
Abstract
Agrin is a heparan sulfate proteoglycan (HSPG) that is highly concentrated in the synaptic basal lamina at the neuromuscular junction (NMJ). Agrin-like immunoreactiv-ity is also detected outside the NMJ. Here we show that agrin is a major HSPG component of the human glomerular basement membrane (GBM). This is in addition to perlecan, a previously characterized HSPG of basement membranes. Antibodies against agrin and against an unidentified GBM HSPG produced a strong staining of the GBM and the NMJ, different from that observed with anti-perlecan antibodies. In addition, anti-agrin antisera recognized purified GBM HSPG and competed with an anti-GBM HSPG monoclonal antibody in ELISA. Furthermore, both antibodies recognized a molecule that migrated in SDS-PAGE as a smear and had a molecular mass of approximately 200–210 kD after deglycosylation. In immunoelectron microscopy, agrin showed a linear distribution along the GBM and was present throughout the width of the GBM. This was again different from perlecan, which was exclusively present on the endothelial side of the GBM and was distributed in a nonlinear manner. Quantitative ELISA showed that, compared with perlecan, the agrin-like GBM HSPG showed a sixfold higher molarity in crude glomerular extract. These results show that agrin is a major component of the GBM, indicating that it may play a role in renal ultrafiltration and cell matrix interaction.
T
Since the characterization of perlecan as an HSPG component of basement membranes (Murdoch et al. 1994), this compound has been studied extensively as a candidate factor in the development of albuminuria. Perlecan consists of five functional domains composing a core protein of 467 kD, carrying three heparan sulfate chains attached to the unique N-terminal domain (Kallunki and Tryggvason 1992; Murdoch et al. 1994). To date however, the involvement of perlecan in renal ultrafiltration is questionable. Well-defined monoclonal antibodies (MAbs) against the perlecan core protein (Murdoch et al. 1994; Couchman et al. 1995; Groffen et al. 1997) showed a clearly different distribution from antibodies directed against isolated GBM HSPG and epitopes on the heparan sulfate chains (Van den Heuvel et al. 1989,1995; Van den Born et al. 1994), indicating the presence of additional HSPG species in the GBM. A possible role for agrin in the kidney was suggested by the possibility of staining neuromuscular junctions (NMJs) with anti-GBM HSPG antibodies (Van Kuppevelt et al. 1992), the identification of agrin as a HSPG (Tsen et al. 1995), and the production of tryptic peptides from bovine tubular basement membrane HSPGs with a high sequence similarity compared with rat agrin (Hagen et al. 1993).
In rat and mouse, the agrin core protein has an approximate molecular mass of 210 kD (Rupp et al. 1991, 1992). Agrin induces the clustering of acetyl-choline receptors (AChRs) and aggregation of sodium channels during development of the neuromuscular junction (Glass et al. 1996; Kleinmann and Reichert 1996; Sharp and Caldwell 1996). The AC
Materials and Methods
Antibodies
Characteristics of the antibodies used are as follows. Antiagrin antiserum PAb 707 was raised in rabbit against full-length chick agrin from a recombinant source (Denzer et al. 1995). Mouse MAbs M215, M138, and rabbit antiserum K42 were raised against human HSPG isolated from the GBM (Van den Heuvel et al. 1989; Van den Born et al. 1994). These three MAbs identically stain human tissues in immunofluorescence. Antiserum K42 crossreacts with this antigen in rat and mouse tissues. The mouse MAb 95J10 is directed against a core protein epitope within Domains I and II of human perlecan (Groffen et al. 1997). Crossreactivity with various human basement membrane components was negligible, as determined by ELISA (Table 1), including collagen Type I from human placenta, collagen Type IV from human placenta, heparan sulfate, chondroitin sulfate B (Sigma; St Louis, MO), and fibronectin from human plasma (Boehringer; Mannheim, Germany). The mouse MAbs Agr-33 and Agr-131 (StressGen Biotechnologies; Victoria, Alberta, Canada) are directed against spatially separated epitopes on the core protein of rat agrin (Hoch et al. 1994). They do not crossreact in immunofluorescence on human tissues (unpublished observations).
