
Abstract
Pro-thyrotropin-releasing hormone (pro-TRH) has been shown to be present throughout the central nervous system and in several peripheral tissues. In adrenals, TRH immunoreactivity has been reported but not characterized. We show here that two rat pro-TRH-derived peptides, TRH and prepro-TRH[160–169] (Ps4), were detected in extracts of rat adrenal glands by enzyme immunoassay. Endogenous TRH and Ps4 were purified by gel exclusion chromatography and reverse-phase HPLC. Structural identification of each peptide was achieved by chromatographic comparison with synthetic standards. By using the indirect immunofluorescence technique, TRH-immunoreactive cell bodies were found rather widely scattered outside the adrenal, in the brown adipose tissue in which the gland is embedded. These immunofluorescent cells have the typical appearance of mast cells and are metachromatic after histological staining with acidic Toluidine Blue. Our findings suggest that pro-TRH-derived peptides exist in rat mast cells.
T
Materials and Methods
Enzyme Immunoassay (EIA) Procedures
TRH EIA was performed as previously described (Montagne et al. 1996). Ps4 was measured by EIA using an enzymatic tracer kindly provided by Drs. C. Creminon and P. Pradelles (CEA; Saclay France). Briefly, the enzymatic tracer was obtained by covalent coupling of the N-terminal amine of Ps4 with the pure tetrameric G4 form of acetyl cholinesterase. Second antibody solid-phase EIA of Ps4 was performed in 96-well microtiter plates coated with mouse monoclonal anti-rabbit immunoglobulins to ensure separation between bound and free moieties of the tracer. The assay was carried out in a volume of 150 μl assay buffer (0.1 M potassium phosphate, pH 7.4, containing 0.4 M NaCl, 1 mM EDTA, and 0.1% BSA). A mixture of 50 μl diluted enzymatic tracer (1:200), 50 μl diluted anti-Ps4 antiserum (lot 371–1406; 1:40,000) and 50 μl standard or biological extract was incubated for 18 hr at 4C. After washing, the enzymatic activity in each well was measured using Ellman's method. The Ps4 EIA system, using Ps4 as standard and Ps4 coupled to acetylcholinesterase as an enzymatic tracer, recognized authentic Ps4 (detection limit 80%; B/Bo = 6 fmol).
Chromatographic Separation
Sixteen adult male Sprague-Dawley rats (Dépré; Saint Doulchard, France) weighing 300 g were sacrificed by decapitation and their whole adrenal glands were immediately removed. Tissues were immersed in 10% acetic acid at 95C for 10 min and then homogenized and extracted at 4C using a polytron homogeneizer. The resulting suspension was centrifuged at 3000 x g at 4C for 30 min and lyophilized. Crude adrenal extracts were first filtered through a Sephadex G-50 (fine) column at a flow rate of 8 ml/hr. For HPLC studies, the prepurified sample was dissolved in 200 μl of water plus 0.1% trifluoroacetic acid (TFA) and applied to 5-μm Lichrospher C-18 reverse-phase column (3.9 × 250 mm; Interchrom, Asnières, France) equilibrated with the same solvent at a flow rate of 0.75 ml/min. Three min after injection the bound material was eluted with a linear gradient (1%/3 min) of acetonitrile (0.07% TFA) in 0.1% TFA/water. Fractions of 750 μl were collected and lyophilized. Aliquots of each fraction were reconstituted in EIA buffer and assayed for TRH or Ps4 immunoreactivity.
