
Abstract
We examined the expression of laminin-5 and its integrin receptors during reepithelialization of human wounds. We used suction blisters of skin as a model of keratinocyte migration on a basement membrane matrix and mucosal full-thickness wounds as a model in which keratinocytes migrate in a provisional matrix. An animal model, in which human epidermal keratinocytes were injected into the back of athymic mice, was used to follow the deposition of the basement membrane components. In 4-day-old blisters, about 20–50 cells at the leading edge of the migrating tongue showed cytoplasmic laminin-5 immunostaining. Laminin-5 mRNA was detected in 15–30 cells at the leading edge of the migrating epidermis. α3β1 and α6β4 integrins were found in membrane projections of the migrating basal cells and also in suprabasal cell layers, suggesting their combined role in binding laminin-5. In mucosal wounds, laminin-5 was the only basement membrane zone component that was deposited between the clot and the migrating keratinocytes. In the animal model, linear deposition of laminin-5 and α6β4 integrin was already seen on Day 2, whereas the other basement membrane zone components were not yet organized. The results suggest that, regardless of the injury and the microenvironment, laminin-5 plays an essential role in the interaction between wound keratinocytes and the surrounding matrix.
L
LM-5 appears to be one of the best ligands for keratinocyte adhesion and migration. It can also promote the formation of hemidesmosomes in vitro (Hormia et al. 1995). Previous investigations have shown that LM-5 is expressed by migrating keratinocytes during reepithelialization of experimental human full-thickness wounds (Larjava et al. 1993). Both α3β1 and α3β1 integrins can act as cell receptors for LM-5 in keratinocytes (Carter et al. 1991; Niessen et al. 1994). Migrating keratinocytes express both of these integrins. It has been proposed that the deposition of LM-5 may be necessary for keratinocyte migration and relocation of integrins from their preexisting functions during reepithelialization and cell differentiation (Larjava et al. 1993). LM-5 is also expressed by squamous cell carcinomas and metastatic lesions arising from these lesions (Pyke et al. 1995; Kainulainen et al. 1997). Therefore, it is suspected that tumor cells of epithelial origin may utilize LM-5 for cell invasion and migration. Because increased expression and abnormal distribution of LM-5 are already present in some of the premalignant oral lesions, LM-5 has also been proposed to play a role in the transformation of dysplastic lesions into invasive carcinomas (Kainulainen et al. 1997).
The aim of the present investigation was to study the role of LM-5 in physiological keratinocyte migration regardless of the microenvironment and injury type of the wound. We used three different experimental models: a suction blister of skin, in which keratinocytes migrate on a BM substratum; a full-thickness mucosal wound, in which keratinocytes migrate through the clot matrix; and an animal model, in which early BM organization can be followed between keratinocytes and the surrounding healthy connective tissue matrix. The results suggest that regardless of the surrounding matrix, LM-5 deposition appears to be critical for cell migration and BM zone organization during reepithe-lialization. Among the cytokines tested in vitro, TGFβ and INFγ appear to act as potential upregulators of LM-5 expression.
Materials and Methods
Tissue Samples
Suction blisters were induced on the abdominal skin of 12 volunteers, aged 22–66 years (Kiistala 1968). In most cases the roof of the blister was removed right after blister induction. Biopsies were taken immediately after blister formation (one case), on Day 2 (two cases), on Day 4 (10 cases), and on Day 9 (one case). One half of each sample was snap-frozen in liquid nitrogen. The other half was fixed in 10% phosphate-buffered formalin and embedded in paraffin.
Experimental human mucosal wounds (full-thickness) were made in human palate as described previously (Larjava et al. 1993). Five subjects volunteered for this study. Punch biopsies were taken from the wounds on Day 3 and frozen in liquid nitrogen.
A spontaneously transformed nontumorigenic human adult skin epithelial cell line that has maintained epithelial differentiation properties (HaCat) (Boukamp et al. 1988) was cultured to confluency in Dulbecco's modification of Eagle's medium containing 10% fetal calf serum and was detached by trypsin. The cells were rinsed and mixed with culture medium without serum and injected (0.2 ml containing 5 X 106 cells) sc into the back skin of athymic mice (Fox Chase Scid, 4 weeks old; Bomholtgaard Breeding and Research Centre, Bommice, Denmark) as described previously (Lavker and Sun 1983). After 2 or 7 days, the animals were sacrificed and the cysts were removed for immunohistochemical analysis.
