
Abstract
We used riboprobes and monoclonal antibodies to characterize tissue distribution of the human 550-kD homologue to gp330/megalin, primarily identified in the rat kidney. Human gp330/megalin mRNA and protein are readily identified in human parathyroid cells, placental cytotrophoblasts, kidney proximal tubule cells, and epididymal epithelial cells. The immunoreactivity is found on the surface of the cells and is heterogeneously downregulated in parathyroid hyperplasia and adenomas. Cells of the proximal kidney tubule and epididymis express the protein on their luminal aspect. Moreover, the protein is expressed in Type II pneumocytes, mammary epithelial and thyroid follicular cells, and the ciliary body of the eye. Sequence analysis of cDNA fragments, obtained by RT-PCR, revealed identical nucleotide sequences in parathyroid, kidney, placenta, epididymis, and lung. Immunohistochemistry for parathyroid hormone-related protein (PTHrP) revealed partial co-expression with human gp330/megalin in parathyroid, placenta, and mammary gland. The findings substantiate human gp330/megalin expression in a variety of human tissues expected to possess calcium-sensing functions. It may constitute a protein of utmost importance to adult and fetal calcium homeostasis, although other important functions may also be coupled to this exceptionally large protein with highly restricted tissue distribution.
Keywords
THE PARATHYROID glands exert an overall regulatory role in systemic calcium homeostasis by their unique ability to respond to changes in the extracellular Ca2+ concentration (Brown 1994). This has been related to the presence of Ca2+-sensing molecules on the parathyroid cell surface (Brown et al. 1993; Juhlin et al. 1987a,b) crucially involved in calcium regulation of parathyroid hormone (PTH) release (Gylfe et al. 1987; Nemeth and Scarpa 1987). Support for the presence of one such cell surface-bound Ca2+-sensing molecule was obtained when monoclonal anti-parathyroid antibodies were found to identify and interact with the Ca2+-sensing mechanism of these cells (Juhlin et al. 1987b). The antibodies were also reactive towards human cytotrophoblasts, which were subsequently demonstrated to exhibit Ca2+-sensing properties virtually identical to that of the parathyroid (Hellman et al. 1992; Juhlin et al. 1990). The antibodies also reacted with cells of the proximal kidney tubule (Juhlin et al. 1987a), and it was proposed that the recognized molecule might influence the cellular and systemic calcium homeostasis in multiple organs.
The antibody-reactive protein was isolated from human placenta and revealed as a 550-kD single-chain glycoprotein (Juhlin et al. 1990). Cloning of the cDNA and database search identified this protein as a human homologue to gp330/megalin, identical to the rat Heymann nephritis antigen (Lundgren et al. 1994; Saito et al. 1994; Raychowdhury et al. 1989). One suggested role of this antigen in the rat is in facilitating cellular uptake of certain apolipoproteins (Kounnas et al. 1995), but it has also been proposed as a calcium receptor of the kidney proximal tubule (Christensen et al. 1992). In a recent report on rat tissue distribution, the Heymann antigen has also been demonstrated in rat placenta, epididymis, thyroid, and Type II pneumocytes in the lung (Zheng et al. 1994). The tissue distribution is somewhat conspicuous, because it comprises several types of cells with presumed or demonstrated Ca2+-sensing ability (Brown 1991), which would be in accordance with our initial observation from parathyroid cells. Interestingly, many of these cells also express parathyroid hormone-related protein (PTHrP), and human cytotrophoblasts, rat pneumocytes, and a rat parathyroid cell line show signs of calcium-regulated PTHrP release (Hastings et al. 1995; Rubin et al. 1994; Hellman et al. 1990,1992; Sakaguchi et al. 1990).
The present investigation utilized immunohistochemistry, reverse transcription polymerase chain reaction (RT-PCR), and in situ hybridization to establish more closely the human tissue distribution of the 550-kD, putative Ca2+-sensing protein, and compared this expression with that of PTHrP.
