
Abstract
The chemokine interleukin-8 (IL-8) mediates infiltration and adhesion of neutrophils during inflammatory processes. We have previously shown that this cytokine can be produced and released by normal and leukemic human mast cells (HMC-1 cells). To assess whether and to what extent this cytokine is stored intracellularly, we investigated production and localization of IL-8 at the single-cell level by combined use of flow cytometry (FACS) and immunoelectron microscopy. Conditions necessary for optimal fixation and permeabilization of HMC-1 cells were determined by measuring changes in cell-specific light scatter parameters and by estimating cellular uptake of propidiumiodide (PI). In this way, we were able to detect IL-8 with a monoclonal antibody in stimulated cells that were microwave-fixed with a combination of paraformaldehyde (4%) and glutaraldehyde (0.1%), followed by permeabilization with saponin (0.025%). FACS analysis revealed time-dependent synthesis of IL-8 with at most 50% positively stained cells at 8–12 hr after stimulation. For pre-embedding immunogold electron microscopy, cells were treated according to the protocol established by flow cytometry. IL-8 was found to be located in specific cytoplasmic, electron-dense granules of stimulated HMC-1 cells. These results confirm and extend our previous findings by demonstrating IL-8 expression in HMC-1 cells at the single-cell level. In addition, we propose that quantitative FACS can be reliably used in a timesaving manner to establish appropriate conditions for pre-embedding immunoelectron microscopy of intracellular antigens.
Keywords
THE CHEMOKINE INTERLEUKIN-8 (IL-8), initially described as a granulocyte-directed chemotactic stimulus, is a potent proinflammatory cytokine and has been identified in a number of inflammatory diseases including psoriasis, rheumatoid arthritis, and lung diseases (Matsushima and Oppenheim 1989). Although under these pathophysiological conditions increased numbers of mast cells were also observed (Toyry et al. 1988; Arnason and Malone 1995; Redington et al. 1995), their precise physiological function in the inflammatory process remains unknown. The ability of mast cells to produce IL-8 has been shown for cells located in the skin and in the lung as well as for the leukemic human mast cell line HMC-1 (Möller et al. 1993; Okayama et al. 1995). In HMC-1 cells and skin mast cells, the expression of IL-8 has been found to be inducible after stimulation of cells with the combination of PMA and calcium ionophore A23187 or by IgE/anti-IgE (Möller et al. 1993). In human lung mast cells a constitutive expression of IL-8 was ascertained that was not affected by anti-IgE challenge (Okayama et al. 1995). All these studies detected IL-8 at the level of mRNA and/or at the level of secreted protein, but did not take into account the amount possibly stored intracellularly in cell-specific granules, as described for a variety of mast cell-associated mediators (Galli 1993). Therefore, we decided to complete our investigations described recently (Möller et al. 1993) by estimation of time-dependent intracellular IL-8 synthesis in HMC-1 cells. For this purpose, flow cytometry would be an objective and easily quantifiable method and immunoelectron microscopic examination would provide information regarding the exact cytoplasmic localization of IL-8 in mast cells.
For intracellular detection of cytokines by these methods, particularly in mast cells, two major difficulties are encountered: (a) cytokines are soluble mediators that are easily eluted during specimen processing (Thorpe et al. 1992), and (b) the plasma membrane of mast cells reacts in a very sensitive fashion to detergents normally used for cell permeabilization (Tainsh et al. 1992; and the present study). Therefore, prefixation of cells is required to ensure sufficient stabilization of cellular integrity during detergent-induced membrane permeabilization. However, during fixation alterations of conformation-dependent epitopes of the cytokine investigated may occur (Sander et al. 1991). In general, paraformaldehyde (PFA), alcohol, or combinations of PFA with glutaraldehyde (GA) or lysine periodate (PLP) have been used for fixation of diverse cell types during immunofluorescence and pre-embedding immunocytochemistry specimen preparation (Sander et al. 1991; Priestley et al. 1992). Protocols applied for the permeabilization of plasma membranes depend on the cell type investigated and include a variety of detergents such as Triton X-100, saponin, lysolecithin, digitonin, or Tween-20 (Labalette–Houache et al. 1991; Sander et al. 1991; Kreft et al. 1992; Priestley et al. 1992; Hsi and Remick 1993). Because of this large variety of reagents, the sole use of immunoelectron microscopic techniques for the elaboration of optimal cell type-and cytokine-specific protocols is very time-consuming and no quantitative comparison between the different protocols applied is possible. In contrast, flow cytometric analysis allows the simultaneous determination of cell permeabilization by measuring propidium iodide (PI) uptake and of cellular integrity by measuring forward and sideward light scatter properties of cells. Shifts in forward and sideward scatter properties represent very sensitive indicators for altered cell size and granular composition, respectively (Lovett et al. 1984).
In addition to these applications, flow cytometry enables quantification of antibody binding under various conditions of cell fixation by use of a fluorescence-labeled secondary antibody. The successful application of flow cytometry for the detection of cell-associated cytokines has been demonstrated predominantly for mononuclear leukocytes (Sander et al. 1991; de Caestecker et al. 1992; Jung et al. 1993).
In this report we present a semiquantitative flow cytometric assay to detect intracellular IL-8 in HMC-1 cells at the single-cell level. Furthermore, we offer a strategic protocol established by flow cytometry that allowed rapid optimization of conditions necessary for fixation and permeabilization of mast cells before sub-cellular localization of IL-8 by pre-embedding immunoelectron microscopy. We suggest that this procedure is applicable to other cells and intracellular antigens.
Materials and Methods
Cell Culture
HMC-1 cells (kindly provided by Dr. Butterfield; Rochester, MN), which are immature human leukemic mast cells (Butterfield et al. 1988; Hamann et al. 1994), were cultured in Iscove's medium (Seromed; Berlin, Germany), supplemented with 10% FCS (Seromed), 10 μM monothiolglycerol (Sigma; Deisenhofen, Germany), and antibiotics (streptomycin and penicillin). Stimulation of HMC-1 cells was done in 24-multiwell plates (Falcon; Lincoln Park, NJ), as previously described (Möller et al. 1993). Briefly, 2 × 106 cells washed in PBS without magnesium and calcium were incubated for 8 hr in serum-free Iscove's medium (calcium concentration 1 mM) containing 25 ng/ml PMA (Sigma) and/or 250 nM calcium ionophore A23187 (Sigma).
