
Abstract
The rabbit suprapatella is a sesamoid fibrocartilage in the deep surface of the tendon of vastus intermedius and an integral part of the knee joint. We report the presence of a variety of proteoglycans (aggrecan and versican), glycosaminoglycans (chondroitin 4 and 6 sulfate, dermatan sulfate, keratan sulfate) and glycoproteins (tenascin) in its extracellular matrix and the intermediate filament vimentin in the fibrocartilage cells. The most significant finding is the presence of aggrecan in the extracellular matrix, along with its associated link protein and several of its integral glycosaminoglycans. Aggrecan probably enables the suprapatella to withstand compression. Although it can be assumed that aggrecan metabolites detected in synovial fluid from some human joints are predominantly associated with articular hyaline cartilage, the presence of aggrecan in the rabbit suprapatella means that this cannot be assumed for all animal knee joints. We conclude that it is important for orthopedic researchers who use animal models for arthritis research to check for the presence of a suprapatella when joint fluid analyses are interpreted.
A number of animals, including the rabbit, have been used as experimental models in studies of osteoarthritis or rheumatoid arthritis (Olszewski et al. 1996; Wei et al. 1996; Rayan et al. 1998). In an effort to monitor disease progression, breakdown products of cartilage are analyzed in the joint fluid. Among the metabolites of greatest interest are those of aggrecan, the large proteoglycan (PG) responsible for the pressure-tolerant properties of articular cartilage (Heinegärd and Oldberg 1993). It is generally assumed that such metabolites originate from hyaline cartilage and not from anywhere else (Lundberg et al. 1996; Ratcliffe et al. 1996; Kojima et al. 2001). However, a conspicuous feature of the knee joints of many animals used in arthritis research is a curious sesamoid fibrocartilage, called the suprapatella, embedded in the deep surface of the quadriceps tendon immediately above the patella itself. It contacts the joint cavity, articulates with the femur, and is an integral part of the knee joint. It is every bit as much an “articular” cartilage as that covering the bones themselves. A suprapatella is characteristic of many mammals, including the mouse, rat, rabbit, lemur, cat, and dog (Lessertisseur and Saban 1967; Jungers et al. 1980; Walji and Fasana 1983; Ralphs et al. 1991, 1992; Benjamin et al. 1995; Bland and Ashhurst 1997), but is absent in humans. It is one of the most striking examples of how tendons become fibrocartilaginous in regions where they are subject to compression, e.g., where they wrap around bony pulleys. Other fibrocartilaginous “wraparound tendons” include the bovine deep flexor tendon (Vogel and Koob 1989), the rabbit flexor digitorum profundus tendon (Merrilees and Flint 1980), and many tendons in humans (e.g., extensor digitorum communis, peroneus longus, Achilles tendon, peroneus brevis, and tibialis posterior) (Vogel et al. 1993; Benjamin et al. 1995; Milz et al. 1999).
Sources and labeling characteristics of the monoclonal antibodies used a Antigen(s) recognized
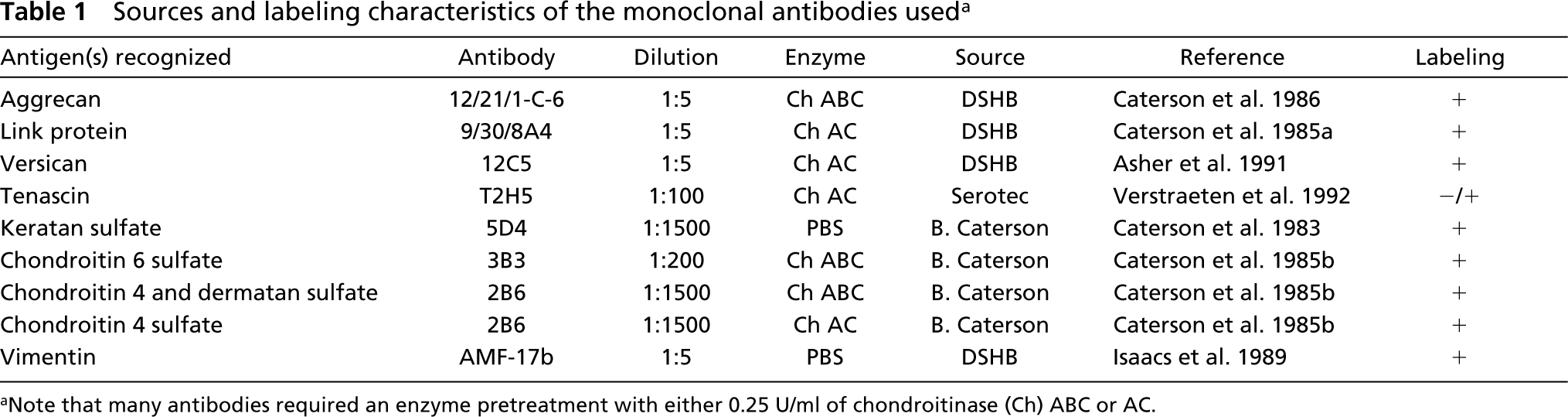
Note that many antibodies required an enzyme pretreatment with either 0.25 U/ml of chondroitinase (Ch) ABC or AC.