Immunohistochemical Studies
Immunostaining by an avidin–biotin technique (Vector Elite kit; Abbott, Chicago, IL) was performed as described previously (Kainulainen et al. 1997). Paraffin sections of 14 suction blisters were stained with polyclonal antibody against the LM-5 γ2-chain (Pyke et al. 1995).
Immunofluorescence staining of integrins and BM zone components was performed as described earlier by Larjava et al. (1993). Antibodies against the following proteins were used for the frozen sections: α3β1 integrin (MAb J143) (Kantor et al. 1987); α6β4 integrin (recognizes β4 chain) (Kennel et al. 1989); Type IV collagen (MAb 68-124-1; ICN Biomedicals, Costa Mesa, CA); Type VII collagen (MAb 1345; Chemicon, Temecula, CA); and LM-5 (GB3) (Verrando et al. 1987) known to react with the γ2-chain of LM-5 (Matsui et al. 1995).
Control stainings were performed without the primary antibodies and with nonimmune mouse/rabbit serum.
In Situ Hybridization
In situ hybridization was done on 10 4-day-old blister samples as described previously by Autio-Harmainen et al. (1991). For the laminin γ2-chain, a PstI-EcoRI fragment of clone L15 (bases 2995–3840) was subcloned into the pSP64 and pSP65 vectors in the sense and anti-sense orientations (Kallunki et al. 1992). The probe in the sense orientation served as a control for nonspecific hybridization. The estimated number of grains was calculated with an image analysis program MCID-M2 (Image Research; St. Catherines, Canada). The microscopic image was acquired with a three-chip color camera (Sony 930).
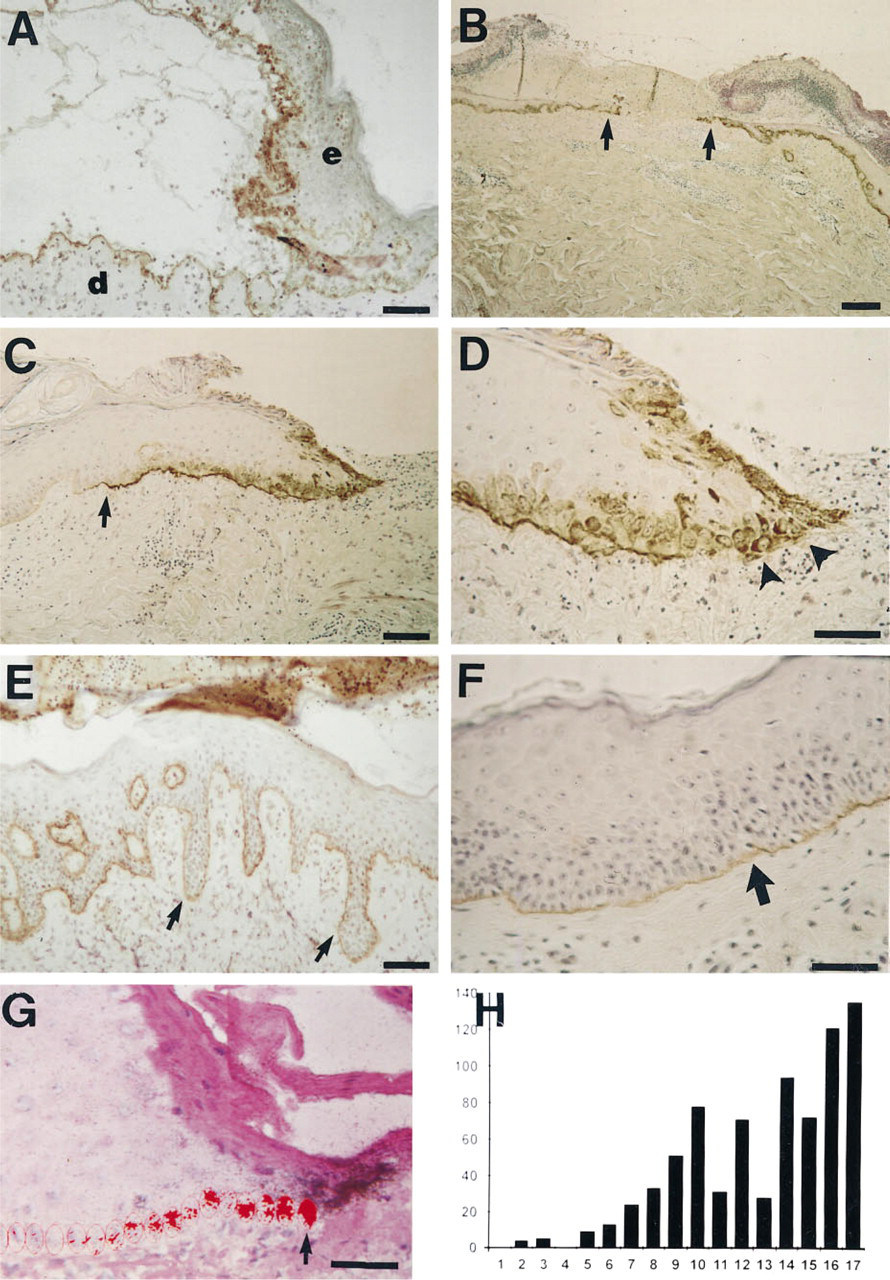
In 2-day-old blisters, LM-5 γ2-chain staining was found in the basal aspect of the basal keratinocytes of the detached epidermis and in the blister floor along the BM (
Immunoprecipitation Analyses
Normal human mucosal keratinocytes were isolated from surgical gingival biopsies. Keratinocytes (passage <6) were cultured as described (Salo et al. 1991) and exposed to TGFβ (5 ng/ml; a gift from Dr. Joan Massague, New York), TNFα (20 ng/ml; Boehringer Mannheim, Mannheim, Germany), INFγ (1000 U/ml; Boehringer), and IL-1β (5 U/ml; Boehringer) in basal keratinocyte growth medium for 24 hr, followed by a medium change with fresh cytokines and radiolabeling with 100 μCi/ml [35S]-methionine (trans 35S label, specific activity >1000 Ci/mmol; ICN Radiochemicals, Irvine, CA) for 24 hr. Integrins were immunoprecipitated as described in detail elsewhere (Roberts et al. 1988), using polyclonal