Crossreactivity of MAbs 95J10, M215, and K42 with human basement membrane components, as determinedby ELISA a
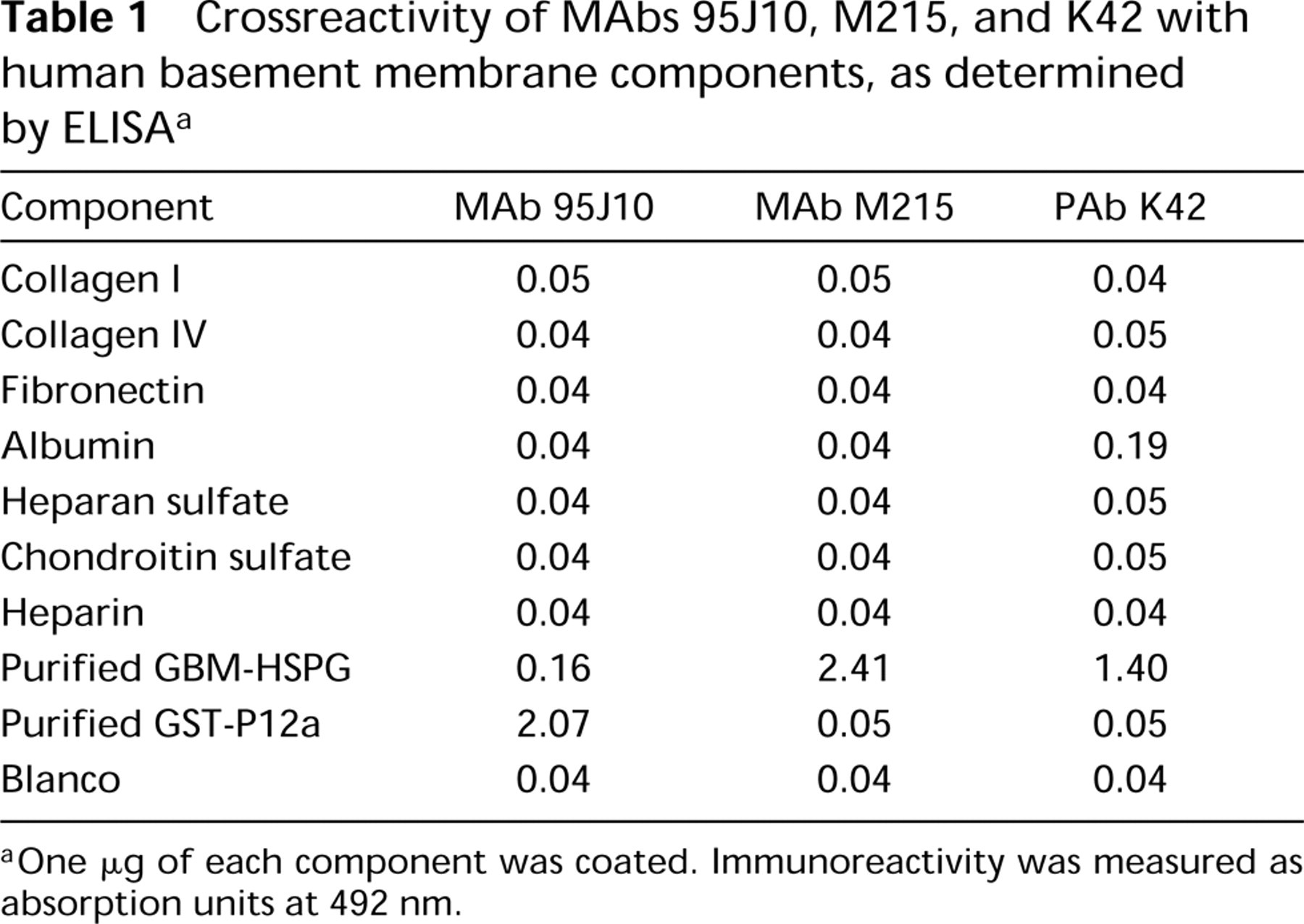
One μg of each component was coated. Immunoreactivity was measured as absorption units at 492 nm.
Indirect Immunofluorescence
Adult human kidney cortex tissue obtained from post-trauma donors was snap-frozen in polybrene, cooled with liquid nitrogen. Two-μm sections were stored at — 80C until use. Skeletal muscle tissue was obtained from adult rat hamstrings and frozen in polybrene immediately after dissection. Six-μm sections were prepared as above. Immunohistochem-istry was performed as described elsewhere (Van Kuppevelt et al. 1992), using FITC-conjugated goat anti-mouse and goat anti-rabbit as the secondary antibodies (Dakopatts; Copenhagen, Denmark). For identification of neuromuscular synapses in skeletal muscle sections, rhodamine-conju-gated α-bungarotoxin was added to the secondary antibody mixture at a final dilution of 1:400. In double immunofluorescence studies, MAbs were detected by goat anti-mouse immunoglobulins coupled to Texas Red (Sanbio; Uden, The Netherlands).
Immunoelectron Microscopy
Incubations were performed at 22C, unless otherwise indicated. Kidney cortex slices were immersed for 3 hr with PLP fixative (a mixture of periodate-lysine and 2% paraformaldehyde), rinsed with PBS (pH 7.4), cryoprotected in 2.3 M sucrose for 45 min, and snap-frozen in liquid nitrogen. Twenty-five-μm cryostat sections were incubated with PAb 707 diluted 1:100 in PBS containing 1% BSA for 18 hr at 4C and then washed three times for 30 min with PBS. This was also done with MAb 95J10 (diluted 1:5) and MAb M215 (diluted 1:10). After rinsing, the sections were incubated for 90 min with the appropriate peroxidase-conjugated secondary antibody (goat anti-rabbit immunoglobulins, Dakopatts; rabbit anti-mouse immunoglobulins, Bio
Preparation of Glomerular Extract
Glomeruli were isolated from human kidney cortex by the sieving method (Langeveld and Veerkamp 1981) and resus-pended in 20 volumes of 4 M guanidine HCl, 0.2% (w/v) Zwittergent 3–12, 50 mM sodium acetate, 10 mM EDTA, 10 mM N-ethylmaleimide, 10 mM 6-aminohexanoic acid, 5 mM 2-iodoacetamide, 1 mM phenylmethylsulfonyl fluoride, and 1 mM benzamidine HCl (pH 6.0). The suspension was incubated for 16 hr at 4C for extraction and centrifuged for 25 min at 10,000 × g to remove insoluble material.
Enzyme-linked Immunosorbent Assay
For quantitative determination of HSPGs by ELISA, crude glomerular extract was diluted 40-fold in coating buffer (Harlow and Lane 1988). Perlecan levels were measured with 95J10 as the primary antibody, using a recombinant fragment comprising Domains I and II of human perlecan (Groffen et al. 1996) as standard. In the case of M215, purified GBM HSPG (Van den Heuvel et al. 1989) was used as standard. To correct for the efficiency of coating, known amounts of the standards were added to the diluted extract before coating. Regression lines were computed for the calibration curves, which were linear functions of the amount of internal standard. The concentration of each HSPG in the crude glomerular extract was calculated as the average of five individual measurements.