Materials and Methods
Tissues
Pathological human parathyroid tissue was obtained during surgery for primary hyperparathyroidism (HPT). Biopsy specimens of normal parathyroid glands were acquired during thyroid surgery. Normal tissue specimens from kidney, epididymis, testis, mammary gland, adrenal gland, smooth and striated muscle, skin, liver, ileum, jejunum, colon, spleen, thyroid, pancreas, lymph node, eye, and lung were removed at surgery. Human placenta was acquired from uncomplicated pregnancies at full term. All tissue pieces intended for immunohistochemistry or RNA preparation were snap-frozen in liquid nitrogen and stored at −70C until processing. Cells intended for immunoprecipitation were prepared from parathyroid adenomatous and chief cell hyperplastic glands of patients with primary HPT, from human placenta, and from normal kidney as previously described (Rudberg et al. 1986). Cell viability routinely exceeded 95% by Trypan blue exclusion.
Monoclonal Antibodies
The production and characterization of the mouse monoclonal anti-parathyroid antibodies E11 and G11 have previously been described (Juhlin et al. 1987a). Briefly, these IgG MAbs recognize different epitopes of the same 550-kD protein and display a similar immunohistochemical reactivity (Juhlin et al. 1990). Moreover, G11, but not the E11 MAb, dose-dependently inhibits the steep increments in the cytoplasmic Ca2+ concentration ([Ca2+]i) and the suppression of PTH release that normally occur when human parathyroid cells are exposed to rising concentrations of extracellular Ca2+ (Hellman et al. 1992; Nygren et al. 1988; Juhlin et al. 1987b).
Immunostaining
Cryosections (6 μm) were fixed in acetone and exposed for 15 min to 1% H2O2 in PBS and 10% normal rabbit serum in PBS. For examination of human gp330/megalin expression, the sections were then incubated for 30 min with MAb E11 (5 μg/ml) in PBS containing 2% bovine serum albumin (BSA, fraction V; Sigma Chemical, St Louis, MO). A general finding with the MAb was that only acetone-fixed cryosectioned tissue could be used for immunohistochemistry. For example, the paraformaldehyde/glutaraldehyde-fixed specimens used for in situ hybridizations were uninformative with Mab E11. After exposure to a secondary rabbit anti-mouse antibody (Dakopatts; Glostrup, Denmark) for 30 min, the sections were treated with anti-mouse peroxidase-anti-peroxidase (PAP) complexes (Dakopatts). For investigation of PTHrP expression, sections were exposed to 10 μg/ml of a murine MAb recognizing human PTHrP(38–64) (Oncogene Science; New York, NY) followed by a biotinylated secondary rabbit anti-mouse antibody (Jackson Laboratories; West Grove, PA) and peroxidase-conjugated biotin (Vector; Los Angeles, CA). Staining was visualized in all sections with 3-amino-9-ethylcarbazol and counterstaining was performed with Mayer's hematoxylin. Negative controls included omission of the primary antibodies and substitution with mouse IgG1 fractions or an irrelevant murine monoclonal IgG MAb specific for collagen Type II.
In Situ Hybridization
Tissues intended for in situ hybridization were fixed for 16–20 hr on ice in 4% paraformaldehyde and 0.25% glutaraldehyde in 0.1 M sodium phosphate (pH 7.0). Dehydration by increasing concentrations of ethanol and xylene was followed by embedding in Histovax (Histolab Products; Gothenburg, Sweden) as described earlier (Schmeltzer et al. 1989). Pieces were sectioned at 6 μm and further treated with 2 μg/ml proteinase K (Cox and Goldberg 1988). Other samples were fixed at 4C in 4% paraformaldehyde for 1 hr, followed by 4% paraformaldehyde and 5% sucrose overnight. These pieces were then treated with 5% and 30% sucrose, followed by snap-freezing in liquid nitrogen and cryo-sectioning at 6 μm.