Cell Fixation
The following fixatives were compared: 1% PFA; 4% PFA; 1% PFA/0.1% GA; 4% PFA/0.1% GA; 4% PFA/2% GA; and 2% PFA/10 mM lysine periodate (PLP). PFA (Merck; Darmstadt, Germany) was freshly prepared and diluted in PBS without magnesium and calcium. PLP was also prepared just before use according to the method of McLean and Nakane (1974). Fixation was done in a conventional temperature-regulated microwave oven with some modifications, as described by Jamur et al. (1995). Briefly, cells were washed, re-suspended in 10 ml fixative, and irradiated at 700 W for nearly 15–20 sec, elevating the temperature of the solution to 40 ± 5C. After fixation the cell suspension was immediately diluted with PBS, centrifuged, and washed twice with PBS.
Cell Permeabilization
Cells were permeabilized with increasing concentrations of Triton X-100 (Sigma) or saponin (Sigma), as indicated. Permeabilization with Triton X-100 (0.02–0.05%) was done for 20 min at 4C before antibody staining. Saponin treatment (0.003–0.05%) was done during the entire staining and washing procedure. The efficiency of permeabilization was controlled by staining cells with propidium iodide (PI; Sigma) at a concentration of 200 μg/ml for 10 min at room temperature. Incorporation of the dye and cellular integrity were quantified by an EPICS XL flow cytometer (Coulter Electronics; Krefeld, Germany).
Antibodies
The following monoclonal antibodies (MAbs) were used for flow cytometry and pre-embedding immunoelectron microscopy: anti-tryptase [AA1; kindly provided by Dr. Walls, Southampton, UK (Walls et al. 1990)], anti-CD68 (KP1; Dako, Hamburg, Germany), anti-IL-8 [52E8; kindly provided by Dr. Schröder, Kiel, Germany (Sticherling et al. 1989)], and the respective irrelevant isotype control MAbs (Dianova; Hamburg, Germany). For both applications, identical antibody concentrations were used: CD68 20 μg/ml, AA1 10 μg/ml, and 52E8 45 μg/ml.
Immunofluorescence Staining Protocol for Flow Cytometry
For detection of intracellular antigens, HMC-1 cells were microwave-fixed in a mixture of 4% PFA and 0.1% GA as described above. To block nonspecific FcR-mediated binding of MAbs, 5 × 105 cells were preincubated for 30 min at 4C with human heat-inactivated AB serum (Behring Werke; Marburg, Germany) containing 0.025% saponin. Saponin treatment was done during the entire staining procedure as described above. After blocking, primary MAbs were added to a final volume of 50 μl. Cells were incubated for 30 min at 4C, washed twice in PBS containing 0.5% BSA/0.025% saponin (washing buffer), and were resuspended in human AB serum. Binding of the primary MAb was visualized by indirect immunofluorescence using a DTAF-conjugated F(ab')2 fragment of goat anti-mouse IgG antibodies (Dianova). The secondary antibody was added at a final concentration of 20 μg/ml, and cells were further incubated for 30 min at 4C. Finally, cells were washed and fixed in PBS containing freshly prepared 1% PFA. At least 10, 000 cells were analyzed using an EPICS XL flow cytometer (Coulter Electronics). Results were expressed as percent positive cells, taking into account the amount of nonspecific binding of the isotype-matched control antibody (negative control).
Immunoelectron Microscopy (Pre-embedding Procedure)
The protocols for cell fixation, permeabilization, and staining with the primary MAb used for immunoelectron microscopy were identical with those described for flow cytometric detection of intracellular antigens. Binding of the primary antibody was visualized by incubating cells overnight at 4C with goat anti-mouse IgG conjugated with 1-nm colloidal gold (British BioCell International; Cardiff, UK). Negative controls included the substitution of an isotype-matched control MAb for the primary antibody in the labeling scheme and omitting the primary MAb. The secondary antibody was diluted 1:200 in buffer containing 0.5% BSA/0.025% saponin and 0.1% fish gelatin (British BioCell International). After intense washing, cells were fixed for 10 min in 2.5% GA and were then washed two more times with PBS for 10 min each, followed by an incubation in 2% OsO4 for 1 hr at 4C. Finally, cells were washed in PBS and distilled water and immunolabeling was completed by silver stain amplification with a commercially available “Silver Enhancing Kit” (British BioCell International) according to the instructions of the manufacturer. Enhancement was performed at 20C for 30 min and was terminated by washing the specimens in distilled water for 5 min. Before dehydration, cells were centrifuged in molten agar (1% in distilled water at 45C) to form cell pellets that were sliced and processed as small blocks. Dehydration was done in a graded series of ethanol solutions at 4C. The dehydrated samples were transferred to propylene oxide and embedded in Araldite (Serva; Heidelberg, FRG). Polymerization was done for 48 hr at 60C. Semithin sections (0.5 μm) were mounted on glass slides and stained with toluidine blue for quick evaluation. Ultrathin sections (50 nm) were picked up on copper grids and stained with aqueous uranyl acetate and lead citrate. Specimens were examined with a Zeiss EM906 transmission electron microscope at 80 kV.
Results
Permeabilization Protocol
Figure 1 shows the influence of increasing concentrations of saponin (0.003–0.05%) on the permeabilization of unfixed HMC-1 cells measured by the uptake of propidium iodide (F12/PI) and on the cellular integrity determined by measuring forward scatter parameter (FS). As little as 0.006% saponin caused a distinct permeabilization of nearly 50% of cells, but this effect was accompanied by cell destruction, as is evident from the decrease in the forward scatter parameter and from light microscopic evaluation. At a concentration of 0.025% all cells were stained. Triton X-100 produced similar effects, but at higher concentrations: at a threshold concentration of 0.03%, Triton X-100 caused permeabilization of 60% of cells, whereas no effect was detectable at 0.02% (data not shown).