For semiquantitative ELISA, wells were uniformly coated with 0.5 μg/well purified GBM HSPG (Van den Heuvel et al. 1989). Protein concentrations were determined according to the Bradford method with BSA as standard (Ausubel 1989). Competition ELISA was performed by using a mixture of PAb 707 (serial dilution) and MAb M138 (0.2 μg/ml) as the primary antibody. Subsequently, bound MAb was measured using a peroxidase-conjugated goat anti-mouse serum (1:2000; Dakopatts) as above.
Nitrous Acid Treatment and Immunoblotting
Five μg GBM HSPG (Van den Heuvel et al. 1989) was dissolved in 100 μl nitrous acid solution, freshly prepared by dissolving 5% (w/v) NaNO2 in 1.5 M hydrochloric acid. This mixture was incubated for 80 min at 22C. Subsequently, the core proteins were precipitated in the presence of 75% ethanol. SDS-PAGE and immunoblotting were carried out according to described procedures (Harlow et al. 1988). All samples were denatured before electrophoresis through 3-20% polyacrylamide, using Multimark as a molecular size marker (Novex; San Diego, CA).
Results
Immunofluorescence Microscopy
The presence of agrin in the glomerulus was investigated by indirect immunofluorescence. Cryosections of human renal cortex were stained with antibodies against agrin, perlecan, and “GBM-HSPG,” previously described HSPG isolated from the human GBM (Van den Heuvel et al. 1989). Combined staining with anti-agrin antiserum (Figure 1A) and MAb M215 against GBM-HSPG (Figure 1B) gave identical results. This consisted of strong linear staining of the GBM together with weak linear staining of tubular basement membranes. Similar results were obtained with four independent anti-agrin antisera, and all corresponding preimmune sera were negative (not shown). The observed staining pattern closely resembled that observed with the polyclonal antibody K42, directed against HSPG isolated from the GBM (Figure 1C). To confirm that the glomerular staining was not an artifact due to crossreactivity of the polyclonal antiserum, two MAbs, Agr-33 (Figure 1D) and Agr-131 (not shown) against rat agrin, were used to stain rat kidney cortex. Both antibodies produced strong staining of the GBM and very weak staining of the tubular basement membrane. In comparison with anti-agrin and anti-GBM-HSPG staining, the anti-perlecan antibody produced a clearly different staining pattern (Figure 1E). The above results indicate that agrin is present in the GBM.
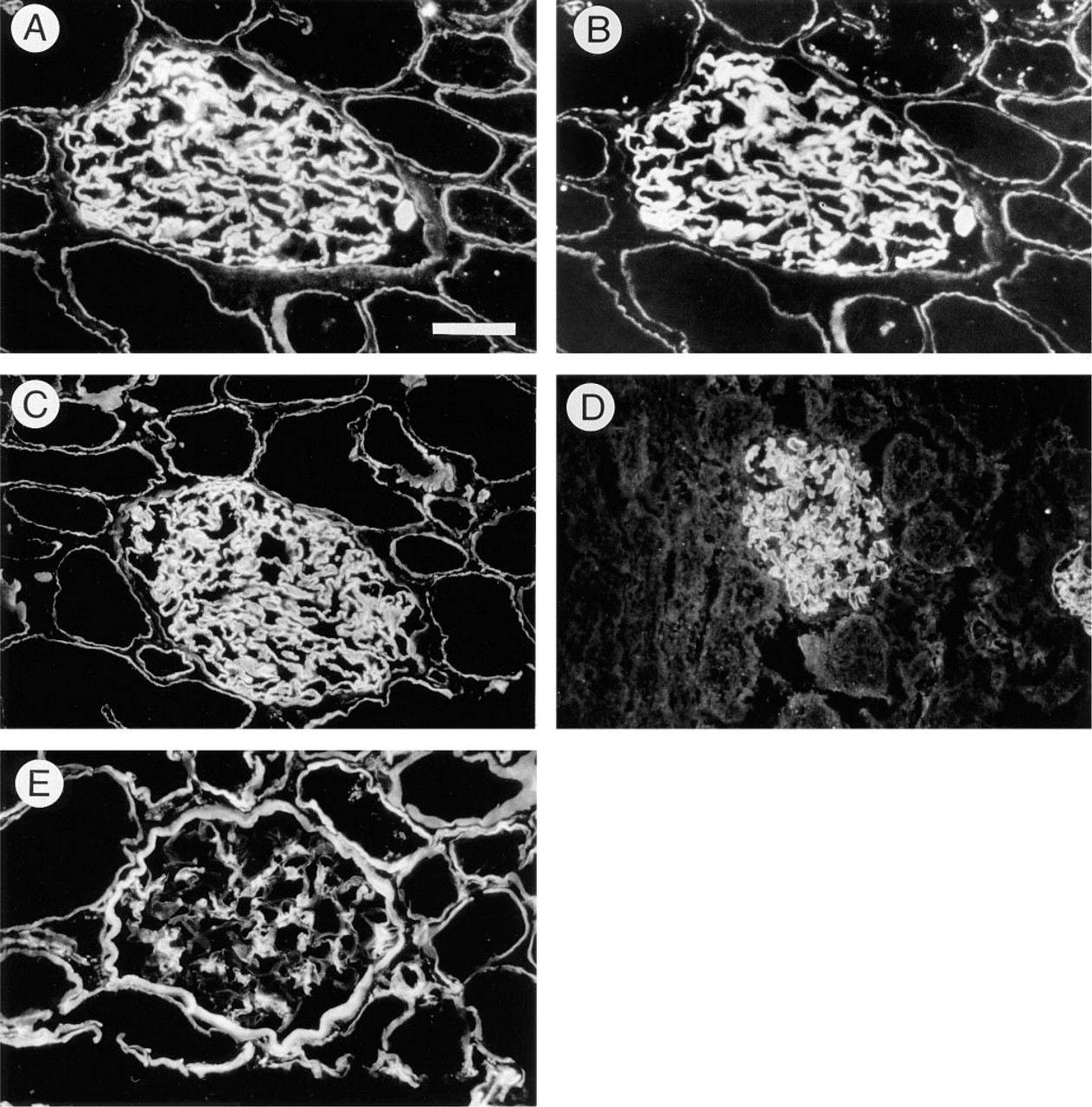
Indirect immunofluorescence staining of glomeruli with anti-HSPG antibodies. (A,B) Combined immunofluorescence staining of a single section with PAb 707 (against agrin, A) and MAb M215 (against the core protein of GBM-HSPG, B). Bar = 37.5 μm. (C) PAb K42 (against GBM-HSPG). (D) MAb Agr-33 (against the core protein of rat agrin). (E) MAb 95J10 (against perlecan). Sections were prepared from human (A-C,E) or rat (D) kidney cortex.