A 377-
Reverse Transcriptase-Polymerase Chain Reaction
Total RNA was prepared from parathyroid, placenta, kidney, epididymis, liver, jejunum, ileum, colon, and lung utilizing the acid phenol/chloroform method (Maniatis et al. 1982). First strand cDNA synthesis was subsequently performed in a total volume of 30 μl containing 50 mM Tris-HCl (pH 8.3), 75 mM KCl, 3 mM MgCl2, 3.33 mM DTT, 45 pmol oligo (dT)17 primer, 0.2 mM dNTP, approximately 2.6 μg of total RNA, 40 U Rnasine RNAse inhibitor (Promega; Madison, WI), and 24.5 U AMV-reversed transcriptase (Promega). Before the enzyme was added, all other components were incubated at 65C for 5 min and then put on ice. The samples were then incubated at 42C for 20 min followed by 40 min at 45C and finally at 65C for 20 min. A sample with the same components but lacking the AMV-reverse transcriptase served as a negative control. After completion of the reaction, 1/75 of the cDNA was used in the PCR.

DNA sequence of 1800 BP of the cDNA clone denoted pCAS-2 encoding the intracellular, transmembrane, and immediate extracellular portion of human gp330/megalin (Lundgren et al. 1994). 377 BP of this sequence was subcloned into pBluescript and used for production of riboprobes for in situ hybridization (underlined and marked in situ). Positions of the primers 5, 6, 1192, and 1193 are indicated.
Two pairs of primers were utilized (Figure 1). The 1192/1193 pair yielded a PCR product of 660
Immunoprecipitation and SDS-PAGE
A total of 2–5 × 106 parathyroid, placental, or renal cells were incubated for 30 min at 37C in 2 ml methionine-depleted DMEM containing 10% dialyzed fetal calf serum. [35S]-Methionine (0.5 mCi, >1000Ci/mmol; Amersham) was added, and the cells were labeled for 4 hr. The labeling was terminated by centrifugation, after which the cells were lysed on ice in 1 ml of 0.01 M Tris-HCl (pH 8.0), 4% Triton X-100, 2% aprotinin (Bayer; Leverkusen, Germany), 0.5 mM phenylmethylsulfonyl fluoride, and 5 mM benzamidine hydrochloride. The lysate was centrifuged at 35,000 X g for 1 hr at 4C and then subjected to antibody precipitation. Cell lysates were incubated with 10 μl normal rabbit serum over-right at 4C. Immunoglobulins were removed by the addition of protein A-Sepharose (50 μl; Pharmacia-UpJohn) and centrifugation. Aliquots of the supernatants were incubated for 4–12 hr with 1 μg MAb E11 or MAb G11. An MAb against a plant protein was used as a negative control. The immunoglobulins were recovered by exposure to protein A-Sepharose, washed with 0.01 M Tris-HCl (pH 8.0) containing 0.1% Triton X-100 and once with 0.01 M Tris-HCl (pH 8.0) containing 0.5 M NaCl and 0.1% Triton X-100. The immunoprecipitated proteins were eluted by heating at 95C in SDS loading buffer under reducing conditions. SDS-PAGE was carried out as described (Hellman et al. 1993) by use of 2.5–15% polyacrylamide gradient gels, which made a spacer gel superfluous. Radioactive proteins were detected by autoradiography on Kodak XAR film, and nonradioactive proteins by silver staining. Apo B lipoprotein (515 kD), myosin (205 kD), and phosphorylase B (97 kD), as well as human gp330/megalin protein purified from human placenta (Juhlin et al. 1990), were run in parallel as molecular weight standards.