These results indicated that the plasma membrane of mast cells reacts extremely sensitively to the detergents used, and it was therefore concluded that permeabilization requires additional cell fixation.
Fixation Protocol
The effect of various fixatives on permeabilization and cell integrity was investigated for saponin- and Triton X-100-treated as well as for untreated cells (Figure 2). The light scatter properties of the majority of cells (>95%) that had not been treated with a detergent but were microwave-fixed with PLP or 1% PFA indicated sufficient preservation of the cell structure. However, under these conditions no cell permeabilization was achieved because these cells stained negatively for PI. The minor population of small cells that had incorporated PI probably represent damaged and dead cells.
Cells fixed with combinations of PFA and GA showed significant PI uptake, indicating partial cell permeabilization (Figure 2). Because cells fixed in 4% PFA alone showed only minor PI uptake, it is likely that permeabilization was mainly induced by GA. To ensure that GA alone or its combination with microwave irradiation is responsible for the permeabilizing effect observed, cells were fixed without irradiation and the same effect was ascertained (data not shown). This fixation-induced permeabilization allowed diffusion of small molecules such as PI through the plasma and nuclear membrane, but was insufficient for immunoglobulins, as could be shown by an insufficient staining of these cells for intracellular antigens such as CD68 and IL-8 in comparison to cells treated with saponin or Triton X-100 after fixation (data not shown). Destruction of cell integrity by saponin could be prevented by prefixation of cells in solutions containing at least 4% PFA and 0.1% GA (Figure 3). Fixation with PFA alone or PLP did not ensure sufficient stabilization of cell structure in the presence of saponin (Figure 3). This holds also true for Triton X-100-permeabilized cells (data not shown).
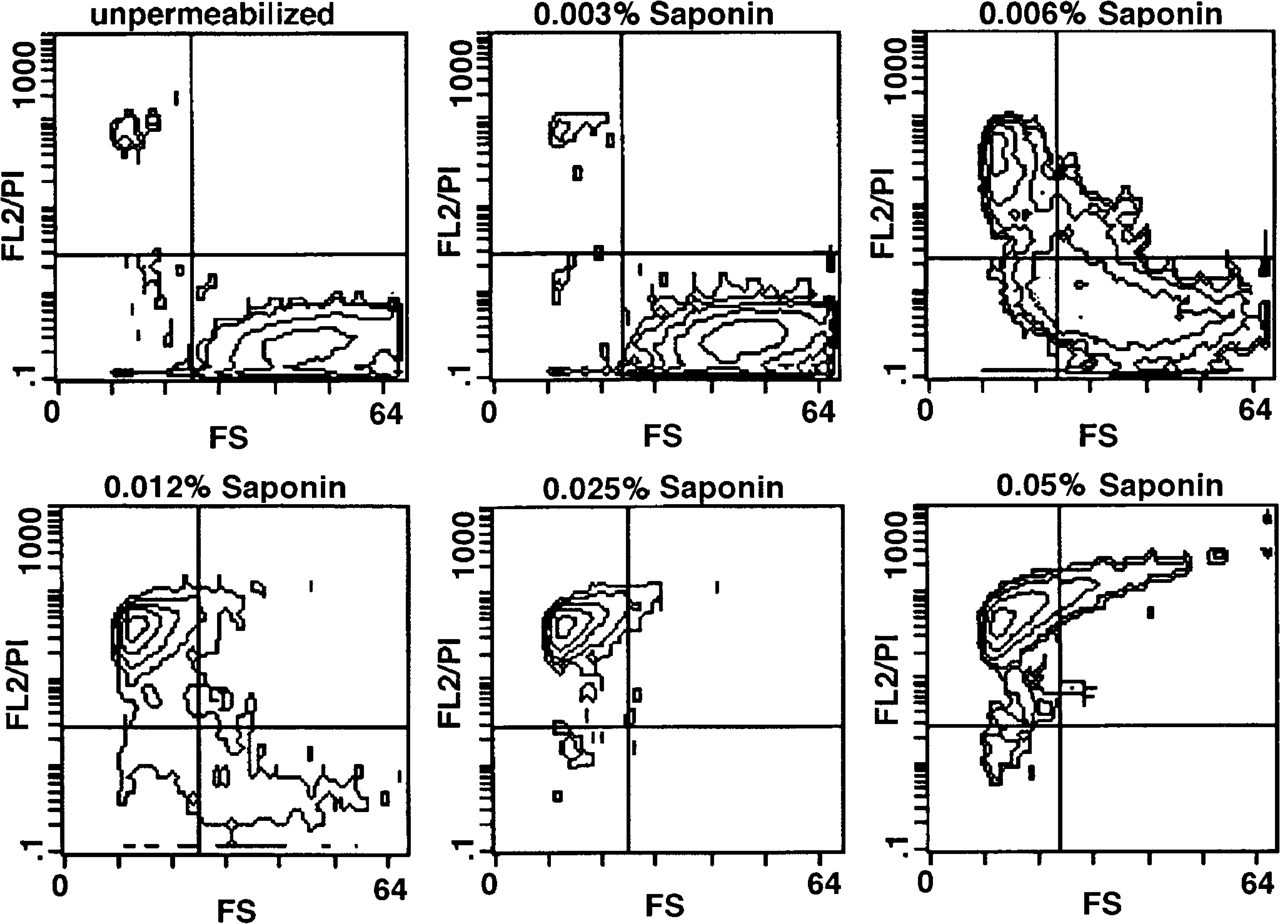
Effect of increasing saponin concentrations (0.003–0.05%) on forward scatter (FS) characteristic and PI uptake (FL2/PI) of HMC-1 cells (contour plots). Unfixed cells were treated for 10 min with increasing concentrations of saponin, as described in Materials and Methods.
