek et al. 1993). MG1 is synthesized by mucous acinar cells (Nielsen et al. 1997). However, there is controversy as to whether MG2 is made in serous or mucous acini (Cohen et al. 1991; Nielsen et al. 1996, 1997; Khan et al. 1998).
Because the parotid gland is considered a purely serous gland on the basis of histological criteria, it has been widely assumed that this gland does not synthesize mucins. The parotid gland does not express MUC5B, the major component of MG1 (Troxler et al. 1995) or MUC7, the gene encoding MG2 (Biesbrock et al. 1997). Although MUC5B and MUC7 are both secreted soluble mucin gene products, the possibility remained that membrane-associated mucins could, in principle, be synthesized in the parotid gland, and this is supported by several earlier observations. For example, histochemical studies on parotid glands identified “neutral” mucins in serous acinar cells, in striated ducts, and in excretory ducts, while both neutral and “acid” mucins were identified in intercalated ducts (Harrison et al. 1987). Subsequently, simple mucin glycans were detected immunohistochemically in parotid serous acinar cells and duct cells (Therkildsen et al. 1994). Nevertheless, it was not possible to determine which mucin gene products were expressed solely on the basis of histochemical staining properties. Interestingly, several other studies demonstrated expression of “epithelial membrane antigen” in serous and duct cells of parotid gland (Tatemoto et al. 1987; Zotter et al. 1988a) and in adenolymphomas of this gland (Zotter et al. 1988b). Subsequent studies showed that this antigen was identical to the membrane-associated mucin MUC1. More recently, immunochemical evidence has suggested that yet another membrane-associated mucin, MUC4, is present in human parotid gland (Li et al. 2001; Weed et al. 2001). Taken together, these studies suggest that the parotid gland is a mucin-synthesizing exocrine gland and that at least two of the mucin gene products expressed are MUC1 and MUC4.
The present investigation was initiated to determine the expression patterns of membrane-associated mucins in the human parotid gland and to compare the expression pattern with that in human submandibular gland. MUC1 and MUC4 transcripts were identified in parotid gland by RT-PCR and by Northern blotting and were localized to epithelial cells lining striated and excretory ducts by in situ hybridization (ISH). MUC1 and MUC4 gene products displayed a similar cellular distribution by immunohistochemistry (IHC). Finally, immunoprecipitation (IP) experiments demonstrated that these mucins are components of parotid secretion.
Materials and Methods
Tissue Samples
Human parotid, submandibular, and sublingual gland tissue was obtained from the National Disease Research Interchange (Philadelphia, PA). All samples were snap-frozen in liquid nitrogen less than 12 hr post mortem, shipped on dry ice, and stored at −80C until used. Tissue samples used for ISH and IHC were thawed on ice, fixed in 4% paraformaldehyde at 4C overnight, embedded in paraffin, sectioned, and mounted on microscope slides. Buccal epithelial cells were obtained from whole saliva sediment as described (Yao et al. 2000). These studies were approved by the Institutional Review Board at Boston University Medical Center.
PCR and Northern Blotting Analyses
RNA was isolated from tissue samples using TriPure Reagent (Roche; Indianapolis, IN). RT-PCR reactions were carried out under standard conditions (30 cycles of 1 min at 95C, 1 min at 62C, 1.5 min at 72C) using primers corresponding to non-tandem repeat regions of the membrane-associated mucins MUC1, MUC3, MUC4, MUC12, and the gel-forming mucin MUC5B (Table 1). Northern blots of salivary gland RNAs (15 μg) were hybridized with 32P-labeled MUC1 and MUC4 probes containing mucin tandem repeat sequences. The 0.4-kb MUC1 probe was derived from a clone isolated from a human gallbladder cDNA library (Keates et al. 1997) and the 0.8-kb MUC4 probe was a cloned RT-PCR product derived from human trachea RNA (Liu et al. 1998). Hybridization and washes were carried out as previously described (Keates et al. 1997).
In Situ Hybridization
MUC1 and MUC4 riboprobes were synthesized using the Riboprobe In Vitro Transcription System (Promega; Madison, WI) according to the manufacturer's instructions. pBluescript plasmids containing the MUC1 and MUC4 tandem repeat probes used for Northern analysis (see above) were linearized with BamH1 and antisense riboprobes were synthesized using T7 RNA polymerase. The corresponding sense riboprobes were synthesized from plasmids linearized with XhoI using T3 RNA polymerase. Riboprobes (labeled with [α-35S]-UTP) were resuspended in 20 μl of 1 M DTT and 180 μl of hybridization buffer (HB) containing 20 mM Tris-HCl, pH 7.4, 5 mM EDTA, 10 mM Na2HPO4, 50% deionized formamide, 300 mM NaCl, 1 × Denhardt's solution, 10% dextran sulfate, and 0.5 mg/ml of total yeast RNA.
Primers used in RT-PCR experiments

Tissue sections on glass microscope slides were deparaffinized in xylene, rehydrated using a graded ethanol series (100% to 30%), rinsed in 150 mM NaCl and PBS, each for 5 min, and postfixed in 4% paraformaldehyde in PBS for 30 min. Tissue sections were treated with proteinase K (Roche; 20 μg/ml) in 50 mM Tris-HCl, pH 7.2, containing 5 mM EDTA at room temperature (RT) for 7.5 min and fixed in 4% paraformaldehyde in PBS. Acetylation was carried out for 10 min with 0.25% acetic anhydride in 100 mM triethanolamine, pH 8.0, and slides were rinsed sequentially in PBS and 150 mM NaCl, dehydrated through a graded ethanol series, and allowed to dry for at least 2 hr before hybridization.