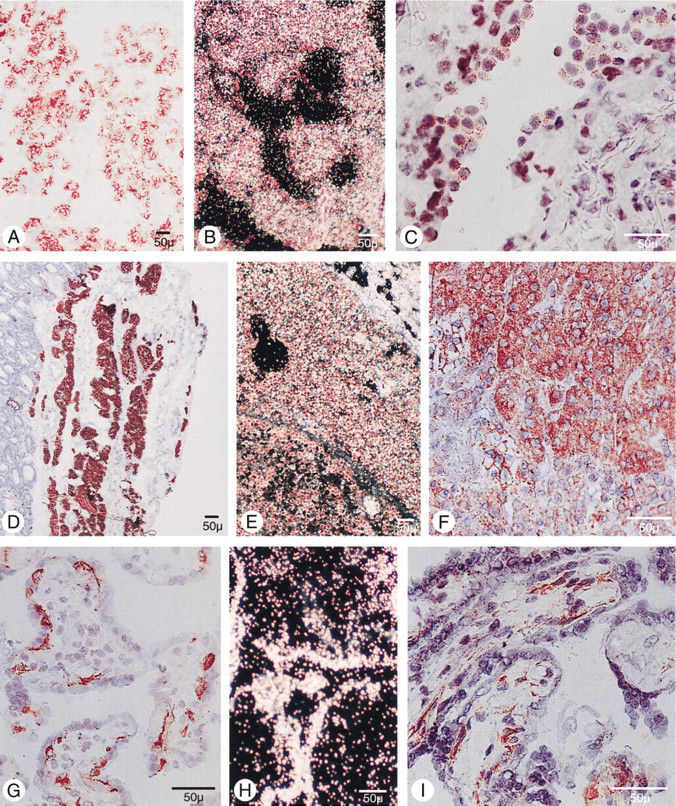
Immunohistochemical staining using the peroxidase-anti-peroxidase technique (
Results
Immunohistochemistry
The chief cells of normal human parathyroid glands demonstrated intense cell surface staining with MAb E11 (Figure 2 A). Pathological parathyroid tissue invariably showed reduced and often irregularly distributed immunoreactivity (Figure 2 d) (Juhlin et al. 1988). The reduced staining was confined to the pathological areas of parathyroid adenomas, whereas the rim of normal tissue outside the adenoma capsule displayed the same intense staining as the normal parathyroid glands (Figure 2 d). In nodularly hyperplastic glands of primary HPT, the more extensively expanding nodules demonstrated less intense E11 staining than the surrounding areas of more normal-appearing cells. Oxyphil parathyroid cells of normal and abnormal glands showed very weak or absent E11 staining.
Human placental tissue obtained at term demonstrated intense E11 staining, which was confined only to the cytotrophoblasts (Figure 2 G). Although some intracellular staining was present, the most intense staining was seen on the surface, as previously described (Hellman et al. 1992). A population of cells with characteristics of Type II pneumocytes in alveolar linings demonstrated positive staining (Figure 2 C). In human kidney specimens (Figure 3 A), intense staining was revealed within the brush border membrane of the proximal tubule cells as well as in cells of the parietal layer, preferentially at the urinary pole of Bowman's capsule. Glomeruli, including the juxtaglomerular apparatus/macula densa area, and recognizable cells in the distal tubule were negative. Follicular epithelial cells in normal thyroid glands were faintly stained on the surface (Figure 3 C), and the caput epididymis demonstrated intense staining predominantly on the luminal side of epithelial cells of the ductus epididymis (Figure 3 D) and rete testis, whereas epithelial cells in the corpus and cauda were negative. Moreover, epithelial cells in mammary gland ducts were positive (Figure 3 H), and cells in the choroid layer of the ciliary body of the eye, as well as scattered cells at the inner aspect of the retina, also reacted with the MAb E11 (Figure 3 I). Human liver, spleen, testis, lymph node, smooth and striated muscle, skin, adrenal gland, and pancreas were unreactive with MAb E11. Immunohistochemical staining of the small and large intestine with MAb E11 was difficult to interpret because of intense mucosal background staining, despite serious efforts to overcome this problem by the application of various protocols.
PTHrP expression was demonstrated in several of the same cells as human gp330/megalin. Thus, human gp330/megalin and PTHrP were co-expressed in parathyroid chief cells, placental cytotrophoblasts, and mammary duct epithelial cells. Although parathyroid chief cells from normal parathyroid glands were intensely positive for both proteins, there was no clear correlation between PTHrP and the downregulated human gp330/megalin expression of parathyroid adenoma (Figures 2D and 2F) or nodular hyperplasia. Apart from the human gp330/megalin-expressing placental cytotrophoblasts, there was also intense PTHrP reactivity within stromal cells of the placental villi (Figure 2 I). The ducts of the mammary gland showed intense PTHrP immunoreactivity, also seen with MAb E11 (Figure 3 G).
