
Abstract
We report the production of a monoclonal antibody (d1C4) by in vitro immunization that has immunoreactivity with a native chondroitin sulfate epitope in embryonic chick limb and heart. Murine lymphocytes were stimulated by direct exposure to unfixed, unsolubilized precartilage mesenchymal aggregates in high-density micromass culture derived from Stage 22–23 chick limb buds. Specificity of d1C4 reactivity was demonstrated by sensitivity of immunohistochemical staining to pretreatment with chondroitinase ABC or AC, preferential immunoreactivity with chondroitin-6-sulfate glycosaminoglycan (CS-C GAG) in ELISA, and competition of immunohistochemical staining with CS-C GAG. Immunohistochemical analysis of the expression of the d1C4 epitope revealed a striking localization of immunoreactivity in the extracellular matrix (ECM) of precartilage aggregates of chick limb mesenchyme in high-density micromass culture by 16 hr and the prechondrogenic limb core at Stage 23 in vivo. Immunoreactivity in both cultured limb mesenchyme and the embryonic limb continued through differentiation of prechondrogenic condensations into cartilage tissue. In the developing chick heart, d1C4 staining was found throughout the ECM of atrioventricular cushion tissue by Stage 25, but was localized to mesenchyme adjacent to the myocardium in the outflow tract cushions. There was an abrupt demarcation between d1C4-reactive intracardiac mesenchyme and unreactive extracardiac mesenchyme of the dorsal mesocardium in the Stage 22 embryo. This study demonstrates the efficacy of in vitro immunization of lymphocytes for the production of MAbs to native ECM constituents, such as CS-GAGs. Immunohistochemical data utilizing d1C4 suggest that CS-GAGs bearing this epitope may be important in early morphogenetic events leading to cartilage differentiation in the limb and valvuloseptal morphogenesis in the heart.
Keywords
P
To study the intricate processes involved in these critical morphological events, it is often advantageous to utilize immunological probes to native ECM components. Such reagents are useful in the evaluation of ECM interactions by avoiding artifactual alterations in the organization of the matrix caused by the enzymatic creation or unmasking of epitopes for immunohistochemical analyses. One problem with the identification of ECM components such as GAGs, however, has been the difficulty in producing antibodies to the native structure. This difficulty is due in large part to the poor immunogenicity of many ECM molecules, particularly carbohydrate sequences of GAG chains (Avnur and Geiger 1984; Rouslahti 1988; Sorrell et al. 1990). The use of in vitro immunization of mouse lymphocytes (Luben and Mohler 1980) offers a potential solution to this problem because the tolerance for conserved immunogens in vivo may be overridden by lymphocytes in culture (Reading 1982).
In this article we report the preparation of a monoclonal antibody (d1C4) by in vitro stimulation of isolated lymphocytes with cultured limb mesenchyme. Characterization of MAb d1C4 demonstrates that it preferentially recognizes a native epitope on GAG chains enriched in chondroitin-6-sulfate. Immunofluorescence microscopy localizes the d1C4 epitope in prechondrogenic aggregates of chick limb mesenchyme in vitro and in the prechondrogenic limb core and cardiac cushion tissue in vivo.
Materials and Methods
Cell Cultures
High-density micromass cultures were prepared using Stage 22–23 (Hamburger and Hamilton 1951) chick limb buds with a slight modification of the procedure described by Ahrens et al. (1977). Briefly, limb buds were removed, washed in PBS, and incubated for 15 min at 37C in 0.1% trypsin-PBS. The enzymatic digestion was stopped by the addition of complete medium, i.e., Ham's F-12 (Gibco; Gaithersburg, MD) containing 10% fetal bovine serum (FBS; Hyclone Laboratories, Logan, UT) and 100 U/ml penicillin-100 μg/ml streptomycin (Gibco). The cell suspension was centrifuged and the pellet was suspended in complete medium and filtered through two layers of #10 Nitex. Cell number was adjusted to 2 × 107 cells/ml and 10-μl drops were plated in 35-mm tissue culture dishes (Falcon; Becton Dickinson Labware, Lincoln Park, NJ). Cultures were incubated for 1 hr at 37C in a humidified atmosphere of 95% air/5% CO2 and then flooded with 2 ml complete medium. Cultures were maintained up to 3 days with complete changes of medium daily. In selected experiments, micromass cultures were grown for 3 days in complete medium containing 2 mM p-nitrophenyl-β-
Production of Monoclonal Antibodies In Vitro
Four 0.1-ml micromass cultures of chick limb mesenchyme (prepared as described above) were established in separate wells of a 24-well tissue culture plate (Corning; Ithaca, NY). After 24 hr of incubation, cultures were washed three times with sterile PBS and air-dried under laminar flow following the method of Gilbert et al. (1990) for the preparation of renal collecting duct epithelia for in vitro immunization.