Wrap-around tendons are frequently characterized by their content of aggrecan and Type II collagen (Vogel et al. 1994; Waggett et al. 1998; Milz et al. 1999; Rees et al. 2000), molecules that are also found in articular cartilage. Although Type II collagen has been detected in the suprapatella (Ralphs et al. 1991; Bland and Ashhurst 1997), aggrecan has not been investigated. The primary purpose of the present study was therefore to demonstrate that aggrecan, together with its associated link protein and glycosaminoglycans (GAGs), is indeed found in the rabbit suprapatella. We also report the distribution of versican, tenascin, and vimentin. These molecules are present in wrap-around tendons and, like aggrecan, they are often assumed to have a mechanical role (Benjamin et al. 1994; Mehr et al. 2000; Boszczyk et al. 2001; Milz et al. 2001).
Materials and Methods
The suprapatellae were dissected out from the hind limbs of six mature female Chinchilla-bastard rabbits (mean body weight 4500 g) that were sacrificed by IV injection of ketamine, xylazine, and thiopental for control purposes associated with a separate and unrelated study by other researchers. The tissue was fixed in 90% methanol at 4C for at least 24 hr, infiltrated overnight in a 5% sucrose solution in PBS at pH 7.4, mounted on chucks in Jung tissue embedding medium (Leica; Oberkochen, Germany), frozen in an HM 500 OMV cryostat (Microm; Walldorf, Germany), and cryosectioned at 12 μm. Sections were stained with toluidine blue for metachromasia and labeled with a panel of monoclonal antibodies directed against PGS, GAGs, glycoproteins, and proteins (Table 1). All sections were treated with 0.3% hydrogen peroxide in methanol for 30 min to block endogenous peroxidase activity, and nonspecific binding of the secondary antibodies was reduced by blocking with normal horse serum for 60 min. Control sections were incubated with normal mouse immunoglobulins (10 μg/ml) or the primary antibody was omitted and sections were treated with PBS alone. Antibody binding was detected with a Vectastain ABC Elite avidin-biotin kit (Vector Labs; Burlingame, CA) and sections were counterstained with Mayer's hematoxylin.
Many of the primary antibodies required some enzymatic predigestion step or other chemical pretreatment (Table 1). This served either to expose epitopes hidden by the extracellular matrix (ECM; antibodies to tenascin and versican), or to generate stubs carrying neoepitopes (antibodies to the chondroitin sulfates, link protein, and aggrecan). It is important to note that enzyme pretreatment is critical for the subtle distinctions among the different GAGs (Caterson et al. 1983, 1985b) and that, before the detection of aggrecan and link protein, a neoepitope must be generated by reducing the sections with 10 mM dithiothreitol in 50 mM Tris-HCl, 200 mM sodium chloride, pH 7.4, for 2 hr at 37C and alkylating them with 40 mM iodoacetamide in PBS for 1 hr (Calabro et al. 1992). Both the epitopes recognized by the antibodies to aggrecan and versican and lie in the hyaluronan-binding region of the molecules (Asher et al. 1991; Calabro et al. 1992). The two epitopes recognized by the link protein antibody lie in the tandem-repeat domains that are also concerned with hyaluronan interactions (Calabro et al. 1992).