Histology and Immunocytochemical Procedure
Two rats were perfused intracardiacally with heparinized saline (100 ml) and then with an infusion of 200 ml of McLean's fixative containing 2.5 mM sodium m-periodate, 18.7 mM
Results
To detect the presence of peptides derived from pro-TRH in rat adrenal glands, tissue extract was subjected to size fractionation using Sephadex G-50 and reverse-phase HPLC analysis. Fractions were analyzed by EIA with two antisera directed against pro-TRH-derived products. By use of a TRH antiserum, a sharp immunoreactive peak was detected that emerged from the total volume of the column (Figure 1A). Application of the Ps4 EIA in the same procedure demonstrated the presence of immunoreactive material (Figure 1B). The predominant product co-eluted with synthetic Ps4 during gel filtration. To characterize more precisely the major pro-TRH-related products from adrenal glands, immunoreactive fractions 57–60 corresponding to the most immunoreactive fractions for either TRH or Ps4 EIA during gel filtration were pooled and subjected to reverse-phase HPLC on a 5-μm Lichrospher OD2 column with a very slow gradient of acetonitrile in water plus 0.1% TFA (Figure 1C). Under these conditions, the putative endogenous TRH from G-50 was fractionated into a major immunoreactive peak corresponding to TRH, which was accompanied by a minor trailing shoulder and two additional species. Aliquots of the fractions depicted in Figure 1C were also assayed for Ps4 immunoreactivity. Authentic Ps4 was recovered as a predominant immunoreactive species eluting in the same position as synthetic Ps4. Two additional and uncharacterized immunoreactive peaks were also detected in significant amounts.
Immunocytochemical localization of adrenal TRH-related peptides was examined in rats using specific anti-TRH immunoglobulins. Observation of the sections indicated that both the adrenal cortex and medulla were devoid of TRH immunoreactivity. At the light microscopic level, specific labeling was observed in scattered cells located in the brown adipose tissue (BAT) surrounding the adrenal glands (Figure 2A). These cells contained a distinguishable nucleus and were observed everywhere in BAT (Figure 2B), although most of them were proximal to the adrenal capsule. These cells were positively stained when sections were treated with Toluidine Blue (pH >2.5), which identifies mast cells. To determine if TRH-immunoreactive cells accounted for all of the adrenal mast cells, we first counted all of the fluorescent cells, then counterstained the sections with acidic Toluidine Blue. No additional metachromatic cells were evident within the tissue sections (Figures 2C–2E). With an Alcian blue/Safranin mixture, mast cells stained red, indicating that they correspond to typical connective tissue mast cells (data not shown). No fluorescence was observed with TRH-treated anti-TRH immunoglobulins (Figure 2F).
