
Abstract
The mechanism of hemopoietic stem cell homing to the bone marrow involves molecular interactions that mediate the recognition and interaction of these cells with the marrow microenvironment, including the extracellular matrix. On selective binding, this environment, in combination with soluble cytokines, regulates stem cell proliferation and differentiation. Using immunofluorescence labeling, we analyzed the location of the prominent extracellular matrix proteins fibronectin, collagen Types I, III, and IV, and laminin in sections of murine femoral bone marrow. Collagen Types I, IV, and fibronectin were localized to the endosteum, the region of the femoral microenvironment for which homing stem cells have a high affinity. The results further demonstrated a strong spatial association of collagen Type IV and laminin with the bone marrow vessels, including arterioles, veins, and sinuses. Fibronectin was distributed throughout the central marrow region, and all the proteins analyzed except collagen Type III were present in the bone, although at different levels. Fibronectin, collagen Types III and IV, and laminin were also present in the periosteum. The distinct locations of particular extracellular matrix proteins support the notion that they may play an important mechanistic role in the homing of engrafting cells.
T
The bone marrow microenvironment is a complex three-dimensional structure composed of many cell types and abundant extracellular matrix (ECM) proteins. These proteins include fibronectin, collagen Types I, III, and IV, laminin, thrompospondin, hemonectin, and proteoglycans. Much of the data regarding ECM was derived through the analysis of the adherent layer of long-term bone marrow cultures (LTBMC) (Dexter et al. 1977). These have been shown to contain many ECM components including fibronectin (Singer et al. 1985), collagen Types I, III, IV, and V (Keating et al. 1982), laminin, and various proteoglycans (Zuckerman and Wicha 1983).
The establishment of in vitro cultures permitted detailed analysis of the relationships between hemopoietic cells and the microenvironment. Many studies have since provided strong evidence of the importance of the bone marrow microenvironment in the development and regulation of hemopoietic cells (Quesenberry et al. 1987; Campbell and Wicha 1988). In murine LTBMC, the deposition of ECM proteins in the adherent layer was shown to coincide with the onset of hemopoietic cell production (Zuckerman and Wicha 1983). In addition, close binding interactions with hemopoietic stem cells and the adherent layer are believed to facilitate the stromal cells’ ability to support hemopoietic growth (Dexter et al. 1977; Gordon 1988). In human LTBMC, most primitive hemopoietic progenitors preferentially reside within the adherent layer (Coulombel et al. 1983), but discrete hemopoietic foci develop across the surface of this complex stromal layer (Cutinho et al. 1993), possibly reflecting restriction of spatial development to particular stromal areas (Zipori 1989).
The existence of hemopoietic “niches” composed of microenvironmental components has been hypothesized for many years. Supporting evidence comes from studies showing that thrombospondin is a cytoadhesion molecule for human hemopoietic progenitor cells in vitro (Long and Dixit 1990). Long et al. (1992) extended this study, showing that the cytokine thrombospondin and the ECM molecule c-kit ligand function as a signal complex regulating hemopoietic stem cell development in vitro. Co-immobilization of these two proteins identified a progenitor population requiring adhesion to both ligands to form colonies. Recently, Bruno et al. (1995) showed that the human marrow ECM component heparan sulfate proteoglycan mediates the adhesion of hemopoietic progenitor cells to cytokines in vitro.
However, because of the diffuse three-dimensional nature of its structure and technical limitations, analysis of the marrow microenvironment in vivo has been severely hampered. As a consequence, the precise location of specific ECM proteins and hence their possible regulatory influences on various populations of hemopoietic cells, including a potential role in the hemopoietic stem cell “niche,” remains poorly understood. Such analysis is critical so that the relevance of in vitro observations can be evaluated. We have mapped five key bone marrow ECM proteins within murine whole-bone marrow sections and have shown spatial localization to different regions. Correlated with accumulating evidence of the spatial localization of different hemopoietic populations in vivo (e.g., hemopoietic stem cells in the endosteal region), these findings give us a better understanding of the complex interactions by which hemopoiesis is regulated.