Because agrin is concentrated at the NMJ, we next compared staining patterns of anti-agrin and anti-GBM-HSPG antibodies in rat skeletal muscle sections. Junctions were identified by staining with rhodamine-α-bungarotoxin, recognizing the acetylcholine receptor clusters on the postsynaptic membrane (Figure 2A). Staining of the same section with MAb Agr-33 (directed against rat agrin) resulted in a strong signal localized at the synaptic cleft of the NMJ (Figure 2B). All junctions identified by rhodamine-α-bungarotoxin staining showed strong immunofluorescence. In addition, extrajunctional patches were observed that were strongly stained by Agr-33 but not by rhodamine-α-bungarotoxin. Furthermore, a weak staining of basement membranes surrounding the muscle fibers was present. Comparably, sections stained by rhodamine-α-bungarotoxin (Figure 2C) were immunostained with anti-GBM-HSPG antiserum K42 (Figure 2D). In agreement with Agr-33, the K42 antiserum stained all neuromuscular junctions together with some focal accumulations outside the junction and weak staining of the basement membrane that surrounds the muscle fibers.
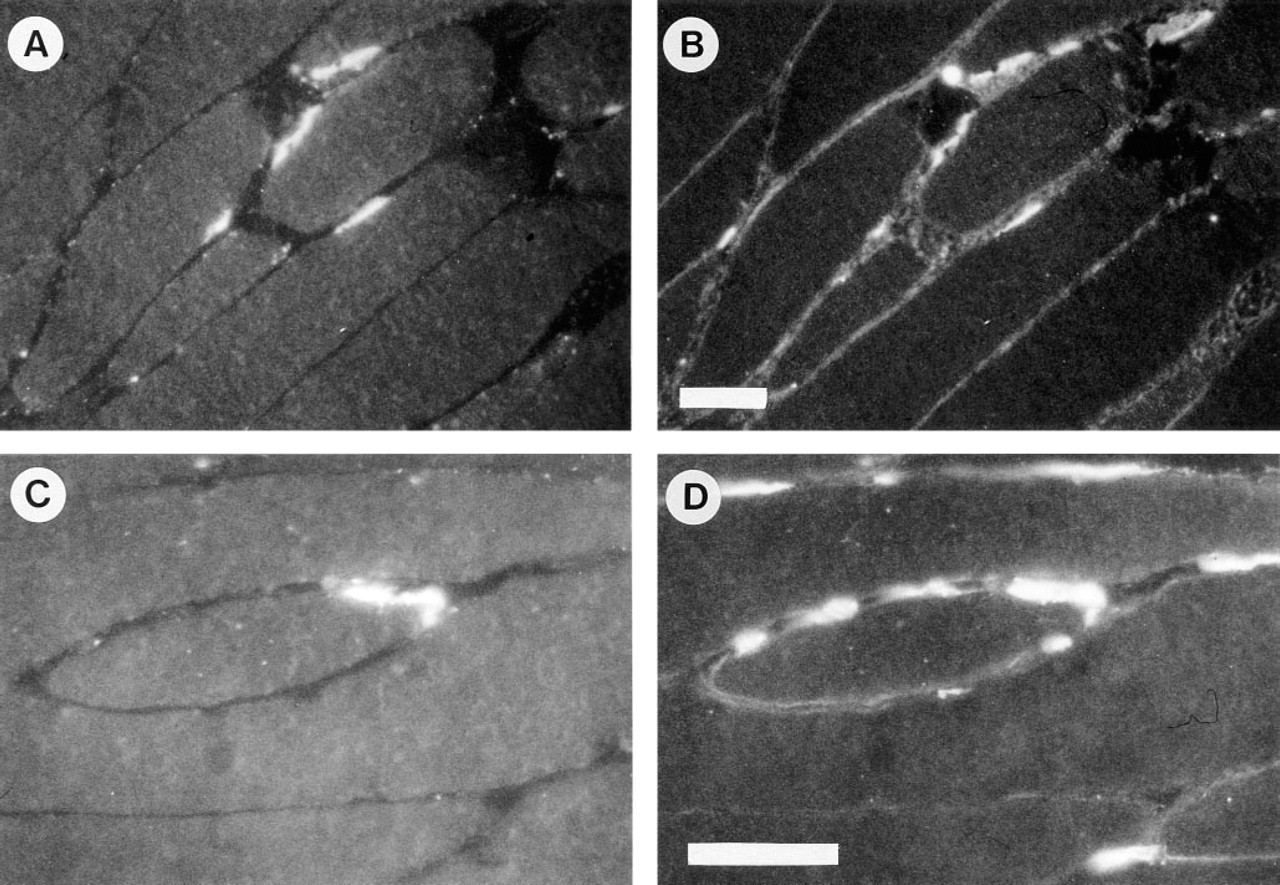
Distribution of agrin (top panel) and GBM-HSPG (bottom panel) in rat skeletal muscle. (
Immunoelectron Microscopy
To study the distribution of basement membrane HSPGs at the ultrastructural level, immunoelectron microscopy was performed on human kidney cortex. The distributions of agrin (top panel), perlecan (middle panel), and the GBM-HSPG recognized by M215 (bottom panel) are shown in Figure 3. With anti-agrin antiserum a strong linear staining was seen along