antibodies against the peptide sequences of the intracellular domain of the β1-integrin subunit (Larjava et al. 1990). Medium samples from the same cultures were used for immunoprecipitation of fibronectin and LM-5 (MAb GB3). Under these conditions, various subunits of fibronectin and LM-5 remain associated and result in a single band on the top part of the resolving gel about 1 cm from the starting point.
Results
Upregulation of the LM-5 γ2-Chain in Blister Wounds
In fresh suction blisters, the LM-5 γ2-chain was mostly visible in the blister floor, but some staining could also be seen in the basal keratinocytes of the detached epidermis (not shown). In 2-day-old suction blisters, the LM-5 γ2-chain was visible in the blister floor in areas not covered by regenerated epidermis, but also cytoplasmically in the keratinocytes at the blister roof, perhaps reflecting active synthesis of the LM-5 γ2-chain (Figure 1A). In 4-day-old blisters, staining of the LM-5 γ-chain was more intense in the areas covered by new epidermis than in normal skin (Figures 1B-D) (Table 1.). At the leading edge of the regenerating epidermis, covering a distance of about 20–50 cells from the tip, the LM-5 γ2-chain was expressed in the keratinocyte cytoplasm (Figures 1C and 1D). Behind these cells, the LM-5 γ2-chain was present as a thin continuous line at the basement membrane zone (Figures 1C and 1D). Table 1 shows a summary of the immunostaining results from 10 subjects. In 9-day-old blisters, the entire blister base was covered by a new acanthotic epidermis, and LM-5 γ2-chain staining was comparable to that of the controls (Figures 1E and 1F).
The expression of the LM-5 γ2-chain in human blister wounds was then studied in 10 cases of 4-day-old blisters by in situ hybridization. In normal skin, no or only a very weak signal could be observed in basal keratinocytes (not shown). In regenerating epidermis, the signal for LM-5 γ2-chain mRNA was especially strong close to the leading edge, with a clear decreasing gradient towards the normal skin areas (Figures 1G and 1H). A strong signal for LM-5 γ22-chain mRNA could also be seen close to the tip of the leading edge in the keratinocytes that were not in apparent contact with the underlying dermis. The activation of the LAMC2 gene during wound healing was independent of the age of the subjects (Table 1). The pattern of LM-5 γ2-chain mRNA expression was remarkably similar in the healing blisters of the subjects aged 22–66 years.