Hybridization was carried out as previously described (Toselli et al. 1992). Briefly, 35S-labeled riboprobes were adjusted to 25,000 cpm/μl with HB before use. Probes were applied directly to tissue sections and the sections were covered with coverslips (22 × 22 mm, 30 μl of probe/slip). Hybridization was carried out at 52C for 16 hr in a humid chamber. Slides were rinsed in 5 × SSC containing 10 mM DTT at 50C for 30 min and subsequently at 65C for 20 min in 2 × SSC containing 50% formamide and 10 mM DTT. Slides were rinsed twice at 37C for 10 min in STE (10 mM Tris-HCl, pH 7.5, 400 mM NaCl, and 5 mM EDTA). Sections were treated with RNase A (20 μg/ml) in STE at 37C for 45 min and rinsed with STE for 5 min, with 2 × SSC for 15 min, and with 0.1 × SSC for 15 min, each at 37C. Sections were dehydrated in an ethanol series (30–95%) containing 0.3 M ammonium acetate and finally in 100% ethanol. Slides were processed for standard light microscopic autoradiography using Kodak NTB-2 nuclear track emulsion and placed in a desiccation chamber for 1–2 weeks at 4C. The slides were developed in Kodak D-19 developer, stained with hematoxylin and eosin, and examined using a Leitz Dialux 20 microscope (Wetzlar, Germany). At least 40 sections from four different parotid gland and four different submandibular gland samples were examined.
Immunohistochemistry
Sections were deparaffinized and rehydrated as described above and endogenous peroxidase activity was quenched by treatment with 0.3% H2O2 in 100% methanol for 30 min at RT. Antigen retrieval was carried out by autoclaving (liquid cycle) the slides in 10 mM citrate buffer, pH 6.0, for 15 min and subsequently allowing slides to stand at RT for 20 min.
IHC was carried out with a series of specific antibodies directed against MUC1 and MUC4 (Table 2). MUC1 was detected using a monoclonal anti-MUC1 antibody (clone E29) purchased from NeoMarkers (Fremont, CA) and MUC4 was detected using rabbit polyclonal antibodies SG9 and MUC4-CT. SG9 (kindly provided by Dr. Sandra Gendler, Mayo Clinic Scottsdale) is directed against a synthetic peptide corresponding to a sequence in the tandem repeat domain and MUC4-CT is directed against a synthetic peptide, SGARF-SYFLNSAEAL, corresponding to a sequence in the cytoplasmic domain. A similar antibody directed against an analogous portion of the cytoplasmic domain of the rat homologue (i.e., ASGP-2 subunit of SMC) of human MUC4 has been used previously to detect the membrane-associated form of SMC in a variety of tissues (Rossi et al. 1996).
Antibodies used for immunohistochemical and biochemical analyses a
IHC, immunohistochemistry; WB, Western blot; TR, tandem repeat.
Tissue sections mounted on glass slides were incubated with blocking buffer for 30 min at 37C to mask nonspecific binding sites. For all antibodies, the blocking buffer was PBS containing 3% BSA and 10% normal goat serum. Slides were rinsed three times with PBS containing 0.1% Tween-20 for 5 min and incubated with primary antibodies for 30 min at 37C. E29 (1:100), SG9 (1:100), and MUC4-CT (1:500) were diluted in PBS, 1% BSA, and 3% normal goat serum. Sections were rinsed and incubated with either biotinylated goat anti-mouse IgG (1:1000) or biotinylated goat anti-rabbit IgG (1:200) at 37C for 30 min. Slides were rinsed three times and incubated at 37C for 30 min with ABC reagent (avidin/biotin complex Elite Vectastain; Vector Laboratories, Burlingame, CA) in PBS containing 0.1% Tween-20 to enhance the color reaction. Color development was achieved using diaminobenzidine (Vector Laboratories) for 2–10 min at RT and sections were stained with 0.68% hematoxylin before examination under the light microscope. Control experiments were carried out in which incubation with primary antibodies was omitted. Sections from four different parotid gland and four different submandibular gland specimens were examined.
Immunoprecipitation and Western Blotting
Stimulated parotid secretion was collected using Curby cups as described (Jensen et al. 1992). Samples were dialyzed, lyophilized, and parotid saliva protein was dissolved at a concentration of 20 mg/ml in IP buffer (50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1% NP-40, 0.5% deoxycholate, 0.1% SDS) containing protease inhibitors (Complete Inhibitor Mixture; Sigma, St Louis, MO). Aliquots of parotid saliva protein (100 μl) were incubated with primary antibodies overnight at 4C on a rotary mixer. The primary antibodies used are listed in Table 2. IP experiments were performed with the anti-MUC1 antibodies HMFG-2 (25 μl) and CT-2 (25 μl) and with anti-MG1 antiserum (10 μl). HMFG-2 and CT-2 (both kindly provided by Dr. Sandra Gendler) are directed against synthetic peptides corresponding to sequences in the tandem repeat and cytoplasmic domains of MUC1, respectively. The anti-MG1 antibody is a rabbit polyclonal antibody directed against high molecular weight mucins isolated from submandibular/sublingual secretion (Rayment et al. 2000) and recognizes both MUC4 and MUC5B (Liu et al. 1998). This antibody was used in the present study to immunoprecipitate MUC4 from parotid secretion because MUC5B was not found to be expressed in parotid gland tissue by both RT-PCR (Figure 1) and by ISH (Figure 3).
Protein A/G beads (Santa Cruz Biotechnology; Santa Cruz, CA) were added and the mixture was incubated for 1 hr at 4C on a rotary mixer. Beads were collected by centrifugation, washed twice in IP buffer and twice in wash buffer containing 20 mM Tris-HCl, pH 8.0, 1 mM EDTA, 900 mM NaCl, and 0.5% NP-40. Bound proteins eluted by boiling in sample loading buffer were electrophoresed on 6% polyacrylamide gels or on tricine gels, transferred to nitrocellulose, and the blots incubated as described below. To detect MUC1, blots containing HMFG-2 or CT-2 IPs were probed with the same antibodies. To detect MUC4, blots containing anti-MG1 IPs were incubated with either the anti-MG1 antibody (1:2500) or with the MUC4-specific antibody SG9 (1:200). Blots were incubated with peroxidase-conjugated species-specific secondary antibodies with detection using the Renaissance chemiluminescence reagent (NEN; Boston, MA).