Immunolabeling for Flow Cytometry
After determination of optimal conditions for cell fixation (4% PFA/0.1% GA) and permeabilization (0.025% saponin), the binding of MAbs directed against intracellular antigens [tryptase (AA1), CD68 (KP1), and IL-8 (52E8)] was quantified by flow cytometry. CD68 and tryptase, two antigens expressed intracellularly in human mast cells (Harvima and Schwartz 1993; Nilson et al. 1994), were used as positive controls. As shown in Figure 4A, both antigens were not detectable on the surface of unfixed and unpermeabilized HMC-1 cells. After fixation and permeabilization, tryptase, and to a much greater extent CD68, could be detected by the corresponding MAbs (Figure 4B). By comparing the curves for the isotype-matched control MAb of untreated and permeabilized cells (negative controls), it was apparent that permeabilization caused a significant increase in nonspecific staining.
The histograms in Figure 5 demonstrate the expression of IL-8 in unstimulated and stimulated HMC-1 cells. After cell fixation and permeabilization, the cytokine was measurable in small amounts in unstimulated and calcium ionophore-treated cells (Figures 5A and 5B). PMA treatment and, to a much greater extent, the combination of calcium ionophore and PMA induced a significant increase in the expression of cytoplasmic IL-8 (Figures 5C and 5D). The plasma membrane of calcium ionophore/PMA-stimulated and unstimulated cells was negative for IL-8, as determined by immunostaining in the absence of cell fixation and permeabilization (data not shown). The time course of the accumulation of intracellular IL-8 in calcium ionophore/PMA-stimulated cells is shown in Figure 6. Even after 2 hr of stimulation there was a measurable increase in IL-8 expression, which reached its maximum at 12 hr. Thereafter, the cytoplasmic IL-8 content declined.
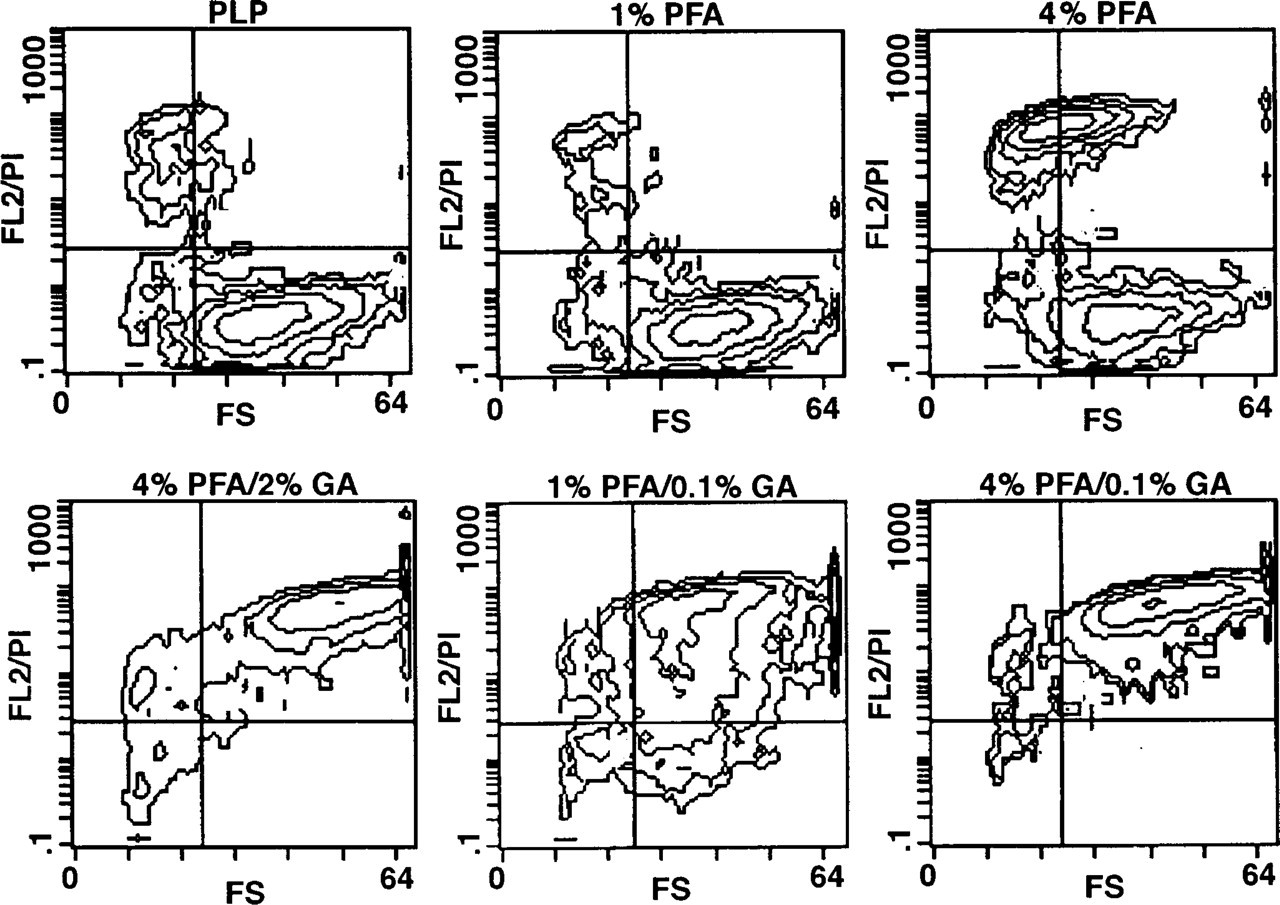
Effects of various fixatives on forward scatter (FS) characteristic and PI staining (FL2/PI) of HMC-1 cells in the absence of detergents (contour plots). Fixation was done in a microwave oven, as described in Materials and Methods.
Stimulation times longer than 12 hr induced a decrease in cell viability of 10–20%, but dead cells and debris were electronically gated out on the basis of shifted forward and side-angle light scatter parameters.