Materials and Methods
Mice
Balb/c mice were purchased from Charles River Laboratories, (Wilmington, MA) and were housed in a conventional clean facility for at least a week before experimental use. All mice received mouse chow and acidified water ad libitum. All animal protocols were approved by the institute's animal research committee and experiments were carried out under its guidelines for the care and use of laboratory animals.
Femoral Sections
Sections of paraffin-embedded whole murine femurs were made as previously described (Nilsson et al. 1996). Briefly, mice were perfused with 4% paraformaldehyde into the descending aorta. Femurs were removed, decalcified, and embedded in Paraplast x-tra (Oxford Labware; St Louis, MO).
Five-μm femoral sections were mounted on 0.01% poly-
Analysis
The expression of ECM was analyzed in multiple sections from at least three different mice. In addition, analysis was done by multiple observers, with the levels of expression in different areas being graded within rigid guidelines. Where the level of expression was abundant, it was described as bright. Where the protein was obviously expressed it was described as moderate. When the overall levels of expression were low it was described as faint, and where the area was clearly negative it was described as absent.
Results
Using fluorescence microscopy, we have mapped the location of the key extracellular matrix proteins fibronectin, collagen Types I, III, and IV, and laminin in paraffin-embedded whole femoral sections (Figure 1; Table 1).
Distribution of Fibronectin
Very high levels of fibronectin were detected in the periosteal region of the femur, in the bone itself, and in the endosteal region (Figure 1A). In addition, fibronectin was present in megakaryocytes and was dispersed throughout the central marrow region of the femur (Figure 1B). No specific fibronectin labeling was observed around the marrow vessels, including the arterioles, veins, and central sinuses.
Distribution of Collagens Type I, III, and IV
Collagen Type I was not detected in the periosteal region (Figures 1C and 1D). In contrast, very high levels of collagen Type I were detected in both the compact bone as well as the bone trabeculae. Collagen Type I was also detected in the endosteum, although the amount of positivity varied greatly, with some areas having very high levels of expression and others having much lower levels. No collagen Type I was detected in the central marrow region, including around the marrow vessels.
Collagen Type III was clearly present in the peri-osteal region of the femur but was undetectable in the bone itself and in the endosteal region (Figure 1E). Similar to collagen Type I, no collagen Type III was detected in the central marrow region, including around the marrow vessels.
Collagen Type IV was clearly present in the peri-osteal region, with very low amounts in the bone itself (Figure 1F). The pattern of collagen Type IV positivity in the endosteal region appeared to be discontinuous, but there was definite positivity located along this region. In the central marrow region, collagen Type IV was present around the marrow vessels, including the arterioles, veins, and central sinuses (Figure 1G).
Distribution of Laminin
Similar to fibronectin, very high levels of laminin were detected in the periosteal region of the femur, but with very little in the bone itself (Figure 1H). No laminin was detected in the endosteal region. High levels of laminin were detected in the walls lining the marrow arteries, whereas laminin was discontinuous along the walls of marrow sinusoids (Figure 1I).
Analysis with either rabbit or goat isotype control and secondary antibody revealed no background or nonspecific fluorescence in any of the marrow or bone areas analyzed (Figures 1J and 1K).
Discussion
We have mapped the location of five key bone marrow ECM proteins in situ. Each protein was shown to have specific areas of localization, suggesting different structural roles in the marrow architecture as well as different functional roles in the regulatory aspects of hemopoiesis.
Of particular interest was the distribution of fibronectin. Reilly et al. (1985a) previously reported that, in human bone marrow biopsies, fibronectin was found only in the walls of arteries and in megakaryocytes. Similar to Reilly et al. (1985a,b), we found fibronectin in megakaryocytes, although we also found significant amounts of fibronectin throughout the marrow cords. This distribution has previously been reported as dividing the marrow of chick embryos (Sorrell 1988) and rats (Hamilton and Campbell 1991) into many small compartments, which were suggested to play a significant role in the differentiation of blood cell clones.