Results
On the basis of previous reports suggesting that mucins might be synthesized in human parotid gland, the present work was undertaken to determine whether genes for membrane-associated mucins were expressed in this gland and to compare the pattern of mucin gene expression in parotid and submandibular glands.
RT-PCR and Northern Analyses
Expression of mucin genes in major salivary glands was investigated by RT-PCR. Parotid gland contained transcripts for the membrane-associated mucins MUC1 and MUC4 (Figure 1, Lane P), whereas transcripts for MUC3 and MUC12 were not detectable (data not shown). Transcripts for the gel-forming mucin MUC5B were also not detectable in parotid gland using this technique (Figure 1, Lane P). In contrast, submandibular and sublingual gland contained transcripts for all three mucins (Figure 1, Lanes SM, SL). Interestingly, buccal epithelial cells also contained MUC1 and MUC4 transcripts with lower but detectable levels of MUC5B transcripts (Figure 1, Lane BEC).
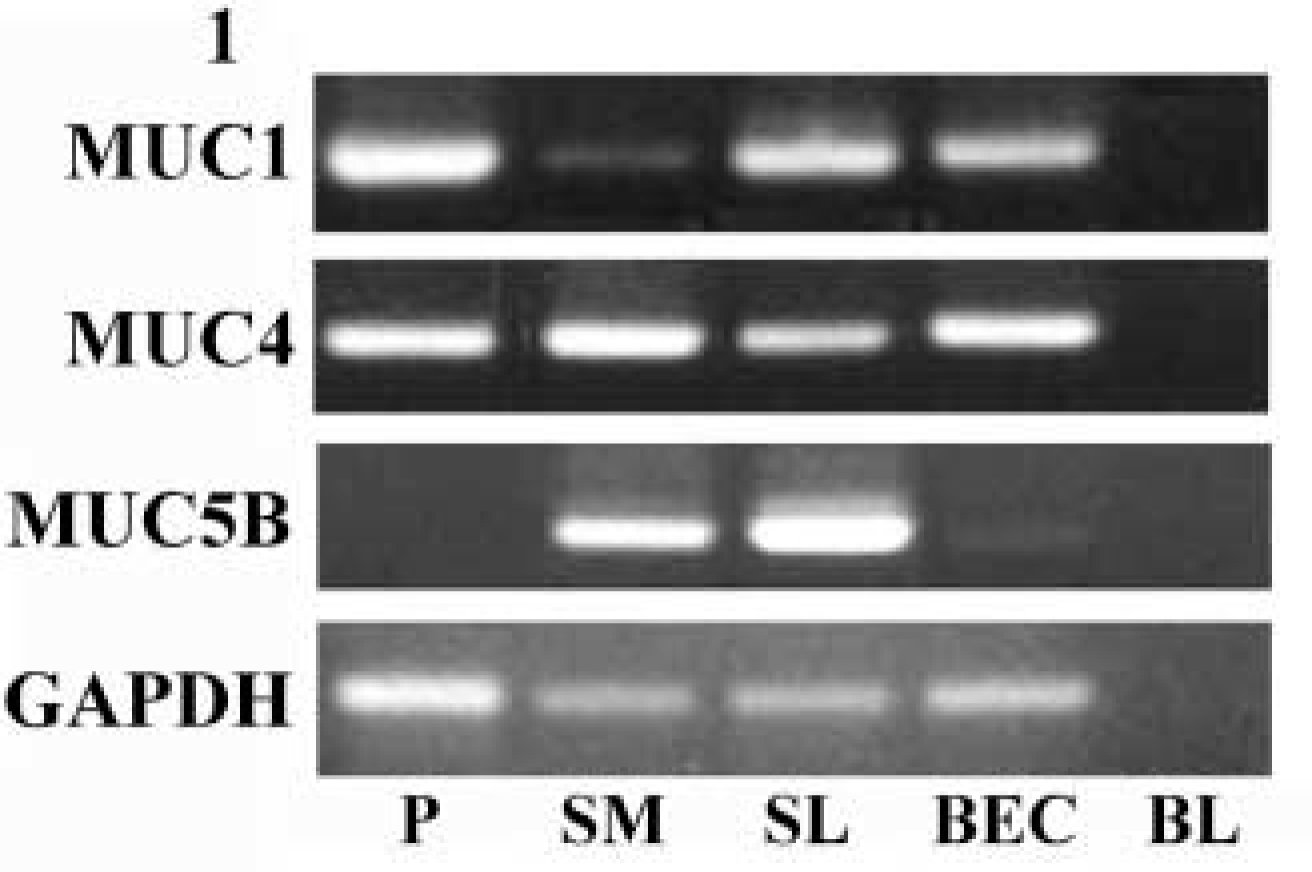
Analysis of mucin gene expression in salivary glands by RT-PCR. RNA from parotid (P), submandibular (SM), and sublingual (SL) glands and from buccal epithelial cells (BEC) was reverse transcribed and the resulting cDNAs were used in PCR reactions with the primers given in Table 1. PCR products were electrophoresed on 1% agarose gels and visualized with ethidium bromide. Control reactions were carried out to assess transcript levels of the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and blank reactions (BL) contained no added cDNA template.