Immunolabeling for Electron Microscopy
The fixation and permeabilization protocol, as established by flow cytometry, was applied for pre-embedding immunoelectron microscopic detection of IL-8 in HMC-1 cells. Under these conditions good ultra-structural preservation was noticeable, as verified by the depiction of mitochondrial membranes and secretory granules (Figure 7). HMC-1 cells exhibited the typical appearance of immature human mast cells, with large nucleoli in a monolobed round or ovoid nucleus, some immature granules, and many mitochondria. Granules of the cells exhibited an amorphous structure without the crystalline gratings frequently seen in the granules of mature tissue mast cells (Figures 7B–7D). Furthermore, the majority of HMC-1 cells cultured under suspension conditions are characterized by the appearance of empty granule containers and degranulation channels, indicating spontaneous piecemeal degranulation (Figure 7A). Most importantly, the antigenicity of intracellular IL-8 was retained. In HMC-1 cells stimulated for 8 hr with 0.25 μM A23187 and 25 ng/ml PMA, silver-enhanced 1-nm colloidal gold complexes were observed as large, specific electron-dense particles that were mainly located at the periphery of cytoplasmic granules (Figures 7B and 7C). Only a few granules showed a more homogeneous or even negative staining for IL-8 (Figures 7B and 7C). Empty granule containers and degranulation channels exhibited only minimal or no immunostaining for IL-8 (Figure 7C). In granules of unstimulated cells, no immunoreactivity was detectable (Figure 7A). The nonspecific background labeling was minimal, and no immunoreactive structures were seen in cells processed in the presence of an isotype-matched irrelevant MAb (Figure 7D, negative control) or after omitting the primary MAb during immunostaining (data not shown).
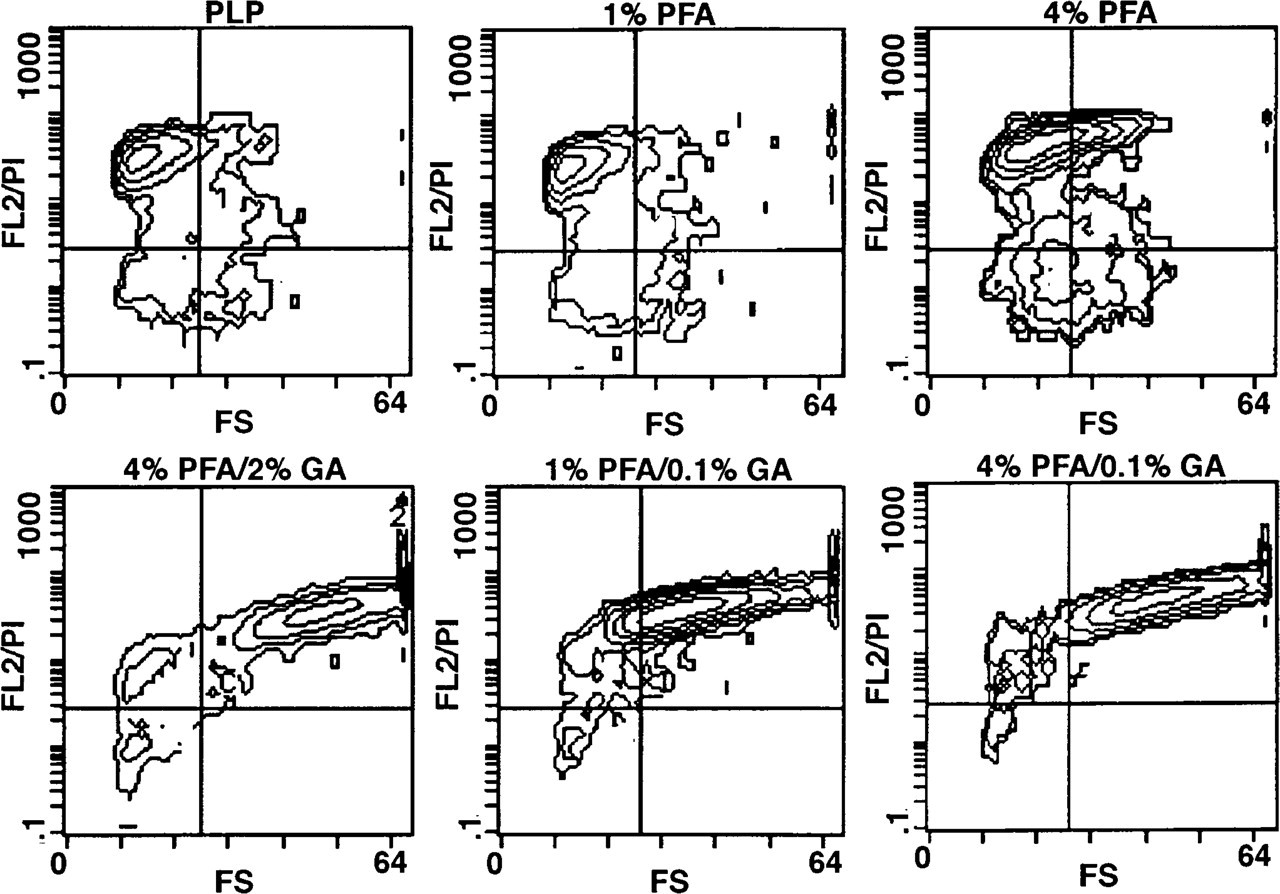
Effect of saponin (0.025%) on forward scatter (FS) characteristic and PI staining (FL2/PI) of prefixed HMC-1 cells (contour plots). After microwave-fixation, cells were treated with saponin, as described in Materials and Methods.