Intense PTHrP staining was also seen in epididymis, but in other areas than the human gp330/megalin immunostaining. The PTHrP staining was confined to the corpus and cauda, and generally failed to appear on the outermost layer of the epithelial lining (Figure 3 F). Kidney glomeruli were negative for PTHrP and weak staining was seen in proximal tubule cells, whereas intense reactivity was obvious in the collecting ducts (not shown). The only cells in the eye staining positive with the anti-PTHrP antibody were cells in the conjunctiva. Therefore, cells in the pigment epithelium were negative, as was the normal thyroid. The lung generally showed weak reactivity except for scattered cells in the alveolar lining.
In Situ Hybridization
The parathyroid tissues revealed variably intense hybridization with the anti-sense pCAS-2 probe (Figures 2B and 2E). Although some background activity was present with the sense probe, this usually caused no problems in interpreting the data. Parathyroid adenomas and nodules of hyperplastic glands displayed substantially lower anti-sense probe signal, whereas the normal parathyroid glands and the rim of normal tissue of parathyroid adenoma elicited considerably more intense and homogeneous staining. This was also true for some areas of diffuse hyperplasia, which surrounded the larger nodules of the primary hyperplastic glands.
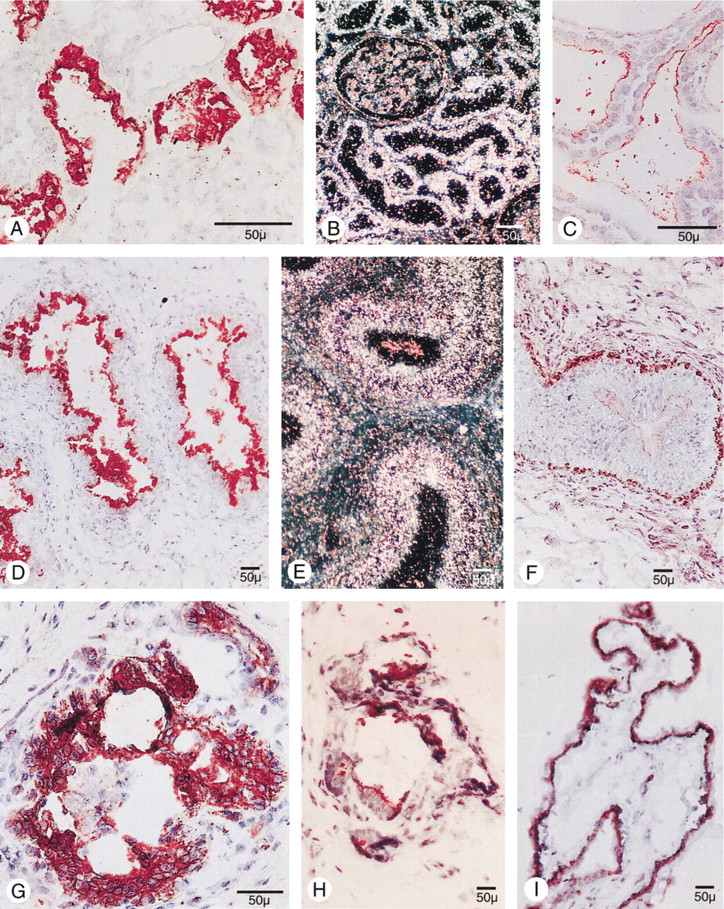