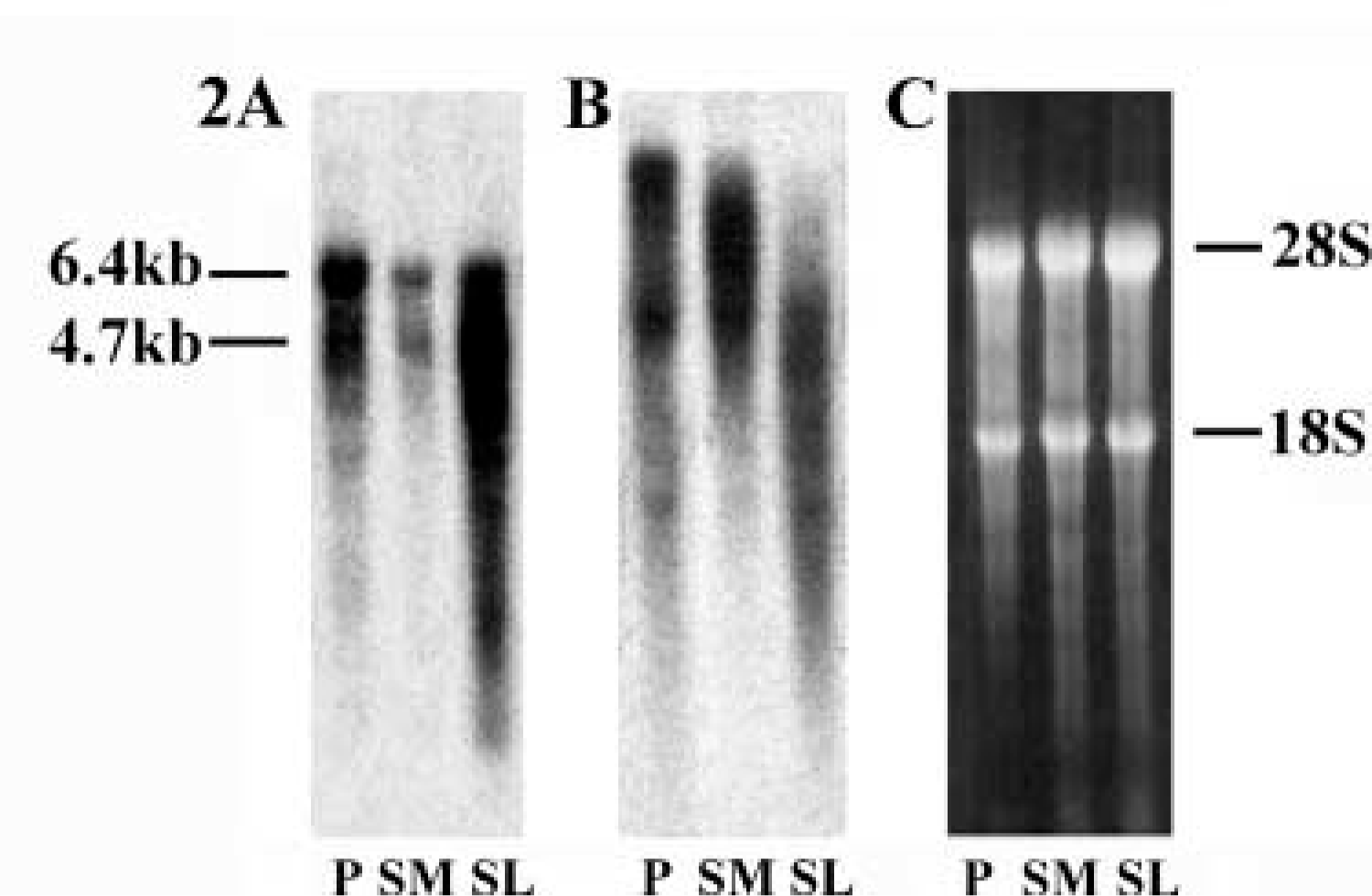
Analysis of mucin gene expression in salivary glands on Northern blots. RNA from parotid (P), submandibular (SM), and sublingual (SL) gland was electrophoresed on 1% agarose denaturing gels, blotted onto nylon membrane, and blots were hybridized with probes for MUC1 (
To determine the relative expression levels of MUC1 and MUC4 transcripts in parotid, submandibular, and sublingual glands, Northern blots of RNA from these glands were hybridized with DNA probes containing MUC1 and MUC4 tandem repeats. Two discrete MUC1 transcripts of approximately 6.4 and 4.7 kb were observed in all three major salivary glands (Figure 2A). The 6.4-kb transcript most likely represents full-length MUC1 RNA, while the 4.7-kb transcript may correspond to an allelic variation containing a lesser number of tandem repeats (Gendler et al. 1990). Similarly, MUC4 transcripts were abundant in all three major salivary glands (Figure 2B). The largest transcripts in parotid and submandibular gland were approximately 25 kb, whereas the largest transcripts in sublingual gland were in the 10–15-kb range. Because the RNAs analyzed came from glandular tissue obtained from different individuals, the variability in size of MUC4 transcripts most likely reflects polymorphism in the number of tandem repeats (Moniaux et al. 1999). It should be noted that the polydisperse hybridization signals in Figure 2 are typical of those for mucin mRNAs reported by us and others (Strous and Dekker 1992; Gum 1992; Keates et al. 1997; Troxler et al. 1997). Although molecular basis for this polydispersity is not understood, it has been suggested that it may be due to a preferential turnover of extremely long mucin mRNAs (Debailleul et al. 1998). The RNA samples from each gland appeared to be largely intact by ethidium bromide staining (Figure 2C), and therefore the polydisperse hybridization signals observed are probably not the result of RNA degradation. These results demonstrate that the mucin transcripts identified by RT-PCR in parotid gland arise from abundant mucin mRNAs.