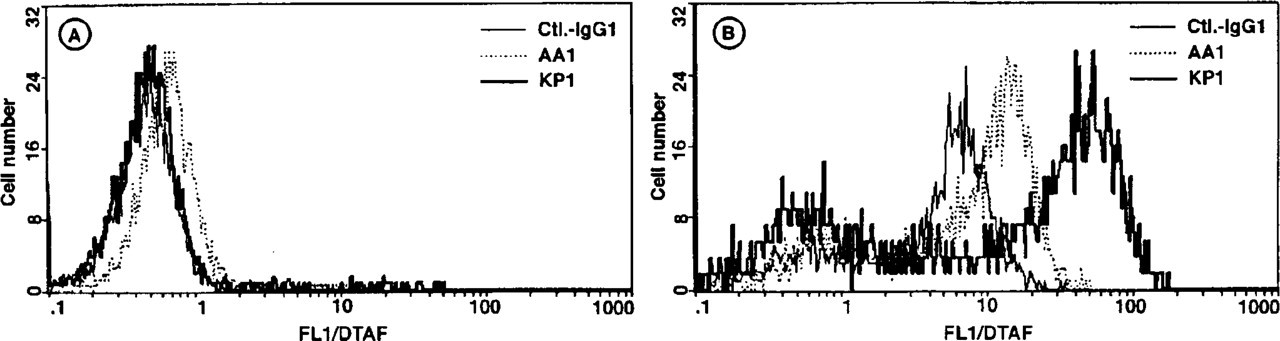
Flow cytometric quantification of intracellular tryptase (AA1) and CD68 (KP1) in unstimulated HMC-1 cells (histograms). These mast cell-associated intracellular antigens were used as positive controls. None of these antigens was expressed on the cell surface, as determined by staining of unpermeabilized and unfixed cells (
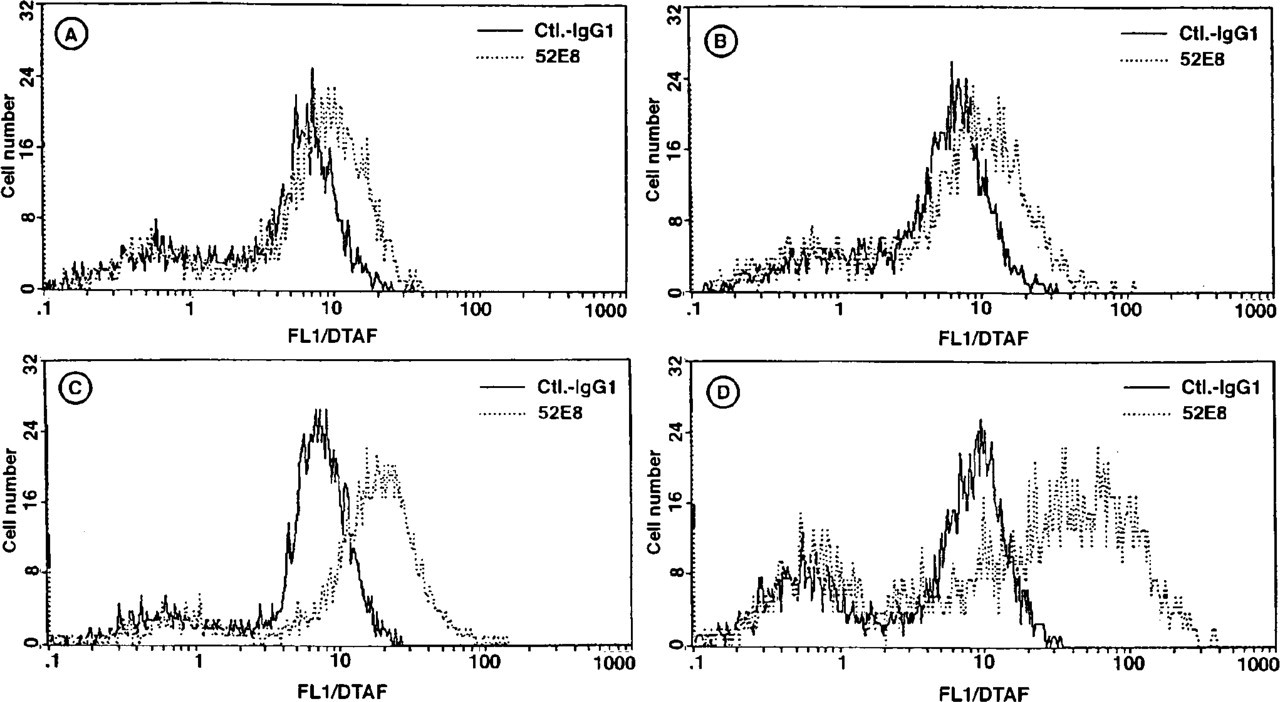
Flow cytometric quantification of intracellular IL-8 (52E8) in unstimulated HMC-1 cells (
Discussion
We previously identified IL-8 expression in human mast cells under inflammatory conditions (Möller et al. 1993). In the present study we complete these findings and demonstrate IL-8 expression at the single-cell level in HMC-1 cells with the help of flow cytometry and pre-embedding immunoelectron microscopy. For this purpose we developed a mast cell-specific fixation and permeabilization protocol that allowed intracellular semiquantitative measurement of IL-8 and other intracellular antigens by FACS so that we could detect intracellular accumulation of IL-8 on cell stimulation with the PKC activator PMA and the calcium ionophore A23187. This result is in accordance with our previous results (Möller et al. 1993) demonstrating secretion of IL-8 in HMC-1 culture supernatants by a cytokine-specific ELISA. However, when we compared the kinetics of intracellular accumulation of IL-8 with the kinetics of IL-8 release, remarkable differences were observed. Flow cytometric detection revealed a more rapid increase in the number of IL-8 producing cells already measurable within 2 hr and 4 hr after stimulation, which plateaued between 8 hr and 12 hr. In the supernatant of stimulated HMC-1 cells, IL-8 was detectable between 4 hr and 8 hr and release continued for up to 48 hr (Möller et al. 1993). This difference indicates that intracellular measurement of IL-8 in mast cells represents the more sensitive detection method. Mast cells probably store IL-8 immediately after its synthesis in their specific granules, as could be observed by immunoelectron microscopy, and an accumulation effect is achieved. The capacity of HMC-1 cells for intracellular storage of IL-8 would also provide a plausible explanation for the fact that it was unnecessary to use agents such as monensin or brefeldin A, which have been used by others to concentrate cytokines intracellularly in lymphocytes, allowing their detection by flow cytometric analysis only (Jung et al. 1993; Elson et al. 1995). These agents disrupt the intracellular transport of secretory proteins, which in consequence were concentrated around the Golgi area (Tartakoff et al. 1978; Misumi et al. 1986). The results obtained by immunoelectron microscopic investigation revealed the subcellular localization of IL-8 exclusively in electron-dense granules of stimulated HMC-1 cells. This finding is in accordance with our previous results demonstrating the expression of granule-associated IL-8 in IgE-stimulated human skin mast cells (Möller et al. 1993), and suggests that not only mature mast cells possess the capability for storage of inflammatory cytokines after appropriate stimulation but also immature cells. Because not all granules of HMC-1 cells exhibited positive immunostaining, it appears reasonable to assume that qualitatively different granules already exist in immature human mast cells, as has been demonstrated for normal mature mast cells (Dvorak 1992).