(
Results
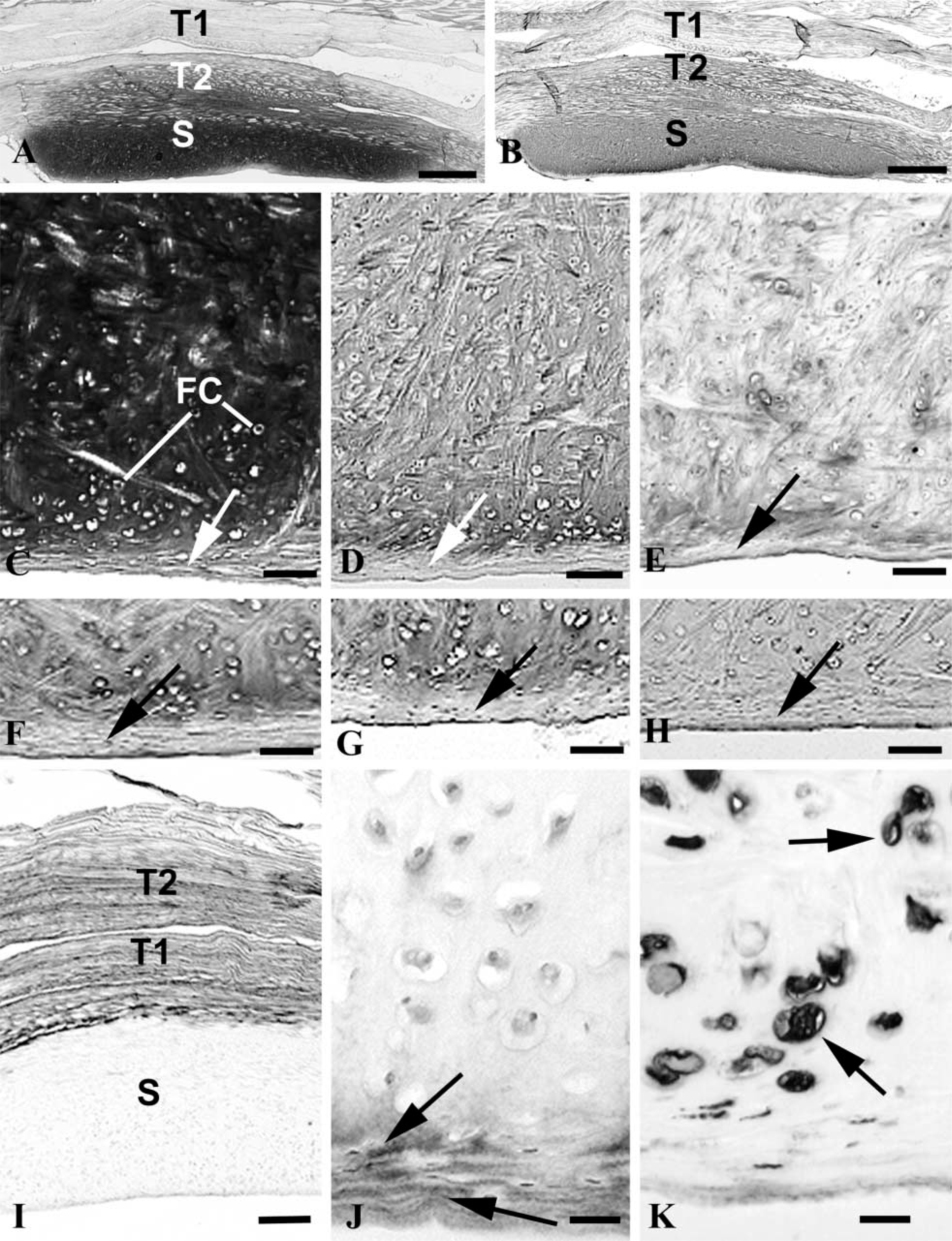
The suprapatella appeared as an intensely metachromatic sesamoid fibrocartilage in the deep part of the tendon of vastus intermedius (Figure 1A). However, near the articular surface of the suprapatella, there was a thin layer of poorly staining fibrous tissue (Figure 1C) in which the cells were flattened and arranged with their long axes parallel to the surface. Immediately deep to this fibrous zone were large chondrocytes separated by an ECM with a more hyaline appearance (Figure 1C). Deeper still, the majority of the metachromatic region of the suprapatella was characterized by a basketweave-like arrangement of collagen fibers. The immunohistological staining characteristics of the fibrocartilage are summarized in Table 1.