In Situ Hybridization
Having identified MUC1 and MUC4 transcripts in major salivary glands (Figures 2A and 2B), ISH was carried out to localize these mRNAs within glandular tissue. The data obtained with antisense riboprobes are shown in Figures 3A, 3C, 3E, 3G, 3I, 3K, 3M, and 3O, and those with sense probes (negative controls) are shown in Figures 3B, 3D, 3F, 3H, 3J, 3L, 3N, and 3P. In parotid gland, weak hybridization of the MUC1 antisense probe was detected in cells lining striated ducts (Figure 3A), whereas in submandibular gland a much stronger signal was observed in striated duct cells as well as in some serous acinar cells (Figure 3C). A similar distribution of mucin transcripts was observed in both glands with the MUC4 antisense probe (Figures 3E and 3G), while the intensity of hybridization in both glands was nearly the same. The MUC4 antisense probe also hybridized strongly to cells lining major excretory ducts in both parotid (Figure 3I) and submandibular gland (Figure 3K). As expected, ISH carried out with the MUC5B antisense probe showed no significant hybridization to transcripts in parotid gland acinar cells (Figure 3M), although transcripts were clearly present in mucous acinar cells of submandibular gland (panel O). No MUC5B transcripts were detected in duct cells in either salivary gland (Figures 3M and 3O). In addition, no significant hybridization signal was detected with the sense riboprobes for any of the mucin genes examined.
Immunohistochemistry
To determine whether MUC1 and MUC4 proteins were expressed in salivary glands, IHC experiments were performed using a monoclonal antibody directed against an epitope in the tandem repeat domain of MUC1 (E29) and polyclonal antibodies directed against pep-tide sequences in both the tandem repeat (SG9) and cytoplasmic (MUC4-CT) domains of MUC4.
Using the anti-MUC1 antibody, strong immunoreactivity was detected in epithelial cells lining striated ducts in parotid and submandibular gland (Figures 4A and 4B). Immunoreactive anti-MUC1 material was also observed in the lumen of striated ducts and diffuse staining was detected in some clusters of serous acinar cells in both glands. In other sections, immunoreactive MUC1 material was also detected in major excretory ducts in both parotid and submandibular glands (data not shown). Both anti-MUC4 antibodies reacted strongly with material in striated ductal epithelial cells of both parotid (Figures 4C and 4E) and submandibular (Figures 4D and 4F) gland. Some immunoreactive material was also observed in serous acinar cells in both glands. The similarity in the pattern of immunochemical staining observed with SG9 (extracellular domain; Figures 4C and 4D) and MUC4-CT (cytoplasmic domain; Figures 4E and 4F) indicated that the MUC4 present on duct epithelial cells and serous acinar cells represents intact mucin molecules containing the tandem repeat, transmembrane, and cytoplasmic regions. Control experiments carried out in which each of the primary antibodies was omitted did not demonstrate immunoreactive protein in either parotid or submandibular gland sections (data not shown).
Immunoprecipitation and Western Blotting
To determine whether MUC1 and MUC4 proteins are normal constituents of parotid secretion, a series of IP experiments were carried out. Western blots of material precipitated with HMFG-2, an antibody directed against the MUC1 tandem repeat domain, and probed with the same antibody revealed a faint smear of high molecular weight protein in the upper portion of the separating gel. This material had a similar electrophoretic mobility as authentic MUC1 in IPs from a lysate of MCF-7 breast cancer cells probed with the same antibody (Figure 5A, Lanes 1, 2). Control IPs were prepared using an irrelevant monoclonal antibody (anti-glutathione-S-transferase; GST), and when blots were probed with HMFG-2 (against MUC1 tandem repeats) no immunoreactive material was detected (Figure 5A, Lane 3). This demonstrates that the extracellular portion of MUC1 is present in parotid secretion. Western blots of anti-MG1 IPs, probed with the same antibody, revealed several discrete protein bands all greater than 200 kD (Figure 5A, Lane 4). A similar pattern of immunoreactive proteins was observed when blots of anti-MG1 IPs were probed with SG9, which recognizes an epitope in the tandem repeat domain of MUC4 (Figure 5A, Lane 5). This demonstrates that the extracellular portion of MUC4 was also detectable in parotid secretion, and we concluded from these studies that MUC1 and MUC4 proteins are normal constituents of this fluid.
Because MUC1 is a transmembrane protein containing both a mucin-like extracellular domain and a short cytoplasmic domain, the immunoprecipitation experiment described above does not distinguish between intact MUC1 (containing the cytoplasmic domain) released into parotid secretion by turnover of duct epithelial cells and soluble MUC1 (lacking the cytoplasmic domain) released into parotid secretion by proteolytic cleavage of the membrane-associated mucin. Therefore, additional IP experiments were carried out using CT-2, an antibody directed against a synthetic peptide corresponding to a sequence in the cytoplasmic domain of MUC1. IPs of parotid secretion prepared with CT-2 revealed no detectable immunoreactive protein when blots of IPs were probed with the same antibody (Figure 5B, Lane 1). Control IPs of MCF-7 breast cancer cell lysates probed with CT-2 contained three major proteins in the 18–24-kD range (Figure 5B, Lane 2). It has been suggested that these immunoreactive bands represent different glycoforms of the membrane-associated MUC1 subunit which contains the cytoplasmic domain (Schroeder et al. 2001). The absence of these bands in IPs of parotid secretion, coupled with the presence of HMFG-2 immunoreactive material in parotid secretion (Figure 5A, Lane 1), provides strong evidence that the MUC1 in this secretion most likely results from proteolytic cleavage of membrane-associated mucin rather than from turnover of duct epithelial cells.
Discussion
The structures and properties of salivary mucins have been studied extensively during the past two decades. These proteins were first identified as biochemical entities MG1 and MG2 and later were found to represent the products of the MUC5B and MUC7 genes, respectively (Bobek et al. 1993; Nielsen et al. 1997; Troxler et al. 1997). Expression of MUC5B (i.e., MG1) has been localized by ISH to mucous acinar cells in submandibular, sublingual, and minor salivary glands (Nielsen et al. 1997). To date, MUC5B has been considered the principal high molecular weight gel-forming mucin in salivary secretions, and investigations into the expression of membrane-associated mucins in salivary glands have been limited.