LM-5 Receptors in Blister Wounds
In normal epidermis, α3β1 integrin was localized around basal keratinocytes, especially in the cell–cell contact areas (Figure 2A). Virtually no expression was seen in suprabasal cell layers. In 4-day-old blister wounds, α3β1 integrin was expressed throughout the cell layers of the leading edge. Remarkably strong staining was seen in the cell extension of the leading keratinocytes (Figure 2B). α6β4 Integrin was localized at the basal aspect of the basal keratinocytes of normal epidermis (Figure 2C). At the leading edge of healing 4-day-old blisters, α6β4 was localized around basal and supra-basal keratinocytes (Figure 2D).
Laminin-5 expression during reepithelialization of blisters in young and old healthy malesa

Suction blisters were induced by constant vacuum on the abdominal skin of healthy males. Blister roofs were cut off and 4 days after blister induction the entire blister area was biopsied.
+ + +, strong staining; ++, moderate staining; +, faint staining.
+ + +, strong labeling; + +, moderate labeling; +, faint/weak labeling; -, unconvincing/nonexistent labeling.
LM-5 in Full-thickness Mucosal Wounds
Using mucosal full-thickness wounds, we examined the distribution of the LM-5 γ2-chain under conditions in which keratinocytes are known to migrate through the clot (Larjava et al. 1993). The expression of LM-5 protein underneath migrating mucosal keratinocytes was semiquantified using a computer-assisted image analyzer. As in blister wounds, the LM-5 γ2-chain was continuously present underneath the