Immunohistochemical staining using the peroxidase-anti-peroxidase technique (
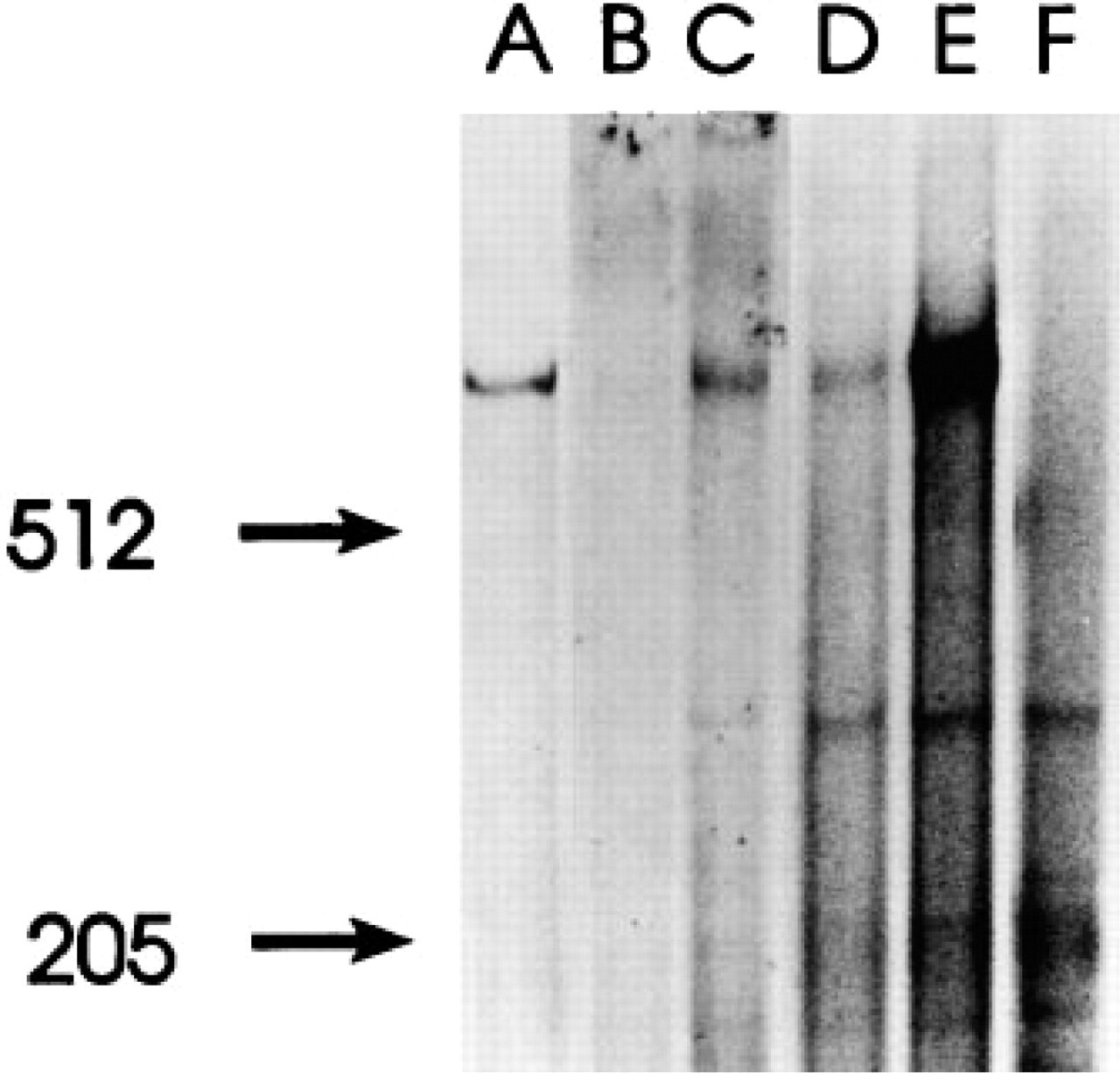
SDS-PAGE of lysates from [35S]-methionine-labeled human kidney (
In placenta, the anti-sense pCAS-2 probe signaled expression of human gp330/megalin mRNA in the cytotrophoblasts only, but the interpretation in the kidney was more difficult. However, after comparison with unspecific sense pCAS-2 signals as well as parallel sections stained with hematoxylin/eosin, it was evident that the human gp330/megalin mRNA was confined solely to proximal tubule cells in the kidney and was thus correlated with the immunohistochemical E11 staining. Epididymal epithelial cells in the caput (Figure 3 E) also demonstrated intense hybridization signals with the anti-sense riboprobe, and only low background with the sense RNA (not shown). No specific signal was detected in human lung. Moreover, in parallel to the findings at immunohistochemistry, in situ hybridization of small intestine and colon was difficult to interpret because of high background signals with the sense probe.
Polymerase Chain Reaction
Polymerase chain reaction of first strand cDNA from human parathyroid, epididymis, kidney, lung, and full term placenta revealed the expected bands of 660
Immunoprecipitation
The E11 and G11 antibodies precipitated a single protein with a similar size of approximately 550 kD from placenta, kidney, and parathyroid (Figure 4).