the full length of the GBM (Figures 3A–3A). The entire width of the GBM was stained, but the signal was markedly stronger at both edges of the GBM. This localization at both the endothelial and the epithelial side of the GBM was confirmed by immunogold labeling (Figure 3C). We could also observe faint staining of the mesangial matrix. The anti-perlecan MAb 95J10 produced strong staining at the interface of the mesangium and the capillary endothelium (Figures 3D-3F). The overall GBM staining was mild and was restricted to the endothelial side in a nonlinear pattern. In addition, by immunogold labeling perlecan staining was found exclusively on the endothelial side of the GBM. With MAb M215, strong linear staining was observed throughout the entire length and width of the GBM (Figures 3G-3I). Immunogold labeling also demonstrated that the M215 epitope was present throughout the GBM. Occasionally, mild staining of the mesangial matrix was observed.
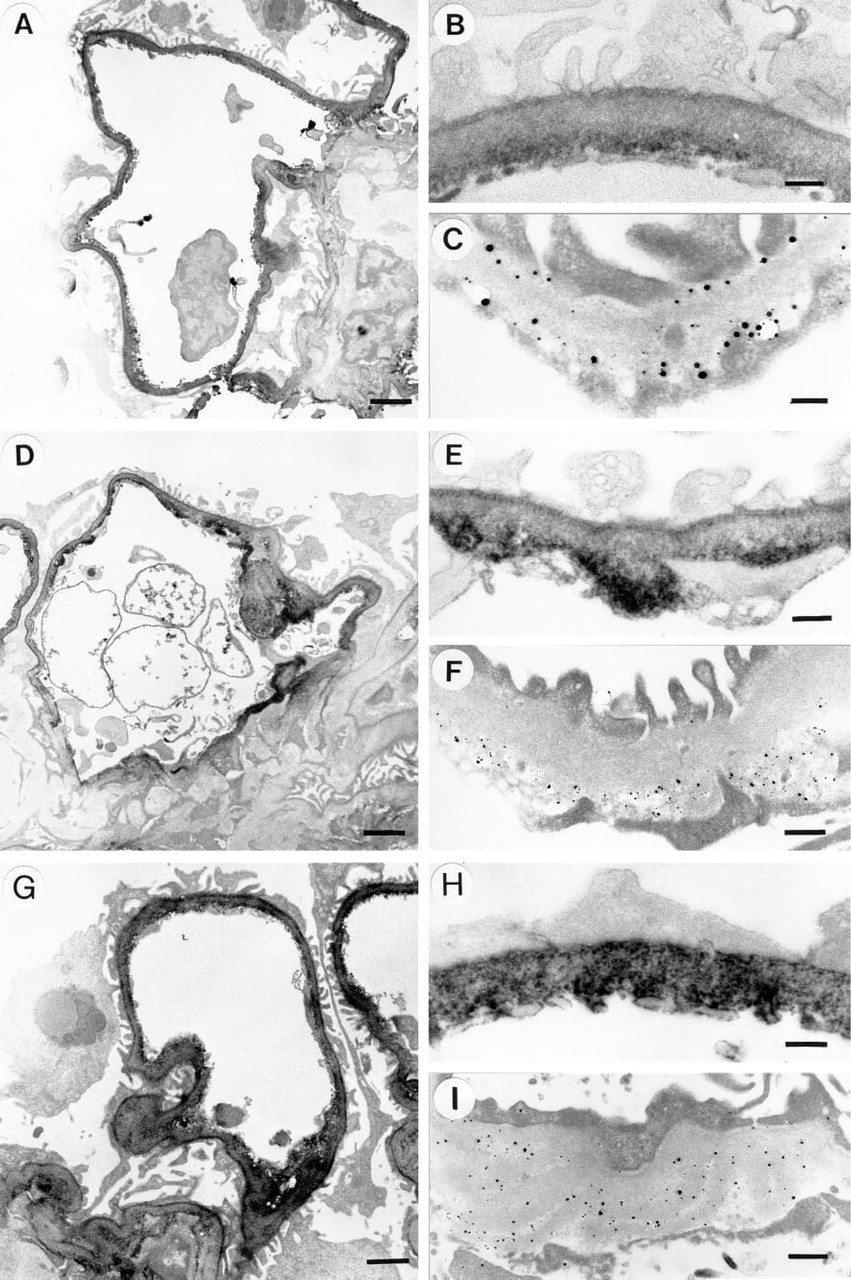
Immunoelectron microscopy of human glomeruli with anti-agrin antiserum 707 (
The linear distribution of the agrin-like HSPG along the GBM corresponds with the homogenous linear staining by immunofluorescence in this and previous studies (Figure 1B) (Van den Heuvel et al. 1989, 1995; Van den Born et al. 1992, 1994). The ultra-structural distribution of perlecan also corresponds with the staining of perlecan in immunofluorescence, characterized by discontinuous staining of the capillaries and relatively strong staining of the mesangium and Bowman's capsule (Figure 1E) (Hassel et al. 1980; Murdoch et al. 1994; Couchman et al. 1995).