Contradictory findings have been reported concerning whether collagen Type I or III is the predominant collagen type of marrow (Bentley 1982; Bainton et al. 1986; Gordon 1988). Both collagen Type I and III have been shown in and radiating out from the walls of large blood vessels in the marrow of both humans (Bentley et al. 1981) and rats (Hamilton and Campbell 1991). The present study suggests that collagen Type I is predominant in the mouse, with very high levels evident in the bone and endosteum. In contrast, collagen Type III was found only in the periosteum. In this study, neither collagen Type I nor Type III was found in or radiating out from the walls of large blood vessels. The reason for this difference is unknown but may be due to differences in species, sampling procedures, or antibodies used for detection.
Laminin has been shown to line rat sinuses in a discontinuous manner (Hamilton and Campbell 1991), and collagen Type IV and laminin have also been demonstrated in human sinuses (Reilly et al., 1985b), although very infrequently. We have confirmed these observations in the mouse, demonstrating laminin and collagen Type IV lining both arteries and sinuses. However, the frequency of positive vessels was significantly higher than that previously reported in humans. Supporting previous studies, expression of laminin was intense in the wall of arteries and was discontinuous along the walls of marrow sinusoids.
The use of perfused, fixed whole femoral sections has enabled us to extend these observations and characterize ECM protein distribution in the endosteal and periosteal regions, as well as within the bone matrix. The use of a longitudinal section of whole femur is distinctly different from and has many benefits over a bone marrow biopsy. The perfusion-fixation at physiological pressure preserves the structural integrity of the natural femoral microenvironment in situ without any sampling-induced tissue damage. Although a biopsy specimen does allow examination of the immediately surrounding cells, most structural elements, such as bone, are lost, and sample damage may occur during the biopsy process.
We have shown very high levels of fibronectin in situ in the endosteum, periosteum, and the femoral bone matrix itself. These observations support the study by Weiss and Reddi (1981), who showed high levels of fibronectin, especially at the periphery, in developing bone matrix on subcutaneously implanted bone plaques. It was interesting to observe the variability in levels of collagen Type I positivity, as well as the discontinuous labeling of collagen Type IV, within the endosteum. The significance of the regions of high positivity of these two ECM proteins in this area remains unclear. It was also interesting to observe the presence of all of these ECM proteins except for collagen Type I in the periosteum. The functional role of this is presently unknown.