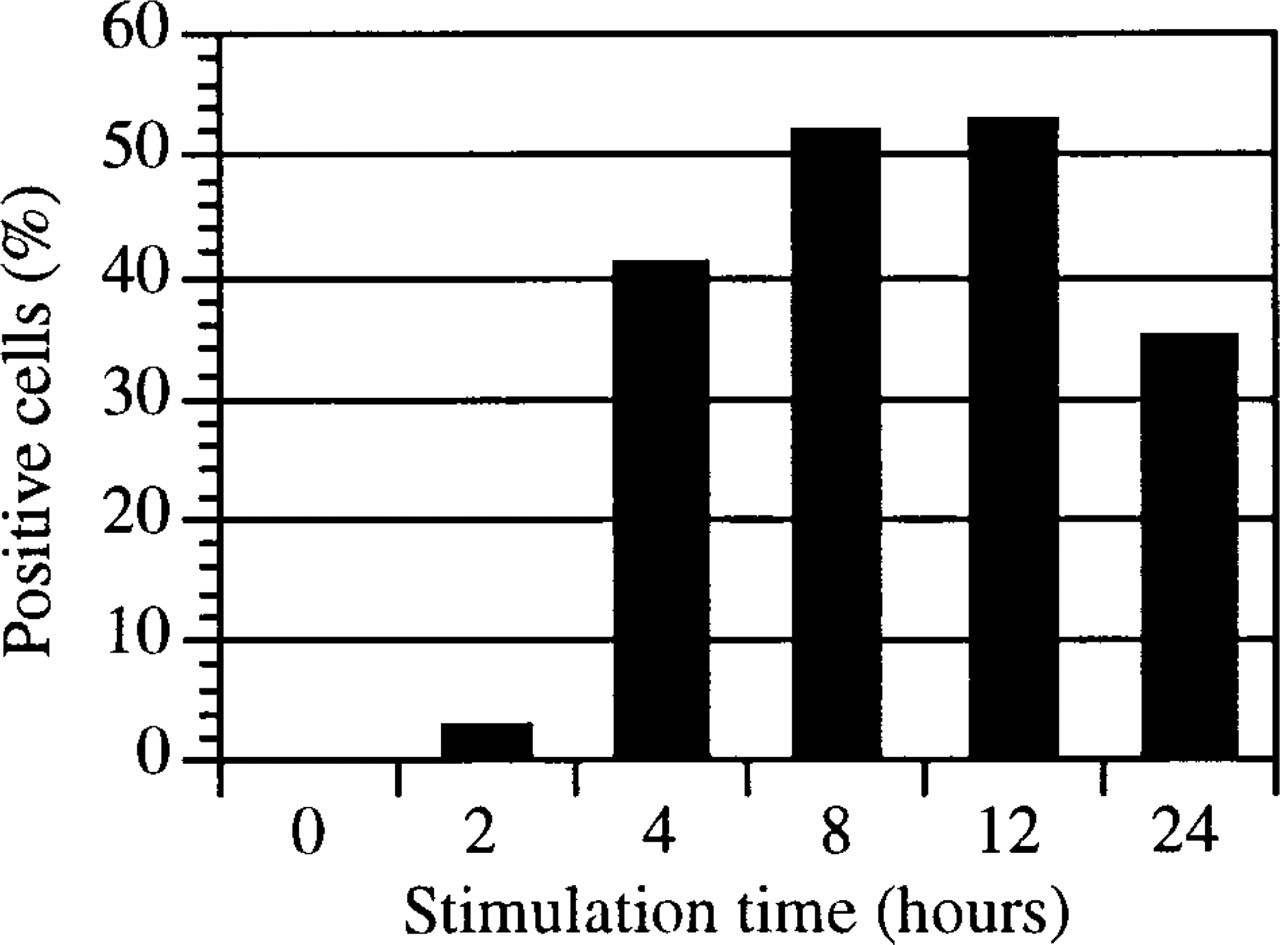
Flow cytometric quantification of intracellular interleukin-8 (52E8) in HMC-1 cells stimulated for indicated time intervals with 0.25 μM A23187 and 25 ng/ml PMA. After stimulation, cells were immediately microwave-fixed with 4% PFA/0.1% GA and permeabilized with 0.025% saponin. The Y-axis represents the percentage of cells positively stained for IL-8. Similar results were obtained in three independent experiments.