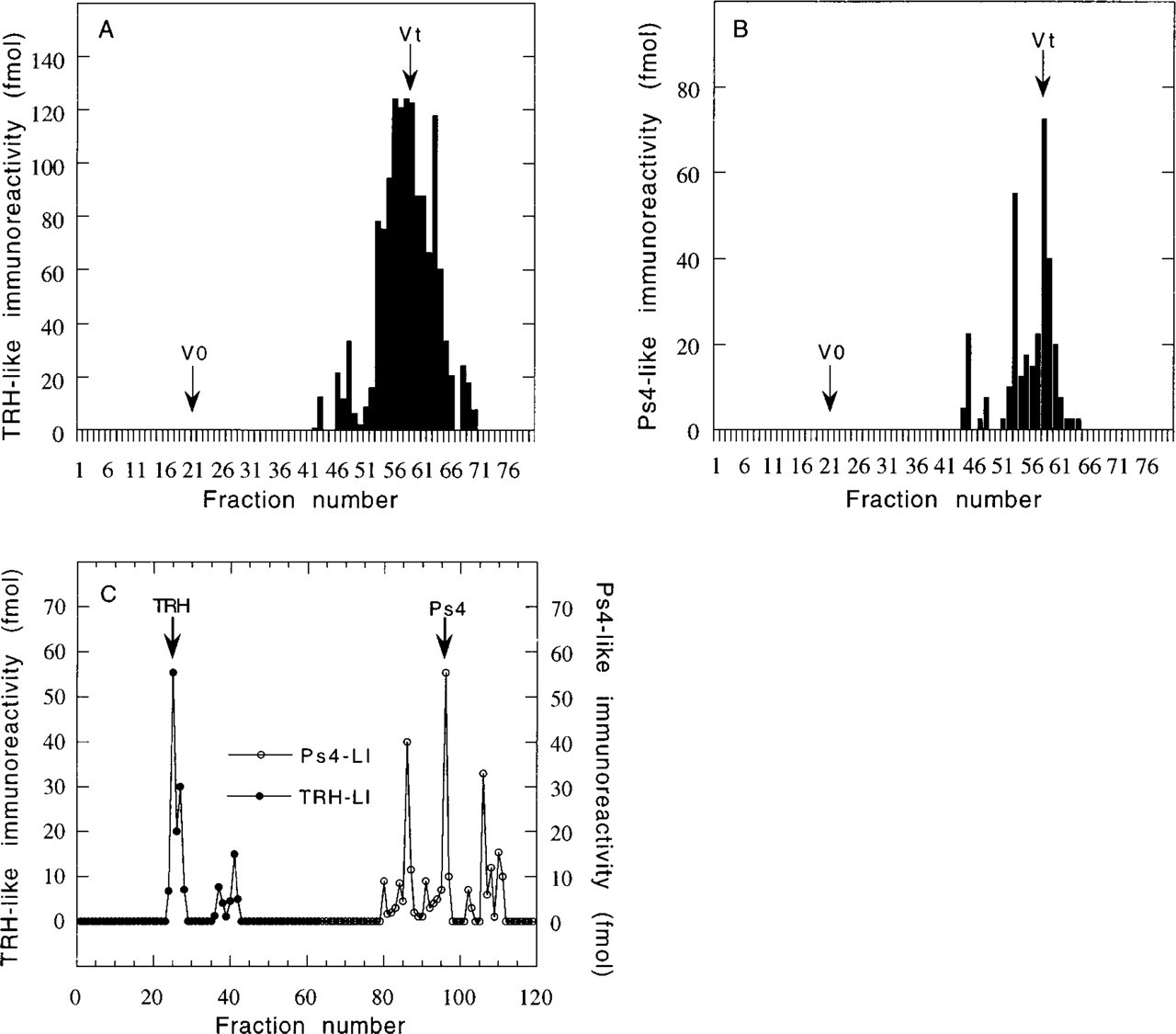
Fractionation by molecular sieve filtration and reverse-phase HPLC of TRH- and Ps4-immunoreactive peptides extracted from rat adrenal glands. (
Discussion
Several studies have indicated the presence of TRH in rat adrenal gland (Simard et al. 1989; Fuse et al. 1990), but its cellular localization and physiological significance were unknown. The present study demonstrates that adrenal TRH derives from a precursor molecule similar or closely related to hypothalamic pro-TRH. Despite the low level of TRH and Ps4 immunoreactivity (> 10 fmoles/adrenal glands), we have identified from 32 glands two forms that correspond to authentic TRH and Ps4. Connecting peptide Ps4 is a non-TRH peptide, which is yielded from the complete processing of the prohormone in the CNS. Several additional peaks of material immunoreactive in the homologous TRH and Ps4 EIA were also detected during chromatographic analysis of the adrenal gland extracts. These peaks were not further characterized because of the small amount available, but they may be derived from processing of pro-TRH in rat adrenal glands. Conversely, these immunoreactive peaks could comprise either proteolytic degradation products of pro-TRH or unidentified substances crossreacting in the assays. Taken together, these data indicate that Ps4 and TRH appear to be major endproducts. The cellular localization of TRH-immunoreactive peptides was obtained through immunocytochemical study using anti-TRH immunoglobulins. Intensive labeling was observed in scattered cells around the adrenal gland capsule, in BAT. These cells can be characterized as tissue basophils with Toluidine Blue and Alcian Blue. On the basis of the size and aspect of their nuclei, these basophils are typical mast cells, as previously described in the BAT (Mory et al. 1983). These data indicate that TRH-immunoreactive peptides are unequivocally localized in rat mast cells surrounding the adrenal glands. Moreover, the fact that absolutely no staining was seen anywhere else in the sections strongly suggests that authentic TRH, which is detected during HPLC separation as a major product, plays a great part in this signal. Preliminary PCR experiments in adrenal glands have permitted amplification of a 600-
The level of TRH expressed in the mast cells is probably too low to exert endocrine hormonal effects on distant tissues, but it appears adequate to support local paracrine or autocrine regulatory effects. Interestingly, it has been reported that TRH receptor mRNA is widely distributed in rat peripheral tissues and that a substantial amount of mRNA was observed in adrenal glands (Fukusumi et al. 1995). Identification of the cell type that expresses TRH receptor should enable identification of the role of local TRH.