Aggrecan was highly characteristic of the suprapatella and strong labeling was detected in most parts of the tissue except for its superficial fibrous zone, where labeling was much weaker (Figures 1B and 1D). It was also present in the deep part of the tendon of vastus intermedius, although not in its superficial part (Figure 1B). Adjacent sections labeled for link protein, although the labeling was patchy (Figure 1E). The distribution of versican was broadly complementary to that of aggrecan. Therefore, whereas strong labeling characterized both parts of the tendon of vastus intermedius (Figure 1I), labeling in the suprapatella was largely restricted to the superficial fibrous zone, although some labeling was also seen in the larger fibrocartilage cells (Figure 1J). Several of the GAGs that form part of aggrecan (i.e., keratan sulfate, chondroitin 4 and 6 sulfates, and dermatan sulfate) were also detected in the suprapatella (Figures 1F–1H; Table 1).
Tenascin labeling was rather variable, but most of the suprapatella was generally devoid of label. Where staining was detected, it was usually in the superficial fibrous zone and in both parts of the tendon of vastus intermedius. Fibrocartilage cells in the suprapatella labeled strongly for vimentin (Figure 1K), but there were only a few strands of label in tendon cells.
Discussion
Aggrecan is a large aggregating PG commonly associated with the water-trapping properties of articular cartilage. The profusion of its negatively charged GAG side-chains creates domains with a high osmotic pressure and great potential to suck water into the tissue (Heinegård and Oldberg 1993). It is the incompressibility of water that gives cartilage its ability to resist compression. We propose that aggrecan plays a similar role in the rabbit suprapatella in view of the load-bearing function of this organ, a role accentuated by the highly flexed position of the knee during normal posture. It is pertinent to note that jumping animals (in which there is repetitive compressive stress on the quadriceps tendon during the hyperflexed phase of leaping) generally have a very conspicuous suprapatella (Lessertisseur and Saban 1967; Jungers et al. 1980; Walji and Fasana 1983). Furthermore, the compression tolerance function of aggrecan is further supported by findings of the molecule at other sites where tendons are pressed against bone. Thus, aggrecan is present in sesamoid fibrocartilages found in the human tibialis posterior tendon (Vogel et al. 1993), bovine deep flexor tendons (Vogel et al. 1994; Vogel 1995), and the extensor tendons of the fingers (Milz et al. 1999). All are pressed against bone in certain joint positions.
Our results are of general interest in relation to animal model studies of osteoarthritis and rheumatoid arthritis, because a suprapatella is present in the knee joints of several laboratory animals used for this purpose. These include not only the rabbit but also the guinea pig, mouse, rat, cat, and dog (Lessertisseur and Saban 1967). In both osteoarthritis and rheumatoid arthritis there is a gradual destruction of articular cartilage so that the underlying bone is eventually exposed. Attempts to monitor early stages in disease progression have focused on the development of molecular markers for cartilage breakdown that are released into the joint fluid. Among these are metabolites of aggrecan, cartilage oligomeric matrix protein (otherwise known as COMP), and collagen II C-propeptide (Lohmander 1997; Lohmander and Felson 1997; Mansson et al. 2001). The general assumption is that any such molecule found in aspirates of synovial fluid must have come from articular hyaline cartilage. Similar reasoning is used in studies on rheumatoid arthritis (Wollheim 1996; Mansson et al. 2001). Although it may be reasonable to assume that aggrecan metabolites detected in synovial fluid from human shoulder joints are predominantly associated with articular hyaline cartilage (Ratcliffe et al. 1996), the presence of aggrecan in the suprapatella means that this is not necessarily the case for animal knee joints. Indeed, Koob et al. (1995) have made a similar suggestion previously for knee joint menisci. It is therefore intriguing to note that the suprapatella itself is subject to degenerative changes in osteoarthritis. Indeed, the earliest signs of change occur in the former rather than the latter (Benjamin et al. 1995).