Thymocyte conditioned medium was prepared by a modification of the method of van Ness et al. (1984). Thymus glands from four 6-week female Balb/C mice were dissected aseptically, teased apart with 21-gauge needles, and washed with serum-free RPMI (SFRPMI; Gibco). Thymocytes were suspended in RPMI 1640 containing 40% heat-inactivated fetal calf serum (HIFCS; Gibco) and 4 mM glutamine, 2 mM sodium pyruvate, 100 μM β-mercaptoethanol, and antibiotics (representing a 2 x supplemented medium stock) for subsequent in vitro immunization. Thymocytes were cultured in T25 flasks (Corning) for 3 days. Cells were removed by centrifugation and thymocyte conditioned medium was stored at −20C until use.
Macrophages were obtained from a nonimmunized 7-week female Balb/C mouse by peritoneal lavage with Hank's balanced saline solution (Campbell 1984). The same mouse was used to harvest splenocytes using standard procedures (Harlow and Lane 1988).
Each of four air-dried prechondrogenic micromass cultures of chick limb mesenchyme received 1 × 107 splenocytes and 1.8 × 104 peritoneal macrophages in 1 ml SFRPMI containing 30 μg/ml N-acetylmuramyl-
Stimulated splenocytes were harvested by centrifugation and fused with the myeloma cell line SP2/0 at a ratio of 3:1 with 50% polyethylene glycol (
Antibody Purification
Antibodies were purified from tissue culture supernatants using anti-mouse IgM–agarose (Sigma). Bound antibodiesc were eluted with 0.1 M glycine, pH 2.6, containing 150 mM NaCl. Fractions were neutralized immediately with 1 M Tris, pH 8, and the aliquots were dot-blotted onto nitrocellulose (Bio-Rad Laboratories; Hercules, CA) and stained with 0.1% amido black (Sigma) to detect protein. Fractions containing eluted antibodies were concentrated and exchanged into PBS using Centricon-10 ultrafiltration devices (Amicon; Beverly, MA). Protein content was determined using the Micro BCA protein assay kit (Pierce; Rockford, IL).
Immunohistochemistry
Micromass cultures of chick limb mesenchyme were fixed in 80% methanol–20% DMSO (Dent's fixative) at room temperature (RT) and processed for indirect immunofluorescence microscopy in the tissue culture dishes. Cultures were rehydrated in PBS and blocked for 30 min at RT with PBS containing 2% bovine serum albumin (BSA; Sigma) and 1% normal goat serum (NGS; Cappel Research Products, Durham, NC). Fixed specimens were incubated for 1 hr with 10 μg/ml d1C4 or undiluted tissue culture supernatants, washed three times with PBS, then incubated for 1 hr with a 1:100 dilution of fluorescein-conjugated goat anti-mouse IgM or IgG (Cappel) in BSA–PBS. For double labeling with rhodamine-conjugated peanut agglutinin (RhPNA; Vector Laboratories, Burlingame, CA), selected cultures that had been reacted with antibodies as described above were washed with PBS and incubated with 0.1 mg/ml RhPNA in PBS for 30 min. All specimens were washed four times with PBS and mounted in 10% PBS–90% glycerol containing 25 mg/ml 1,4-diazabicyclo(2,2,2)octane (DABCO; Sigma) (Johnson et al. 1982). In selected experiments, micromass cultures were fixed in ethanol containing 3% glacial acetic acid for 10 min, washed with 0.1 N HCl, and stained with 1% Alcian blue-0.1 N HCl (Lev and Spicer 1964) for 1 hr to reveal chondrogenic foci. Cell layers were destained with 0.1 N HCl, washed thoroughly with PBS, and processed for indirect immunofluorescence microscopy.