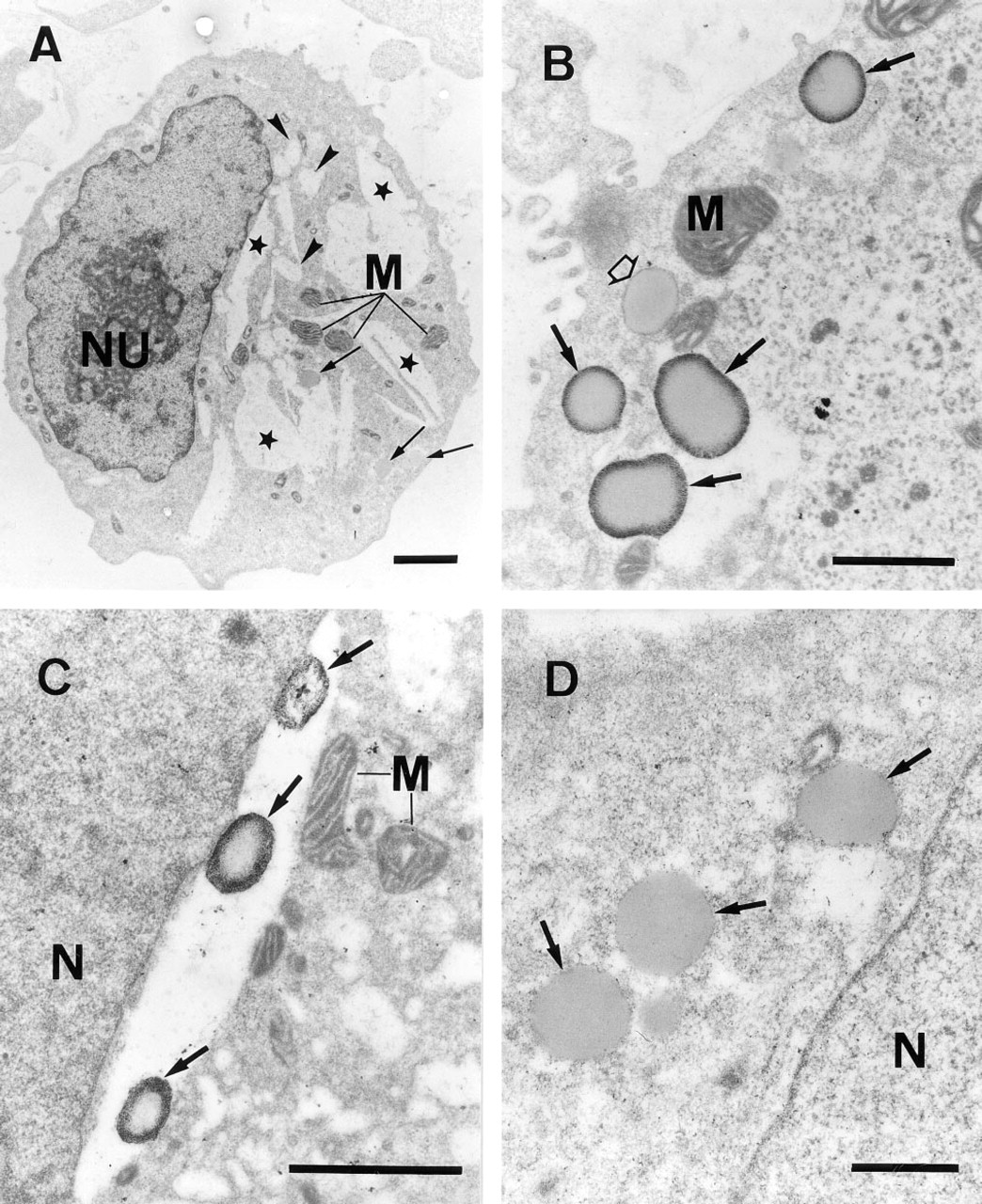
Transmission electron micrographs of an unstimulated HMC-1 cell (
The successful identification of IL-8 by the pre-embedding immunogold technique described suggests that flow cytometry is a useful tool for rapid screening of multiple parameters necessary for optimal cell fixation, permeabilization, and antibody binding. We used PI as a reliable probe for the determination of the extent of cellular permeabilization, although it must be remembered that PI possesses a substantial lower molecular size than immunoglobulins. For cell fixation, we applied the microwave energy technology because this method has been described to ensure an excellent preservation of the ultrastructure of mast cells and of various mast cell-associated antigens (Login et al. 1987; Beil et al. 1994; Jamur et al. 1995). In this way, we were able to show that permeabilization of HMC-1 cells required prefixation with 4% PFA and at least 0.1% GA. Fixatives without GA, such as PFA alone or PLP, did not ensure cellular integrity after detergent-induced permeabilization, as shown by measurement of the forward scatter parameter during FACS analysis. These conditions are mast cell-specific and differ from those applied for monocytes and lymphocytes, which have been reported to be sufficiently fixed by PFA alone before permeabilization (Halldén et al. 1989; Hofsli et al. 1989; Sander et al. 1991; de Caestecker et al. 1992).
Fixation with 4% PFA and 2% GA according to Karnovsky (1965) ensured excellent preservation of ultrastructural morphology, but the high content of GA frequently destroys the antigenicity of many proteins (Leenen et al. 1985). In addition to cell fixation, GA alone induced partial permeabilization of HMC-1 cells even at a minimal concentration of 0.1%. This membrane-permeabilizing effect of GA was also described by Dykstra (1992). GA-induced permeabilization, however, was insufficient for successful antibody penetration of the plasma membrane. In accordance with van Dort et al. (1988), we were unable to detect microwave irradiation-induced membrane permeabilization, because irradiated cells fixed with 1% PFA or PLP showed no PI uptake.
Treatment with Triton X-100 and saponin induced sufficient membrane permeabilization at very low but distinct threshold concentrations (0.025% and 0.006%, respectively). This extreme sensitivity of mast cells in response to saponin indicates a high cholesterol content in the plasma membrane because it has been reported that the plant glycoside intercalates into eukaryotic cell membranes by reversibly interacting with this steroid (Bangham and Horne 1962). In addition to this characteristic of saponin, it also does not induce autofluorescence or cell aggregation and it does not alter antigenicity of most membrane antigens investigated (Jacob et al. 1991). Therefore, we chose this detergent at a concentration of 0.025% for partial permeabilization of mast cell membranes prefixed with 4% PFA and 0.1% GA. The usefulness of this fixation protocol for detecting intracellular antigens in mast cells could be demonstrated by the flow cytometric measurement of CD68 and tryptase, two antigens expressed exclusively in the cytoplasm of HMC-1 and other mast cells (Horny et al. 1990; Harvima and Schwartz 1993; Nilson et al. 1994). Surface staining for these antigens, as determined by analysis of unfixed and unpermeabilized cells, was not detectable. Successful identification of intracellular CD68 by flow cytometry has already been described for peripheral leukocytes and bone marrow cells (Knapp et al. 1994; Strobl et al. 1995).
Taken together, the present findings demonstrate semiquantitative IL-8 expression at the single-cell level and its ultrastructural localization in stimulated HMC-1 cells. Furthermore, this study provides a strategic protocol established by flow cytometry that allows rapid optimization of fixation and permeabilization conditions for detection of intracellular antigens in mast cells by pre-embedding immunoelectron microscopy.
Footnotes
Acknowledgements
Supported in part by “Forschungsprojektschwerpunkt” of the Free University of Berlin. We are grateful to Prof Schnoy from the Department of Pathology, Virchow Clinics, Berlin, for making available the facilities of transmission electron microscopy, to Ms Lajous–Petter for helpful suggestions and advice, and to Ms Pröhl for photographic assistance.