migrating keratinocytes of 3-day-old wounds, whereas LM-1 and collagen Types IV and VII were almost completely absent (Figure 3). The expression of immunoreactive LM-5 protein was higher in the wound area compared to the non-wounded site in oral mucosa (Figure 3).
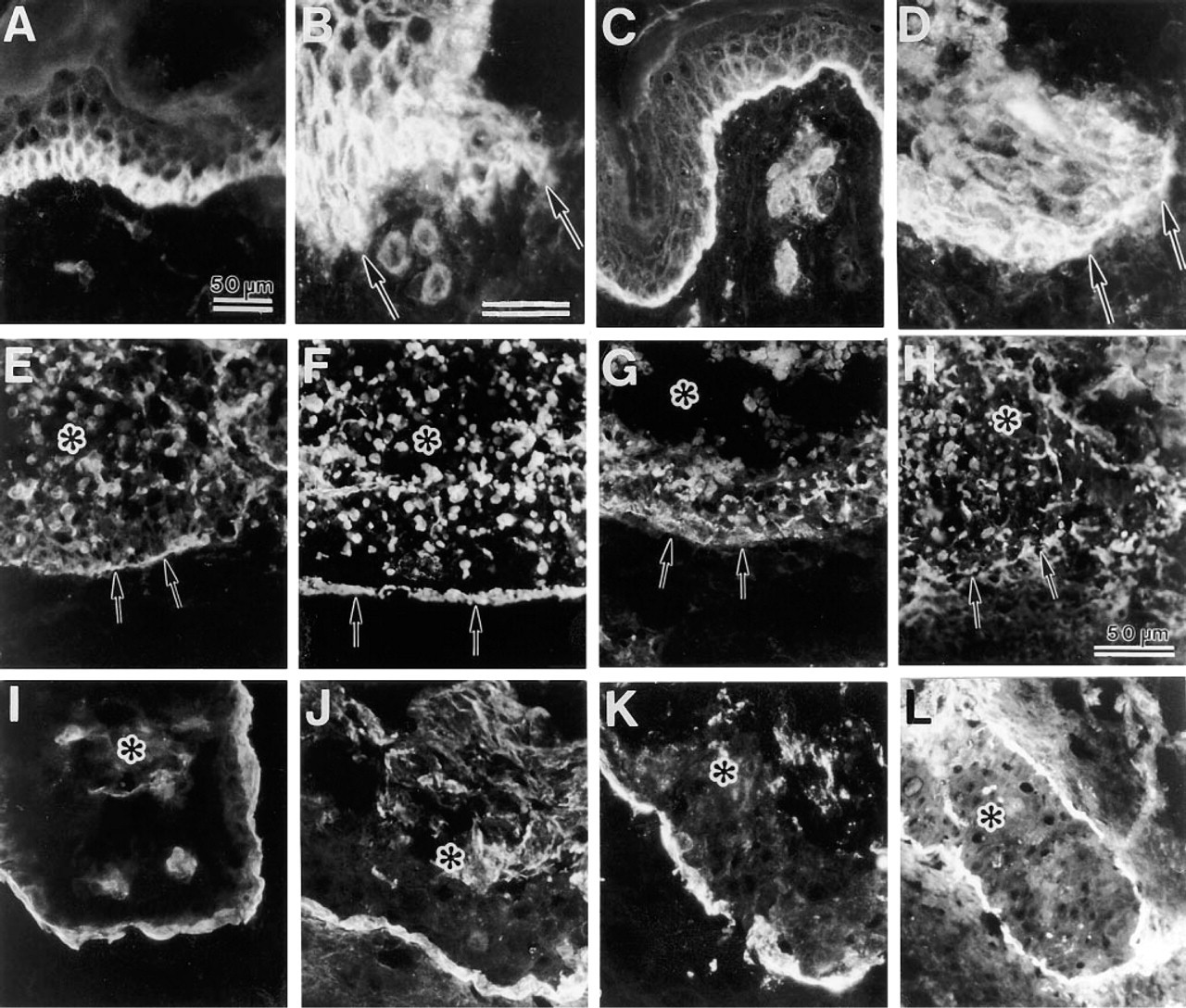
Expression of α3β1 (
Development of a BM Zone in a Cyst Cavity in Athymic Mice
Using the inclusion cyst model, we explored which component of the BM zone plays an initiating role when BM is deposited in inflammation-free connective tissue. In immature 2-day-old cysts of HaCat keratinocytes, the LM-5 γ2-chain was found both in peripheral cells and inside the cyst cavity (Figure 2F). Type VII and IV collagens were found only in cells inside the cyst (Figures 2G and 2H, respectively). The immunolocalization pattern of α6β4 integrin was similar to that of the LM-5 γ2-chain in 2-day-old cysts (Figure 2E). After 7 days the cysts appeared to be mature, with cells sloughing centrally. At this stage, all the components of the BM zone were localized around the cyst periphery (Figures 2I-2L). At that time, there were practically no staining signals in keratinocytes located inside the epithelial cysts.
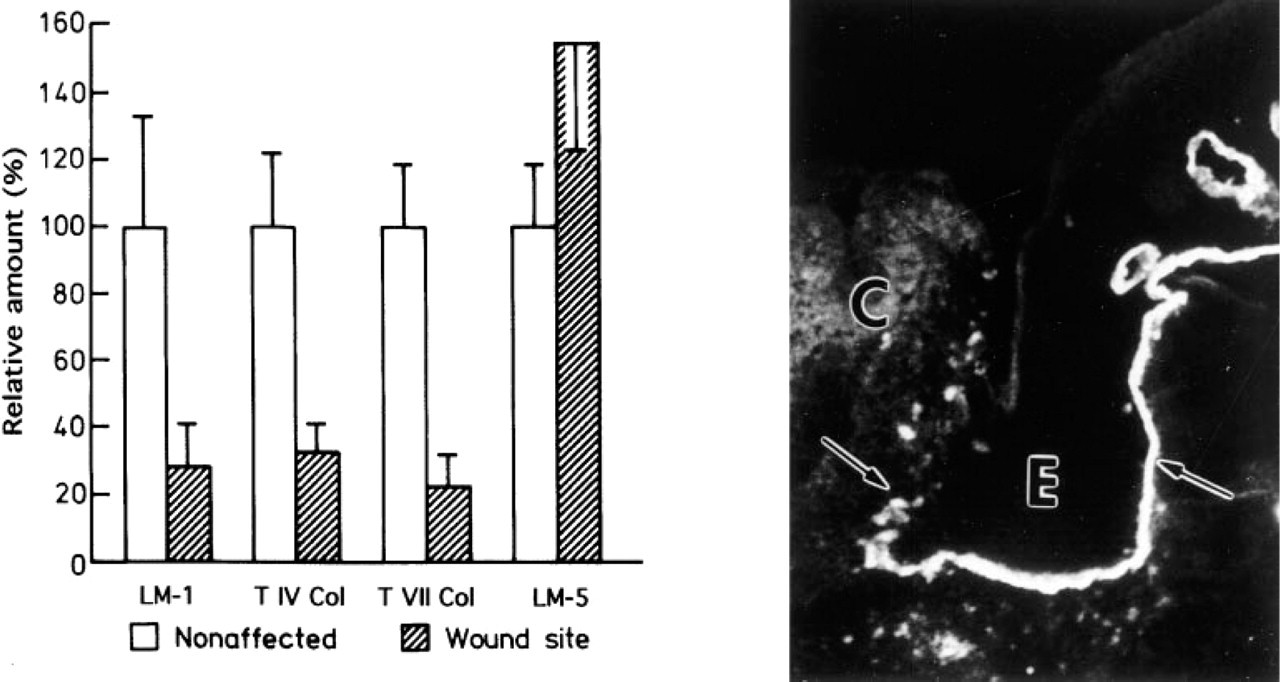
Relative amounts of BM zone components (LM-1, laminin-1; T IV Col, type IV collagen; T VII Col, type VII collagen; LM-5, laminin-5 γ2-chain) underneath keratinocytes in nonaffected and wound sites was quantified by measuring the brightness of immunofluorescence using a computer-assisted image analyzer. The LM-5 γ2-chain is present underneath migrating mucosal keratinocytes in 3-day-old wounds. E, epithelium; C, clot.