Early histochemical studies identified a mucin-like substance in serous acini and duct epithelial cells in both parotid and submandibular glands (Harrison et al. 1987), and antibodies directed against an abundant “epithelial membrane antigen” recognized protein with a similar cellular distribution in both glands (Zotter et al. 1988a). It was only after MUC1 was cloned independently by two different groups (Gendler et al. 1990; Lan et al. 1990) that it became known that the epithelial membrane antigen identified by histochemical and immunological criteria was synonymous with MUC1. This mucin was subsequently identified in submandibular gland by IHC (Ho et al. 1993) and RT-PCR (Biesbrock et al. 1997) and in minor salivary glands by ISH (Sengupta et al. 2001). Less is known about the expression of MUC4 in salivary glands. Although MUC4 was first cloned from a trachea cDNA expression library a decade ago (Porchet et al. 1991), the complete nucleotide and deduced amino acid sequence have been reported only recently (Moniaux et al. 1999). Furthermore, until the putative membrane-spanning and cytoplasmic domains were identified, it was not recognized that MUC4 was indeed a membrane-associated mucin. Nevertheless, we and others have provided preliminary evidence that MUC4 transcripts occur in submandibular and sublingual glands (Biesbrock et al. 1997; Troxler et al. 1997; Liu et al. 1998). In addition, we have isolated MUC4 clones from a submandibular gland cDNA library screened with a DNA probe containing MUC4 tandem repeats (Liu et al. 1998). Finally, an antibody directed against MG1 in submandibular/sublingual secretion was found to react with a GST-MUC4 fusion protein, providing unmistakable evidence that MUC4 is a component of this secretion (Liu et al. 1998).
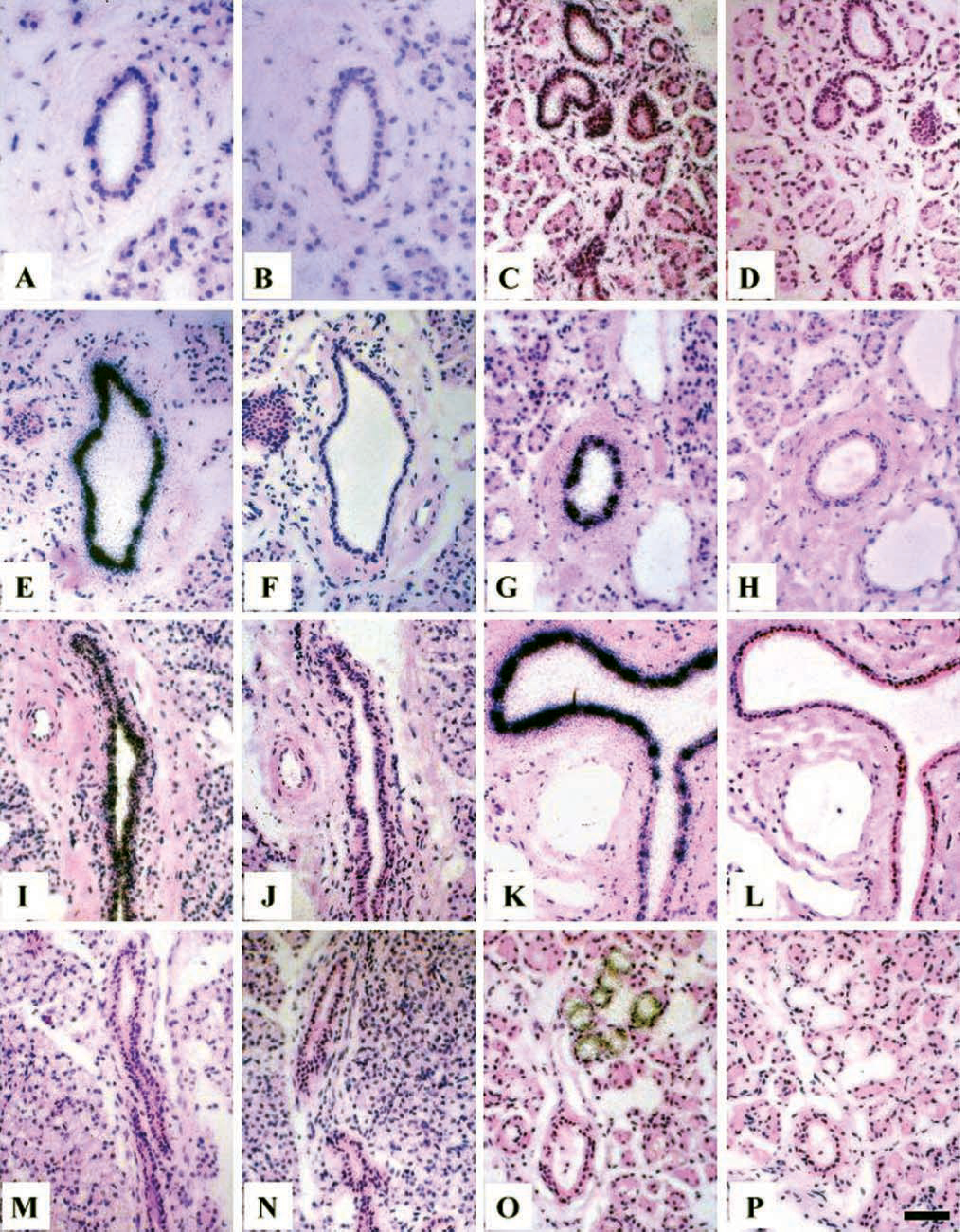
Analysis of mucin gene expression in parotid and submandibular gland by ISH. Tissue sections were hybridized with MUC1, MUC4, and MUC5B tandem repeat riboprobes as described in Materials and Methods. After development slides were stained with hematoxylin and eosin and examined in the light microscope. Parotid gland hybridized with MUC1 antisense (
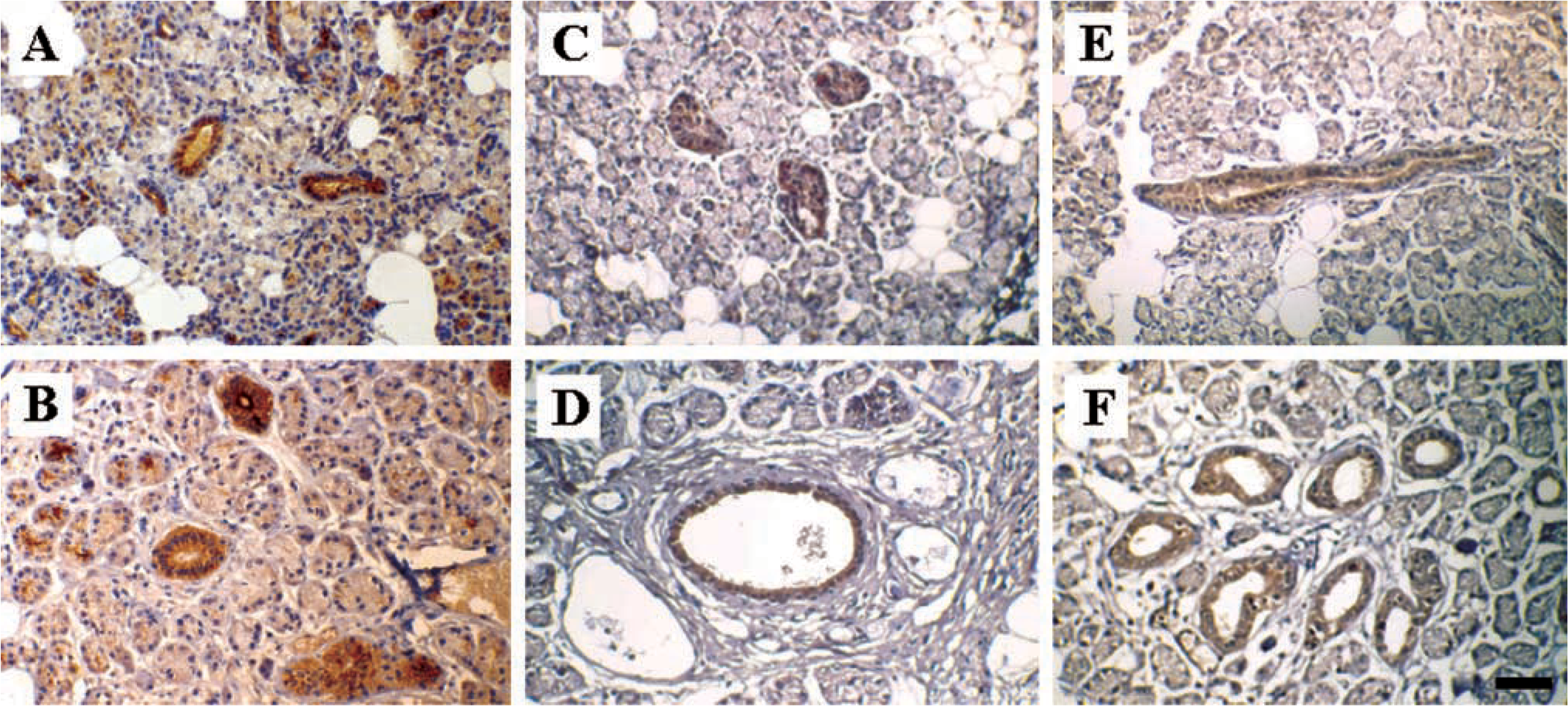