Discussion
Extracellular calcium is the predominant regulator of cytoplasmic calcium concentration ([Ca2+]i) in parathyroid cells (Gylfe et al. 1990). Several lines of evidence have suggested the presence of cell surface calcium sensor mechanisms crucially involved in the regulation of [Ca2+]i and PTH release in relation to extracellular Ca2+ concentrations (Gylfe et al. 1987; Nemeth and Scarpa 1987). By immunizing mice with human parathyroid cells, two monoclonal antibodies (denoted E11 and G11) recognizing cell surface structures on parathyroid, cytotrophoblasts, and kidney tubule cells were produced (Juhlin et al. 1987a,b). The 550-kD protein was further isolated from human placental tissue and partially sequenced with the aid of the E11 and G11 antibodies (Lundgren et al. 1994; Juhlin et al. 1990). In addition, MAb G11 was found to recognize an apparently functionally active epitope of this molecule, since exposure to G11 made parathyroid cells insensitive to changes in extracellular Ca2+ for both [Ca2+]i and PTH release (Juhlin et al. 1987b). Antibody-reactive cytotrophoblast cells of the human placenta were subsequently demonstrated to exhibit virtually parathyroid-identical regulation of [Ca2+]i in relation to changes in ambient Ca2+ (Hellman et al. 1992; Juhlin et al. 1990). These results led us to propose that the 550-kD protein may function as a Ca2+ sensor in these tissues (Hellman et al. 1992; Gylfe et al. 1990). Close sequence homology was demonstrated with the rat Heymann antigen, suggesting that the 550-kD protein is the human homologue. The Heymann nephritis antigen is a member of the LDL receptor superfamily and has previously attained considerable interest as a target molecule in experimental membranous glomerulonephritis in the rat (Raychowdhury et al. 1989). The complete amino acid sequence of this rat antigen, also denoted gp330 or megalin, has recently been published (Saito et al. 1994), confirming the revised size of this rat protein at approximately 500 kD (Christensen et al. 1992). Interestingly, analyses of the sequence and structure of rat and human gp330/megalin reveal Ca2+ binding sites in the extracellular domain (Lundgren et al. 1994).
In situ hybridization with the pCAS-2 anti-sense riboprobe closely corresponded to the immunohistochemical staining with the E11 and G11 antibodies. A concomitant decrease in human gp330/megalin mRNA and protein expression was demonstrated in the pathological parathyroid parenchyma of adenoma and hyperplasia, suggesting that the reduced gp330/megalin protein expression does not occur at the translational level. This decrease is compatible with the characteristic right shift in the set-point for PTH release and [Ca2+]i, due to decreased sensitivity to changes in ambient Ca2+, previously recognized as a characteristic feature of the pathological parathyroid tissue in all entities of HPT (Brown et al. 1990). The recently identified and cloned parathyroid calcium receptor (CaR), identified as a 120-kD seven transmembrane spanning G-protein-coupled receptor, is apparently also down-regulated in the pathological parenchyma of HPT (Kifor et al. 1996; Brown et al. 1993). Therefore, reduced expression of human gp330/megalin and CaR appears to be related to a deviant Ca2+-sensing function and hypercalcemia in HPT.
In situ hybridization of epididymis, kidney, and placenta also displayed concordance with the immunohistochemical investigations. In epididymis the signals were confined to the luminal side of the ductus epididymis as well as the rete testis, but no clear immunostaining was seen in visible sperm cells. The function of human gp330/megalin in epididymis is unclear, but it is intriguing that the protein is directed towards a fluid-containing space in this organ, as in the kidney, placenta, thyroid, eye, and the mammary gland. In the proximal tubule cells, gp330/megalin has been proposed to facilitate uptake of other proteins from the primary urine (Farquhar et al. 1994). Calcium appears to be needed for this uptake, and gp330/megalin is suggested to function as a Ca2+ receptor in this respect (Christensen et al. 1992). Other possible Ca2+-sensing functions may be involvement in the Ca2+ or PTH regulated 1-hydroxylation of 25(OH) vitamin D3, or in influencing Ca2+ reabsorption from the primary urine (Mathias and Brown, 1991). Placenta displayed intense gp330/megalin protein and mRNA expression only in cytotrophoblast cells, where previously a relation to PTHrP release has been proposed (Hellman et al. 1992). Human gp330/megalin expressed on the fetal aspect of the placental membrane may sense the fetal Ca2+ concentration and thereby regulate PTHrP release from the placenta. PTHrP, in turn, could promote transport of Ca2+ from the mother to the fetus and thereby participate in maintenance of the Ca2+ gradient over the placental membrane (Rodda et al. 1988). In addition, in the present study expression of PTHrP was detected in cytotrophoblasts.