Regulation of LM-5 Expression in Cultured Mucosal Keratinocytes
Because LM-5 appears to be upregulated during epidermal and mucosal wound healing, we wanted to examine whether the cytokines present during the different various stages of wound healing could influence LM-5 expression. Among the cytokines tested, TGFβ and INFγ were found to upregulate the synthesis of LM-5 in cultured mucosal keratinocytes (Figure 4). Both of these cytokines also stimulated the biosynthesis of fibronectin, whereas they had no noticeable effect on the expression of β1 integrins (Figure 4). TNFα slightly stimulated the synthesis of fibronectin, whereas it did not have any visible effect on LM-5 synthesis (Figure 4).
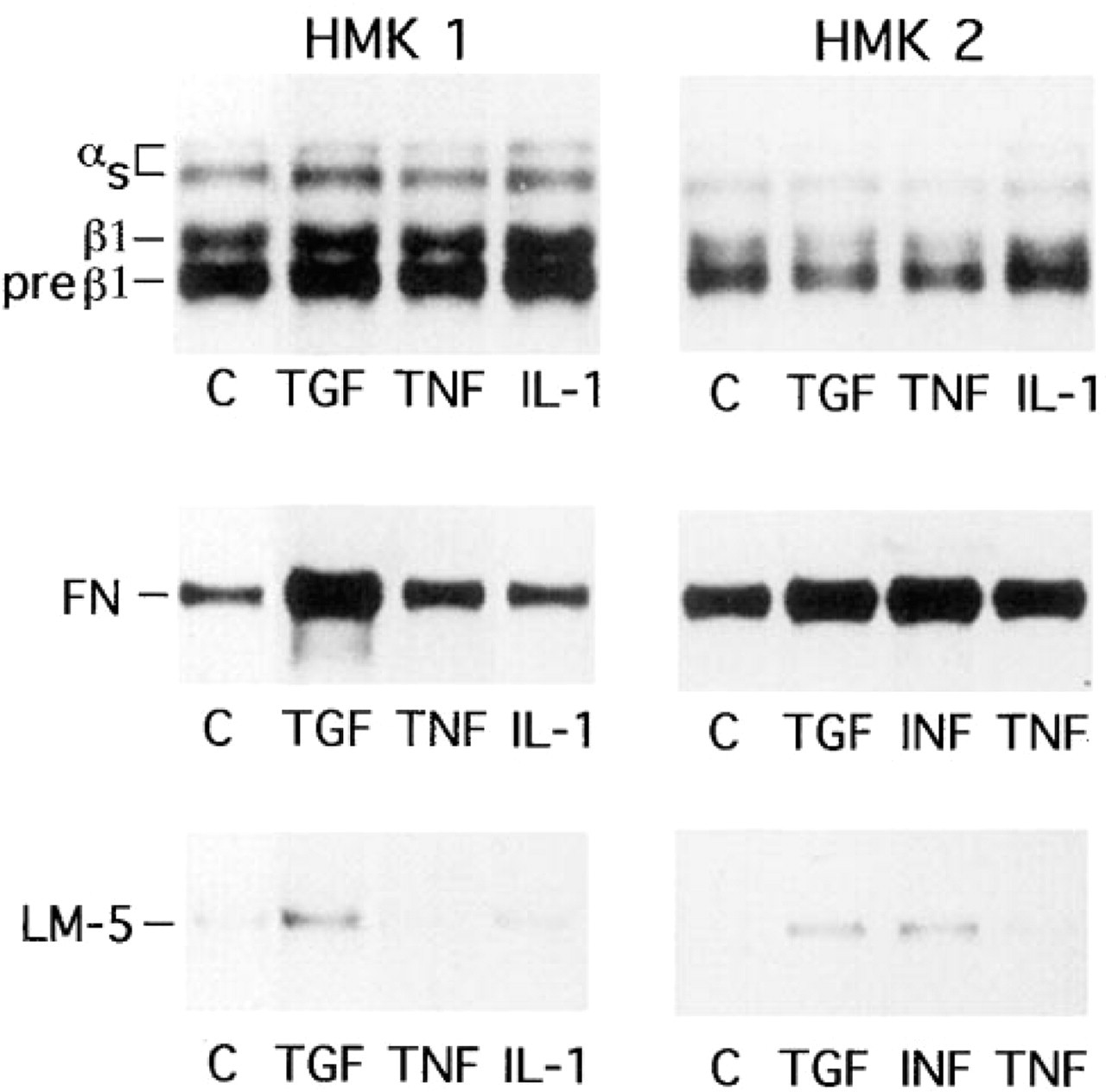
Human mucosal keratinocytes were exposed for TGFβ, TNFα, IL-1β, and INFγ, followed by simultaneous radiolabeling and immunoprecipitation of β1 integrins (upper panels), fibronectin (FN) (middle panels), and LM-5 (lower panels). Various integrin, fibronectin (FN), and laminin-5 (LM-5) bands are indicated. HMK 1 and 2 are two separate human mucosal keratinocyte cell lines.
Because the association of LM-5 protein with the cell culture fraction of keratinocytes, it is not possible to quantitate all the LM-5 deposited by keratinocytes, and these results therefore provide only semiquantitative information about the regulation of LM-5 by different growth factors.
Discussion