The distribution of versican in the suprapatella is the reciprocal of that of aggrecan. Therefore, it is more conspicuous in the overlying quadriceps tendon than in the suprapatella itself, and within the latter it is most prominent in the superficial fibrous zone, i.e., where aggrecan labeling is much weaker. Such a restriction of labeling largely to the periphery of cartilaginous tissue has been reported previously for versican in developmental studies (e.g., Shibata et al. 2001). Although this may suggest an association with higher levels of shear, it is also in line with other work on tendons and ligaments suggesting that versican is more characteriztic of their fibrous than their fibrocartilaginous regions (Waggett et al. 1998; Milz et al. 2001). This would indicate a closer relationship with tensile loading.
It is intriguing that some intracellular signal for versican was evident in the fibrocartilage cells of the suprapatella. This suggests either that the cells can synthesize versican without releasing it to the ECM or that it is destined only for the immediate pericellular matrix. It is therefore pertinent to note that the molecule can bind to cell membranes (Kjellen and Lindahl 1991) and that it may play a role in regulating cell motility, growth, and differentiation (Perides et al. 1992; Hayes et al. 2001). Subtle hints of versican label can also be seen in other fibrocartilage cells, because the illustrations in Melrose et al. (2001) suggest that it is occasionally seen in the annulus fibrosus cells of elderly sheep. However, Melrose et al. (2001) consider versican to be most typical of the ECM between adjacent annular lamellae and suggest that it contributes to the viscoelastic properties of the tissue.
Tenascin labeling in the suprapatella was highly variable, even though large rounded fibrocartilage cells were always present. Although labeling was occasionally seen around these cells, it was sometimes more obvious in the more flattened cells near the surface of the patella. Therefore, our results do not fit easily with the suggestion of Mehr et al. (2000) that tenascin-C acts as an anti-adhesive protein in tendon fibrocartilage, allowing the cells to maintain a rounded shape and thus the tissue to have a fibrocartilaginous phenotype. There were methodological differences between the two sets of procedures, but these alone do not appear sufficient to account for the different results. We used a monoclonal antibody that recognizes a range of tenascin isoforms (including tenascin C) and pretreated our sections with chondroitinase AC. Mehr et al. (2000) focused exclusively on tenascin C and used no epitope unmasking procedure. Neither are the results likely to reflect species differences, for tenascin C is certainly characteristic of rabbit cartilage (Mackie and Ramsey 1996).
Vimentin expression has also been associated with a fibrocartilage phenotype (Benjamin et al. 1994), and the presence of vimentin in suprapatellar cells is in line with previous reports in the rat (Ralphs et al. 1991). It is possible that this cytoskeletal component could enable the cells themselves to resist compression. This is supported by the experimental studies of Durrant et al. (1999), who showed that vimentin expression is modified according to the hydrostatic load placed on the tissue.
We therefore conclude that it is important for orthopedic researchers to be aware of the suprapatella and to consider its presence when joint fluid analyses in animals widely used for arthritis research are interpreted. This structure is commonly neglected or overlooked, but it can be as prominent as the patella itself. It contains a wide range of molecules that are also expressed in articular cartilage and is present in a number of animals that are highly significant in orthopedic research.
Footnotes
Acknowledgements
Supported by the Friedrich Baur Stiftung Munich. The monoclonal antibodies 12/21/1-C-6, 9/30/8A4, 12C5 and Amf-17b were obtained from the Developmental Studies Hybridoma Bank maintained by The University of Iowa, Department of Biological Sciences, Iowa City, IA 52242, under contract NO1-HD-7-3263 from the NICDH.
The practical work was performed at the Anatomy Department, Munich.