Analysis of mucin protein expression in parotid and submandibular gland by IHC. Parotid gland sections were probed with E29 to detect MUC1 (
In view of earlier evidence summarized above that epithelial membrane antigen occurred in both parotid and submandibular glands, it seemed likely that the membrane-associated mucins MUC1 and MUC4 would also be expressed in parotid gland. In a preliminary communication, we reported that transcripts for MUC1 and MUC4, as well as the corresponding proteins, were synthesized in parotid gland (Offner et al. 2001). More recently, MUC4 expression has been reported in parotid gland on the basis of IHC and Western blotting (Li et al. 2001; Weed et al. 2001). The present investigation is the first to systematically compare the expression patterns of the membrane-associated mucins in major human salivary glands. RT-PCR and Northern blotting studies have shown that both parotid and submandibular glands produce MUC1 and MUC4 transcripts (but not MUC3 and MUC12), and these have been localized primarily to cells lining striated ducts and major excretory ducts, with some expression in serous acinar cells. Immunohistochemical localization confirmed these findings and showed that MUC1 and MUC4 proteins had the same cellular distribution as did the mRNAs for these mucins.
After having identified both MUC1 and MUC4 transcripts and proteins in parotid gland tissue, we were interested to determine whether these mucins were present in parotid secretion and, if so, to ascertain whether they were soluble forms arising by proteolytic cleavage of membrane-associated forms or were full-length species arising from turnover of duct epithelial cells. It was found that both MUC1 and MUC4 could be immunoprecipitated from parotid secretion with antibodies directed against extracellular portions of the respective molecules (see Figure 5). However, MUC1 could not be detected in immunoprecipitates using CT-2, an antibody directed against the cytoplasmic domain of this mucin. This provides strong evidence that MUC1 in parotid secretion is probably derived proteolytically from the membrane-associated form. The origin of MUC4 in parotid secretion remains to be determined.