Regeneration of surface epithelium during wound healing takes place in a controlled fashion through several mechanisms, such as migration, proliferation, and differentiation (Clark 1995). Depending on the type of injury, keratinocytes must be able to interact with the BM zone matrix, the blood clot matrix composed mainly of fibrin and fibronectin, or with the connective tissue matrix, which contains various collagen types and proteoglycans. Keratinocytes might use two mechanisms to mediate this interaction. Either they vary their integrin receptors, adapting to the new matrix as proposed (Larjava et al. 1996), or they deposit an intermediating molecule linked to one receptor on the surface of keratinocytes able to bind various matrix components.
Suction blisters have proved to be a convenient model for studies of wound healing under conditions in which the BM remains relatively intact, containing Type IV collagen, for example (Oikarinen et al. 1982; Saarialho–Kere et al. 1995). In full-thickness mucosal wounds, the BM structure is mechanically broken and keratinocytes migrate through the fibronectin-fibrin-rich matrix clot (Larjava et al. 1993). The athymic mouse model (Grimwood et al. 1988; Larjava et al. 1993) provided us with an experimental tool to follow the reorganization of the BM zone in wound-like conditions within a healthy connective tissue matrix. HaCat keratinocytes appeared to form inclusion cysts very similarly to freshly isolated epidermal cells. Within 7 days a mature epithelial cyst-like structure was formed (Grimwood et al. 1988; Larjava et al. 1993).