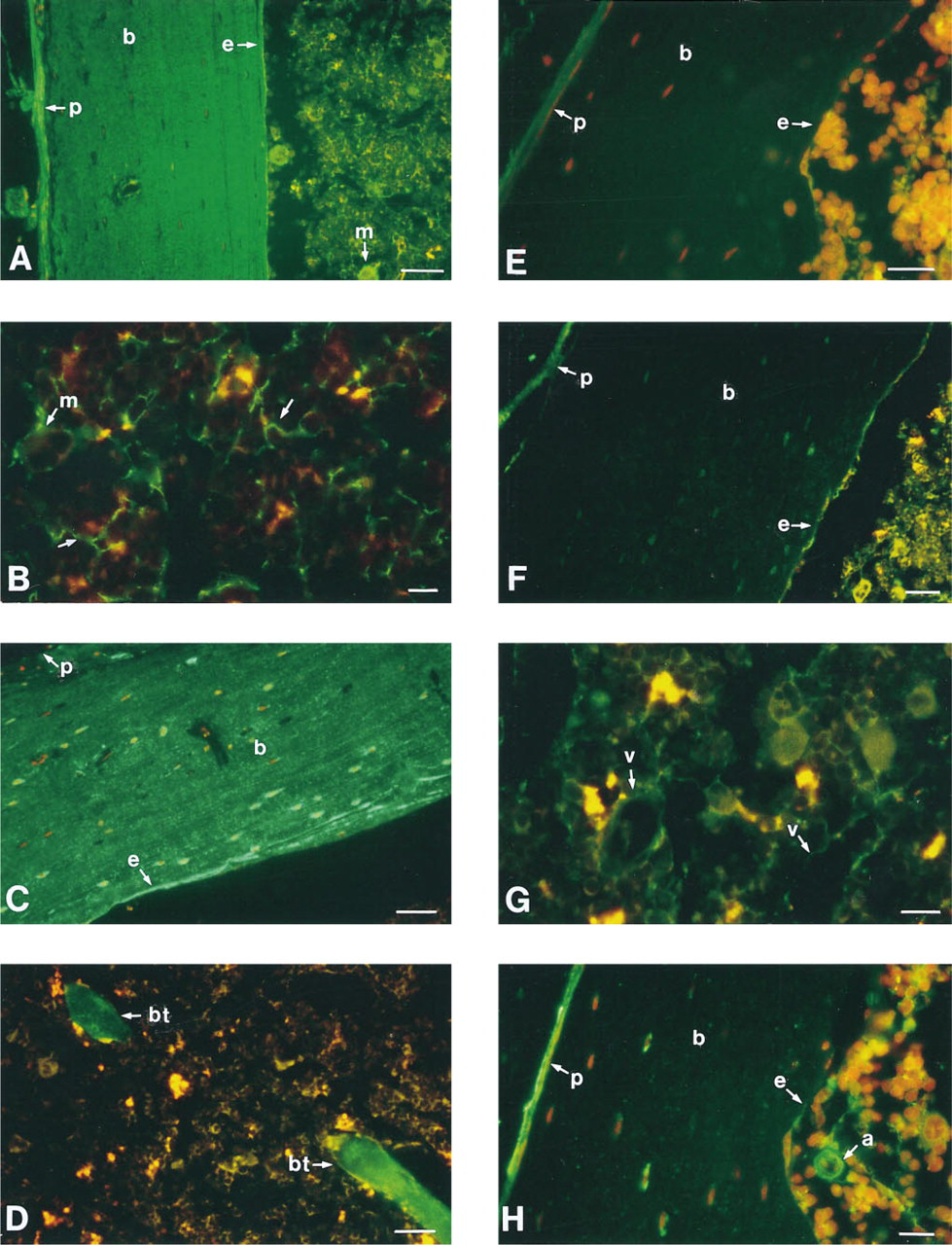
Longitudinal sections of whole murine femurs embedded in paraffin and labeled with antibodies specific for the extracellular matrix proteins fibronectin, collagen Types I, III, and IV, and laminin. Sections are of the bone and marrow interface (
There is strong evidence that the regulation of hemopoiesis is due to interactions between the bone marrow microenvironment and hemopoietic cells and is a consequence of mutual recognition and adhesion. For example, transplanted repopulating stem cells “home” to the bone marrow. In LTBMC, hemopoietic activity is localized within the adherent layer, indicating a physical association between stem cells and components of the microenvironment.
Evidence also suggests that marrow cells are located according to lineage specificity and maturational stage, delineated by the expression of adhesion receptors and by adhesive interactions with components of the marrow microenvironment. Integrins are the major receptors by which cells attach to the ECM, and some integrins also mediate important cell–cell adhesion events (Hynes 1992). For example, the β1 subunit α-chains bind differentially to fibronectin, collagen, and/or laminin. The different α-chains are also expressed on a variety of marrow cell populations, such as immature blasts, erythroid precursors, myeloid cells, osteoblasts, and megakaryocytes (Soligo et al. 1990; Williams et al. 1991; Kerst et al. 1993). Recently our laboratory showed that the murine lineage-negative, SCA+ and/or rhodaminedull, Hoechstdull hemopoietic stem cell populations expressed many of the β1 class α-subunits (manuscript in preparation).
There is also extensive evidence that different hemopoietic cell populations have adhesive interactions with ECM components (Long 1992). For example, adhesive properties have been used to distinguish hemopoietic cells with different spleen colony-forming and marrow-repopulating abilities (Bearpark and Gordon 1989). Immature myeloid and erythroid cells have been shown to bind to both fibronectin (Giancotti et al. 1986) and collagen Type I (Tsai et al. 1987; Koenigsmann et al. 1992). Human CD34+ progenitors bind to fibronectin in an activation-dependent manner (Kerst et al. 1993). In addition, human osteoblasts have a high affinity for collagen Type I and, to a lesser extent, fibronectin (Saito et al. 1994).