Immunoreactivity of Anti-Agrin Antiserum with Purified GBM-HSPG
The described results show a co-localization of agrin-like and GBM-HSPG-like immunoreactivity in both the GBM and the NMJ. Given the unknown identity of the purified GBM-HSPG and the identification of agrin as a HSPG, we asked whether both proteogly-cans could be identical. To address this, isolated GBM-HSPG was coated into microtiter wells. Antisera raised against agrin showed a strong reaction with this proteoglycan, whereas the corresponding preimmune sera were negative (Figure 4, open squares and circles). In addition, we tested the capability of anti-agrin antiserum to compete in ELISA with four different MAbs against human GBM-HSPG (including M215 and M138, all recognizing the same core protein). The binding of three MAbs was not blocked. Because the anti-agrin antiserum was raised against chick agrin, this suggests that the corresponding epitopes of human agrin are not immunogenic or absent in chick agrin. However, the binding of M138 to native GBM-HSPG was inhibited by anti-agrin (Figure 4, solid squares). These results show that the isolated GBM-HSPG is recognized by both anti-GBM-HSPG and anti-agrin antibodies.
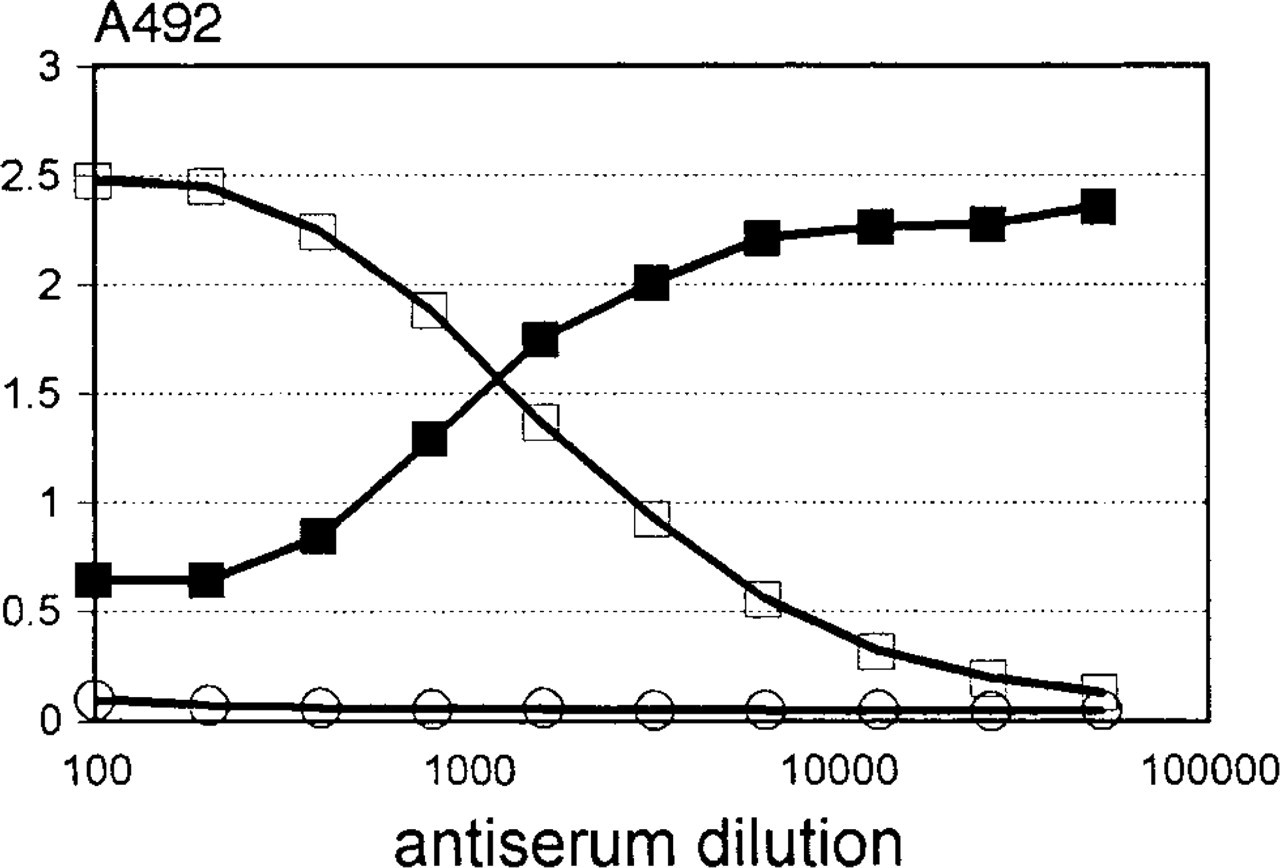
Recognition of GBM-HSPG by serially diluted anti-agrin antiserum (open squares) or preimmune serum (open circles) in ELISA. Similarly, binding of a mouse MAb M138 was measured in the presence of serially diluted anti-agrin antiserum as a competitor (solid squares).