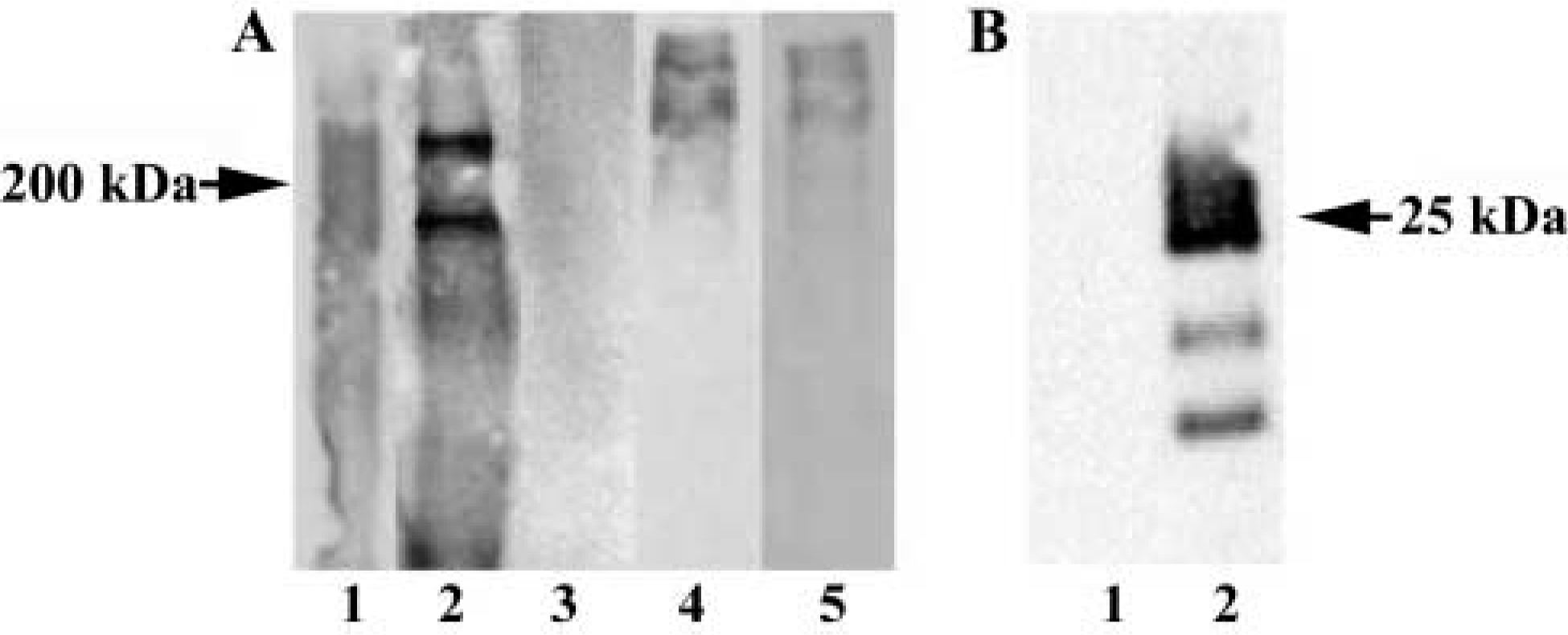
Analysis of mucin protein expression in parotid secretion by IP and Western blotting. The antibodies used are described in Table 2 and experimental details are given in Materials and Methods. (
The physiological function of MUC1 and MUC4 in salivary gland duct cells is not known. By analogy to breast tissue, it has been suggested that in minor salivary glands MUC1 may facilitate flow of secretion through the duct system (Sengupta et al. 2001). A somewhat more attractive alternative might be that the cytoplasmic domain of these mucins may, in some way, be involved in signal transduction. For example, the cytoplasmic domain of MUC1 contains several tyrosine residues that can be phosphorylated. In transgenic mice overexpressing human MUC1, phosphorylation of tyrosine residues in the cytoplasmic domain in response to EGFR ligands resulted in activation of ERK1/2 in the MAPK signal transduction pathway (Schroeder et al. 2001). In addition, phosphorylation of tyrosine residues in a chimeric CD8-MUC1 construct in response to exogenous CD8 was also found to activate ERK2 (Meerzaman et al. 2001). Recently, it has been reported that the rat homologue of MUC4 binds to the receptor tyrosine kinase ErbB2, resulting in activation of the kinase that initiates signaling through MAPK and PI3K (Ram et al. 2000). Based on the foregoing and demonstration of MUC1 and MUC4 in human parotid and submandibular gland (see Figures 3 and 4) these membrane-associated mucins may play a critical role in signaling processes related to the normal physiologic functions of these glands.
In addition, it is possible that MUC1 and MUC4, synthesized in duct epithelial cells, are released proteolytically and enter the oral cavity, where they interact with one or more receptor-like molecules on oral epithelial surfaces. The resulting membrane-associated mucin complexes could interact with the gel-forming mucin MUC5B and form a “scaffold” (Offner and Troxler 2000). Evidence for the formation of such a scaffold on an epithelial surface derives from a study with muc1-null mice and mice lacking the cystic fibrosis transmembrane conductance regulator (CFTR) gene (Parmley and Gendler 1998). Homozygous CFTR-null mice produced excessive amounts of PAS-positive material (i.e., mucus) in the lumen of the small bowel. However, when CFTR-null mice were crossed with muc1-null mice, double-knockout mice exhibited a dramatic reduction in the quantity of mucus in the intestinal lumen. These results were interpreted to mean that muc1 in the CFTR-null mice formed complexes with the gel-forming mucins, resulting in a mucin scaffold. In the double-knockout mice lacking muc1, such a scaffold could not form. Finally, these results suggest that in humans, MUC1 might form complexes with the gel-forming mucin MUC5B, and the resulting scaffold on oral epithelial surfaces could protect against mechanical injury, pathogenic bacteria, and other noxious agents.
Footnotes
Acknowledgments
Supported in part by NIH grants DK 44619, DE 11691, and DE 07652.
We wish to thank Dr Sandra Gendler (Mayo Clinic; Scottsdale, AZ) for providing antibodies to MUC1 and MUC4.