During reepithelialization, abundant deposition of LM-5 molecules was observed by immunohistochemistry under the regenerating epidermis of suction blisters. The in situ hybridization findings confirmed the new transcription of LM-5 molecules, showing that the most actively synthesizing cells were at the leading edge of the regenerating epidermis. Because the total length of the reepithelialization was about 100–120 cells and the number of LM-5 γ2-chain mRNA-containing cells was 15–30 on Day 4, the activation of the LAMC2 gene is relatively transient. The expression pattern of the LM-5 γ2-chain mRNA was similar in blisters in which remnants of LM-5 could be seen in the blister floor and in those in which LM-5 was absent. This indicates that activation of the LAMC2 gene is required even when LM-5 molecules already exist in the wound bed. It therefore appears that LM-5 molecules, in the form of broken anchoring filaments, are not used by the migratory keratinocytes, but instead they deposit new LM-5 molecules for interaction with the surrounding matrix. In full-thickness incisional mucosal wounds, the deposition of LM-5 occurs clearly before the deposition of the other BM zone components, such as Types IV and VII collagen or laminin-1, as shown here and in our earlier publication (Larjava et al. 1993). These studies do not provide a conclusive answer to the question of whether LM-5 has a more important role in cell migration or in cell adhesion. There are studies in which LM-5 has been shown to be crucial for cell adhesion and to reverse the hypermotility of keratinocytes (Gagnoux-Palacios et al. 1996). On the other hand, it has been shown, employing JEB cells and normal keratinocytes, that exogenous LM-5 stimulates cell migration (Verrando et al. 1994).
The α3β1 and α6β4 integrins, receptors for LM-5, localize in similar areas as LM-5, suggesting that keratinocytes might use either of these integrins as their LM-5 receptor (Verrando et al. 1994). The distribution of these integrins in blister wounds and full-thickness mucosal wounds is also similar (Larjava et al. 1993). It has been proposed that α6β4 integrin could have two stages of interaction with LM-5 (Larjava et al. 1996), one during active migration, when the distribution of this integrin is diffuse (Kurpakus et al. 1991), and another when stable hemidesmosomes containing both LM-5 and α6β4 integrin develop.
In the epidermal inclusion cyst model, LM-5 and its receptor, α6β4 integrin, were seen to organize as early as Day 2, first at the cyst periphery, whereas there was no accumulation of Types IV or VII collagen. Previously, neither laminin-1 nor bullous pemphigoid antigen have been reported to be detectable in immature 2-day-old cysts but were reported to be present in mature 7-day-old cysts (Grimwood et al. 1988). Fibronectin appears to precede the formation of the BM zone in this model (Grimwood et al. 1988). Clearly, LM-5 could play a key role in BM organization against the fibronectin-rich matrix, because it appears to be the first keratinocyte-derived molecule to accumulate in the BM zone.
We found that TGFβ and INFγ were able to induce the expression of the LM-5 protein in cultured mucosal keratinocytes, whereas there was no effect with IL-1β or TNFα. This result is in accordance with the report by Korang et al. (1995), who found LM-5 mRNA levels to be stimulated in cultured epidermal keratinocytes, HaCat, and Balb/K cells by TGFβ but not by TNFα. TGFβ also induced the production of fibronectin, as reported previously (Roberts et al. 1988), whereas none of the cytokines tested had any clear effect on the production of β1 integrin, also supporting a previous report (Hertle et al. 1995). Among the wound cytokines, TGFβ and IFNγ appear to be the best regulators of LM-5 expression.
In conclusion, induction of the LM-5 gene and protein expression appears to be critical during reepithelialization, regardless of the injury or the microenvi-ronment of the wounds. The LM-5 gene can be activated by TGFβ and INFγ present in the wound fluids. LM-5 appears to be used as an intermediate bridging molecule between the α3β1 or α6β4 integrins of migrating keratinocytes and the wound matrix. At the later stages of healing, LM-5 could serve as the first anchoring element inhibiting cell migration and promoting the formation of stable contacts with BM components via hemidesmosomes.
Footnotes
Acknowledgements
Supported by grants from the Medical Research Council of the Academy of Finland, the Medical Research Council of Canada, the Cancer Societies of Finland, Finland's Cancer Institute, and the Sigrid Juselius Foundation.
We thank Ms Annikki Huhtela for expert technical assistance, Dr Juha Tuukkanen for assisting in the image analysis of in situ hybridizations, and Dr Vesa Koivukangas for providing biopsy material.