Western blot analysis of isolated GBM-HSPG was performed with antibodies against agrin and GBM-HSPG (Figure 5). All antibodies recognized a large proteoglycan with a characteristically smeared appearance and similar molecular mass. The antigen was sensitive to nitrous acid treatment, indicating the presence of heparan sulfate residues. The nitrous acid-treated core protein was recognized by all antibodies and displayed a molecular mass of approximately 200-210 kD. In addition to this prominent band, two additional bands were weakly stained by M215 and M138. These bands are presumably degradation products and could also be visualized with PAb 707 after prolonged exposure (not shown).
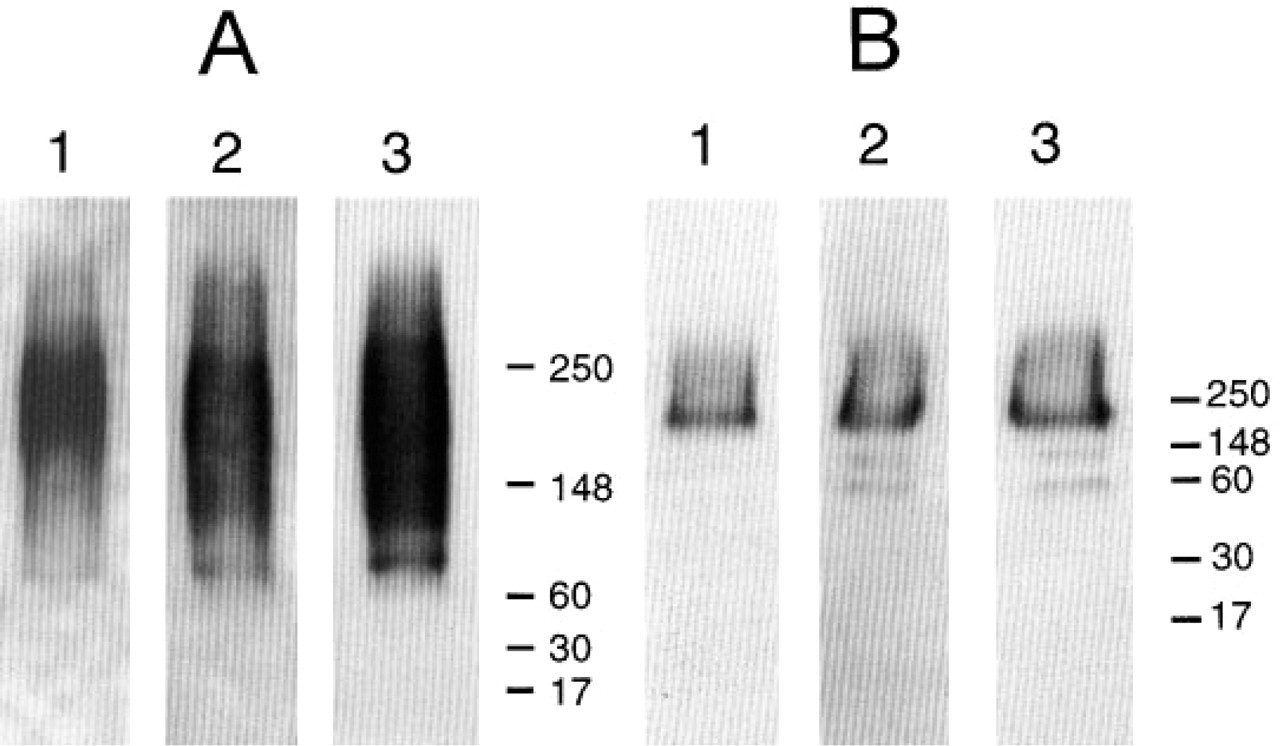
Western blot analysis of intact (
Quantitative Detection of HSPGs in Isolated Glomeruli
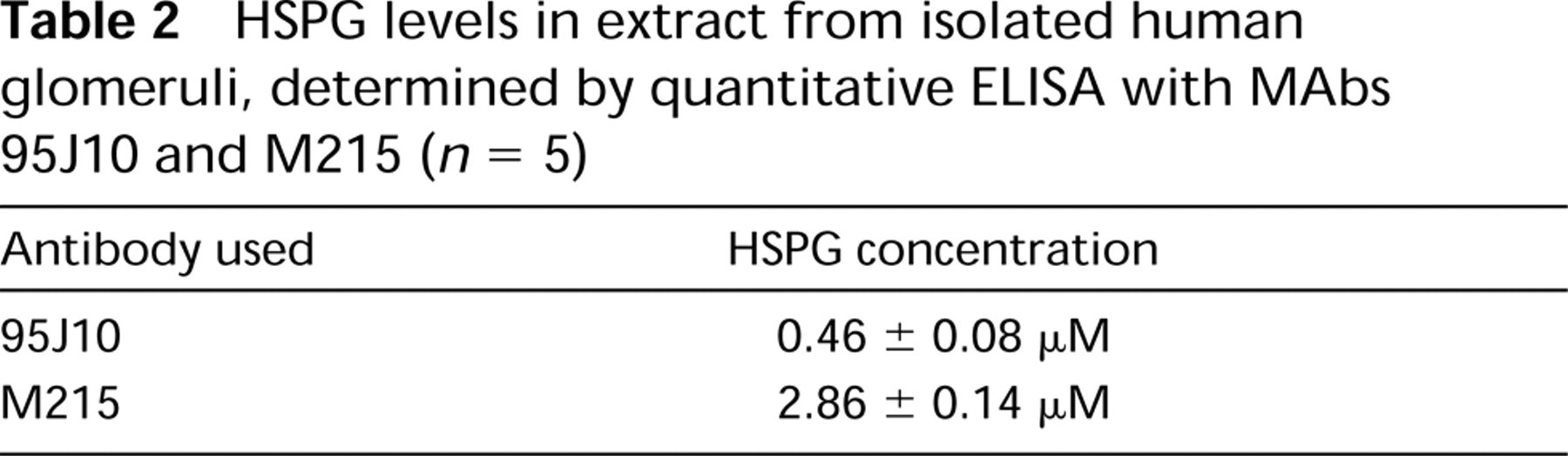
The strong staining of agrin in immunofluorescence and immunoelectron microscopy suggests that it is highly concentrated in the GBM. To determine the relative contribution of each molecule to the total HSPG content of the glomerulus, quantitative ELISA was performed with 95J10 and M215. A standard curve for 95J10 was constructed using an affinity-purified recombinant fragment comprising Domains I and II of human perlecan (Groffen et al. 1997). For M215, the purified GBM-HSPG preparation was used as standard. The results (Table 2) show that the agrin-like HSPG is present in sixfold molar excess over perlecan.