In the thyroid gland, weaker expression of human gp330/megalin was detected within follicular cells, where PTHrP appeared absent. In the human eye, clear expression of gp330/megalin was seen in the choroid layer in the ciliary body but also stretched into the corresponding layer of retina. These cells produce the vitreous humor and are proposed to be equipped with proteins regulating the Ca2+ concentration in this liquid (Johnson et al. 1995). Epithelial cells of ducts in the mammary gland were intensely reactive with antibodies against human gp330/megalin and PTHrP. During lactation high amounts of Ca2+ and PTHrP are released with the milk (Budayr et al. 1989), and we can speculate that these cells and their possible Ca2+ sensing may be involved in maintaining an appropriate Ca2+ gradient or PTHrP concentration. Indeed, other proposed functions of gp330/megalin, such as apolipoprotein uptake, may well occur in the fat-containing milk.
Human gp330/megalin immunostaining was also detected in human lung tissue and the nucleotide sequence was at least partially identified. Although we were unable to achieve specific in situ hybridizations with human lung tissue, the PCR analysis and immunohistochemistry substantiate previous findings of gp330/megalin expression in Type II pneumocytes. In these cells, calcium has been shown to stimulate PTHrP and phospholipid secretion (Hastings et al. 1995; Rubin et al. 1994), and the present results confirmed the presence of PTHrP in human lung, although apparently in low amounts.
In virtually all tissues expressing human gp330/megalin, the protein is located on the cell surface, ideal for a receptor function. In contrast, hithertho published and unpublished data have shown that malignant tumors such as parathyroid carcinoma, choriocarcinoma (e.g., JEG-3 cells), renal cancer, and pulmonary adenocarcinoma display a markedly different and mainly intracellular localization of the protein (Hellman et al. 1993; Juhlin et al. 1989). The reason for this deviant distribution is unclear but may possibly relate not only to gp330/megalin itself but also to abnormal expression or function of RAP, the 44-kD receptor-associated protein, which has been proposed to act as a chaperone molecule involved in trafficking of gp330/megalin to the cell membrane (Willnow et al. 1996; Williams et al. 1994).
The results in this study indicate that only one form of gp330/megalin is expressed throughout the body, since the E11 antibody precipitated only one protein version from the investigated tissues and the PCR analyses demonstrated identical sequences. Furthermore, Northern blots of total RNA from human parathyroid, kidney, epididymis, and placenta have earlier shown only one species of hybridizing mRNA (Lundgren et al. 1994).
Evidence obtained thus far strongly suggests the involvement of CaR, and possibly also human gp330/megalin, in sensing of extracellular Ca2+ concentrations and thereby in the regulation of [Ca2+]i and PTH release in the parathyroid gland. Although available evidence can be considered circumstantial, functional studies appear to strongly suggest that human gp330/megalin is involved in sensing of extracellular Ca2+ concentrations and regulation of [Ca2+]i also in placental cytotrophoblasts (Hellman et al. 1992), as well as in a cell line originating from the proximal tubule (Hellman et al., unpublished results).
In conclusion, the distribution of human gp330/megalin is demonstrated in a number of tissues claimed to possess calcium sensing mechanisms (Brown 1994). However, it appears likely that several different proteins may function as Ca2+ receptors mediating the various effects seen by extracellular Ca2+. This also includes cells not involved in the regulation of systemic Ca2+ concentration but in which the local Ca2+ concentration varies, regulating specific cellular functions, such as the Type II pneumocytes of the lung, mammary gland, epididymis, and the eye.