This view is consistent with the hemopoietic “niche” hypothesis, which proposes specific associations between stem cells and stromal elements (Schofield 1978). We recently reported that 6 weeks after a transplant of “stem cells,” donor cells had a high spatial affinity for the endosteal surface (Nilsson et al. 1997). These data add to the mounting evidence that “stem cell niches” have an intimate association with bone (Lord et al. 1975; Schofield 1978; Taichman et al. 1996). Our present data show that the ECM proteins fibronectin, collagen Type I and, to a lesser extent, collagen Type IV have a high spatial association with this area. As a consequence, they may contribute to the regulation of “stem cells” either as a structural component of the “niche” or in the maneuvering to and from the “niche.” As described above, primitive hemopoietic progenitor cells express the integrin α4, which binds to fibronectin. In addition, Williams et al. (1991) demonstrated that an anti-α4 antibody blocked Day 12 spleen colony-forming units (CFU-S12) from binding to fibronectin in vitro. In vivo, CFU-S formation could be inhibited after a transplant of β1 blocked bone marrow cells. However, because β1 binds to many other α-chains with alternative functions, the significance of this in vivo finding is unclear. In addition, Becker et al. (1996) recently showed that blocking α4 in a transplant of whole marrow cells did not affect engraftment levels 6 weeks post transplant into nonablated animals.
Summary of extracellular matrix proteins and their spatial localization within the femoral bone marrowa
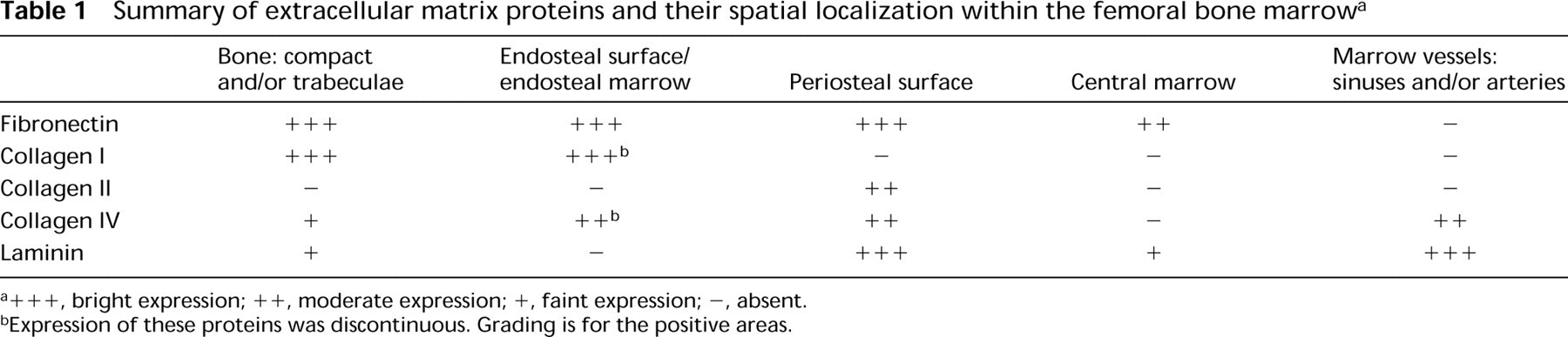
+ + +, bright expression; + +, moderate expression; +, faint expression; -, absent.
Expression of these proteins was discontinuous. Grading is for the positive areas.
The presence of different ECM proteins in all areas of the bone marrow may potentially play an important role in the process of cell homing and engraftment. Future in vitro binding studies and in vivo blocking studies of stem cell-enriched marrow sub-populations to ECM proteins will help to further delineate possible regulatory roles.
Footnotes
Acknowledgements
Supported by NIH grants DK #49650-02 and DK #50222-01 and by HL #02656-05. SN is a CJ Martin Fellow granted from the National Health and Medical Research Council, Australia, and is supported by the Our Danny Cancer Fund, University of Massachusetts Medical Center.