HSPG levels in extract from isolated human glomeruli, determined by quantitative ELISA with MAbs 95J10 and M215 (n = 5)
Discussion
In this study we have used an antiserum against chick agrin (Denzer et al. 1995) to investigate the presence of agrin in the human glomerulus. To this end, we first confirmed that the antiserum retains a high specificity for agrin in mouse, rat, and human tissues. Comparing the distribution of agrin and GBM-HSPG by immunofluorescence, we found an exact co-localization in both the glomerulus and the neuromuscular junction. Consistently, the anti-agrin antiserum recognized the HSPG isolated from the human GBM in a direct ELISA assay. The antiserum could also inhibit the binding of MAb M138 to GBM-HSPG in a competition ELISA assay. This demonstrates that one of the epitopes recognized by PAb 707 is identical or proximate to the M138 epitope, known to be located on the core protein (Van den Born et al. 1994). Finally, the estimated molecular mass of the core protein (200- 210 kD) corresponds to that of rat agrin (Rupp et al. 1991; Tsen et al. 1995), as shown by immunoblot analysis. These data suggest that the unidentified HSPG component of the GBM is identical to agrin. Nevertheless, small differences are seen by immuno-electron microscopy with PAb 707 and MAb M215. The anti-agrin antibody is particularly bound to the endothelial and epithelial side of the GBM, whereas M215 binding was observed throughout the width of the GBM. This difference, also confirmed by immu-nogold labeling, might be related to different access of both antibodies to the lamina densa. Alternatively, the corresponding epitopes may be shielded by interactions of agrin in the lamina densa.
The quantitation of the agrin-like GBM-HSPG and perlecan in extract from isolated glomeruli could be misleading in two ways. First, the standard used for quantitation by M215 cannot be considered absolutely contaminant-free, even though it was extensively purified (Van den Heuvel et al. 1989). Although no contaminants were observed by visual inspection of SDS-PAGE analyses, a trace amount of perlecan was detected by ELISA (Table 1). Likewise, the presence of trace amounts of other hypothetical HSPG species cannot be excluded. Secondly, the relative concentrations of both HSPGs in whole glomeruli are not necessarily identical to those in the GBM. Considering that perlecan is also present in Bowman's capsule (Figure 1E), the relative abundance of the agrin-like HSPG within the GBM is presumably even higher.
What functions might agrin serve in the human GBM? Although many aspects of the structure and function of agrin isoforms have been studied in detail, the presence of agrin in the mammalian kidney and GBM has not been previously described. One study reported the occurrence of agrin-like proteins in the tubular structures of chick kidney and showed that these molecules have only little AC
Second, agrin may provide a cytoskeletal link for the GBM through its interaction with α-dystroglycan (Deyst et al. 1995) and laminin (Denzer et al. 1997). Although α-dystroglycan does not appear to be the receptor involved in AC
A third possible function of agrin in the GBM is suggested by the presence of nine Kazal-type protease inhibitor domains (Rupp et al. 1991). Agrin inhibits the proteases trypsin, chymotrypsin, and plasmin (Biroc et al. 1993), and therefore it may stabilize the structure of the extracellular matrix.
In contrast to the early expression of agrin in synaptogenesis, the onset of expression in the GBM may be delayed to later stages of development. Agrin-defi-cient mutant mice show aberrations in the development of neuromuscular junctions and die shortly before birth. At this stage, no prominent kidney malformations were mentioned (Gautam et al. 1996). This relates to the abundance of agrin mRNA in the early development of the rat nervous system, in later stages (E15-P1) accompanied by lower but significant mRNA levels in the kidney (Stone and Nikolics 1995). Important alterations in proteoglycan composition are known to occur during development and maturation of the GBM (Couchman et al. 1990; Van den Heuvel et al. 1994). Agrin is likely to be one of the components involved in these changes.
From this study, we conclude that agrin is a major HSPG component of the human GBM. This finding has drastic implications for the investigation of structural and functional properties of the GBM. Changes in agrin structure and content may be important for glomerular function and may play a role in various types of glomerulopathy. The structure of the agrin isoform that is present in the GBM and its relevance for cell-matrix interactions are subjects for further investigation.
Footnotes
Acknowledgements
Supported by grant C93.1309 from the Dutch Kidney Foundation (AJG, CAB, LAM, JHV, LPH) and by grant #31-33697.92 from the Swiss National Science Foundation (MAR).