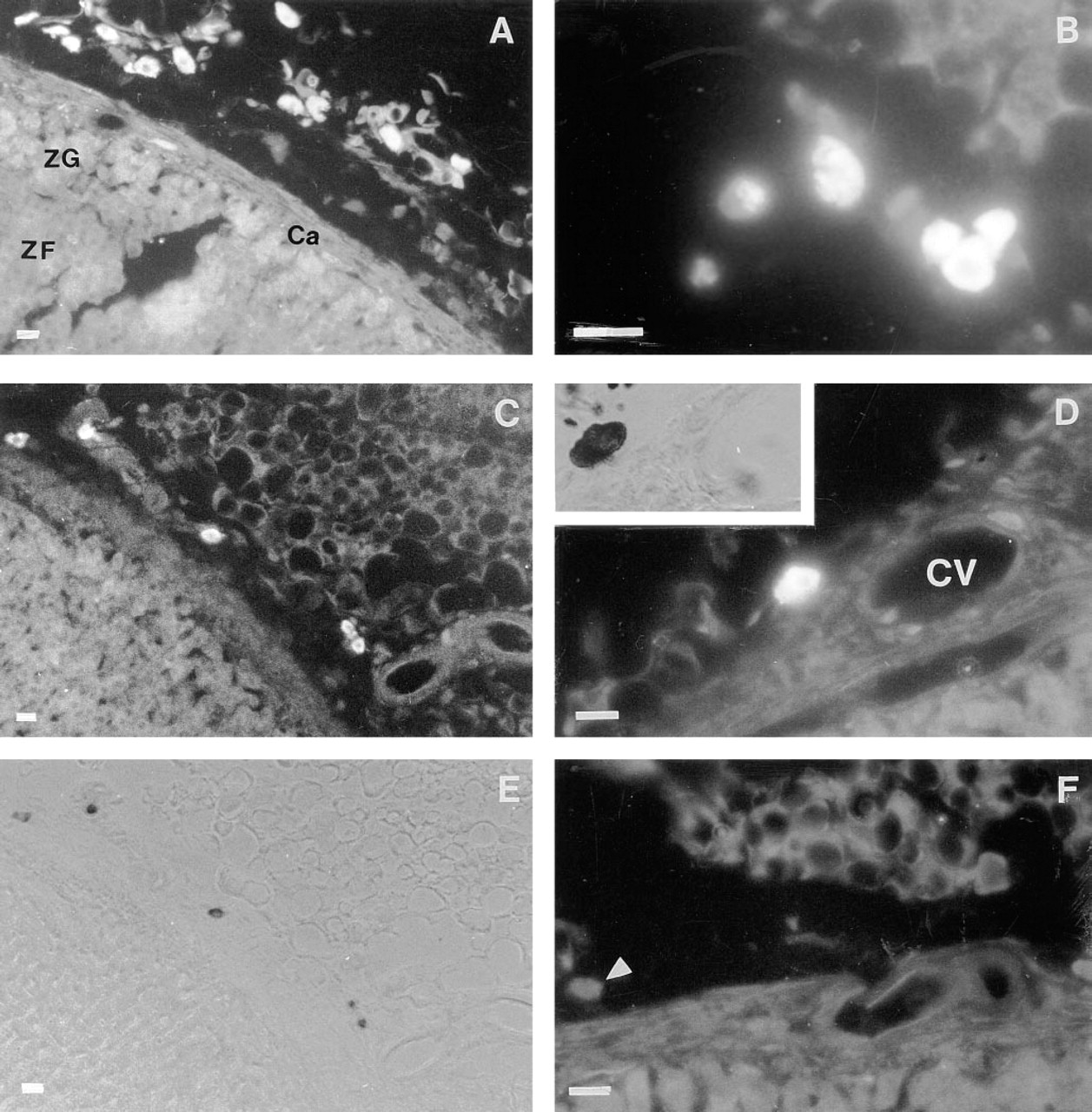
Immunocytochemical study demonstrating the presence of TRH immunoreactivity in mast cells. The mean (± SEM) number of mast cells in 8-μm transverse sections was 16.5 (± 2.6). (
Footnotes
Acknowledgements
We are grateful to Dr G. Mory for his pertinent comments.