Chick embryos were fixed in 80% methanol–20% DMSO at −20C, embedded in paraffin, and sectioned at 5 μm. Alternatively, embryonic chick thoracic regions with attached hearts were cryopreserved by freeze-substitution into ethanol (Kitten et al. 1987), followed by embedding in paraffin. Deparaffinized sections were rehydrated to PBS, blocked with BSA–PBS containing 1% NGS, and processed as above with primary antibody incubations overnight at 4C in a humidified chamber and fluorescein-conjugated secondary antibody incubations for 2 hr at RT. Sections were washed four times with PBS after each antibody incubation and mounted in DABCO medium.
Controls for immunohistochemistry included omission of the primary antibody or use of an irrelevant IgM. Specificity of d1C4 immunoreactivity was also tested by inclusion of a 100-fold (w/w) excess of chondroitin sulfate Type A (CS-A, sturgeon notochord; Seikagaku America, Rockville, MD) or chondroitin sulfate type C GAGs (CSC, shark cartilage; Sigma) with the primary antibody incubation. II6B3, an MAb to Type II collagen, was obtained through the Developmental Studies Hybridoma Bank maintained by the Department of Biological Sciences, University of Iowa. Specimens were examined with a Zeiss Axioskop equipped with epifluorescence optics and were photographed using Kodak TMAX 400 film.
Characterization of the d1C4 Epitope
Fixed micromass cultures were treated for 2 hr at 37C with either 0.1 U/ml chondroitinase ABC (Sigma) in 0.1 M Tris, pH 8, or 0.2 U/ml chondroitinase AC (Sigma) in PBS, pH 7.2, before immunostaining. For hyaluronidase digestion, micromass cultures and deparaffinized tissue sections were incubated for 30 min at RT in 0.1% testicular hyaluronidase (Sigma) in 50 mM sodium acetate, pH 6, with 0.9% NaCl, 1 mM phenylmethylsulfonylfluoride (PMSF), 1 μM leupeptin, 1 μM pepstatin A, and 1 mM EDTA before blocking and immunofluorescence staining. Periodate oxidation of fixed micromass cultures was performed using 10 mM periodic acid in 50 mM sodium acetate, pH 4.5, for 1 hr in the dark as described previously (Woodward et al. 1985). Cultures were washed with PBS and incubated in 50 mM sodium borohydride–PBS for 30 min. Control cultures were incubated in 50 mM sodium acetate alone followed by reduction with 50 mM sodium borohydride. Cell layers were washed thoroughly with PBS before the blocking and immunostaining steps.
Microtiter plates (Immulon 4; Dynatech Laboratories, Chantilly, VA) were treated with 1 μg/ml poly-
Results
Mouse lymphocytes were immunized in vitro with unfixed, air-dried micromass cultures of chick limb mesenchyme to produce MAbs reactive with native ECM molecules expressed during the critical process of cellular aggregation preceding overt limb chondrogenesis. At the time of screening, macroscopic hybridoma colonies were found in 83% of wells (20/24 wells), with 40% of these (8/20) yielding MAbs reactive by indirect immunofluorescence microscopy on frozen chick limb sections. Several antibodies recognized ECM components in both embryonic limb buds and high-density cultures of limb mesenchyme, one of which, d1C4, is the subject of this report.
The MAb d1C4 exhibited particularly strong immunoreactivity with mesenchymal condensations in 24-hr micromass cultures (Figure 1A). Digestion of micromass cultures or tissue sections by testicular hyaluronidase pretreatment in the presence of protease inhibitors before antibody staining abolished reactivity with d1C4 (data not shown). This result suggested that the antigen was either hyaluronic acid or a chondroitin sulfate GAG, because both are digested by testicular hyaluronidase. Pretreatment of 24-hr micromass cultures with either chondroitinase ABC or chondroitinase AC also revealed sensitivity of the d1C4 epitope (Figure 1B), suggesting that the antigen was a chondroitin sulfate. Periodate treatment of micromass cultures of limb mesenchyme to oxidize non-reducing termini of the GAG chains did not inhibit or diminish immunoreactivity (Figure 1C), which is consistent with an internal carbohydrate epitope.
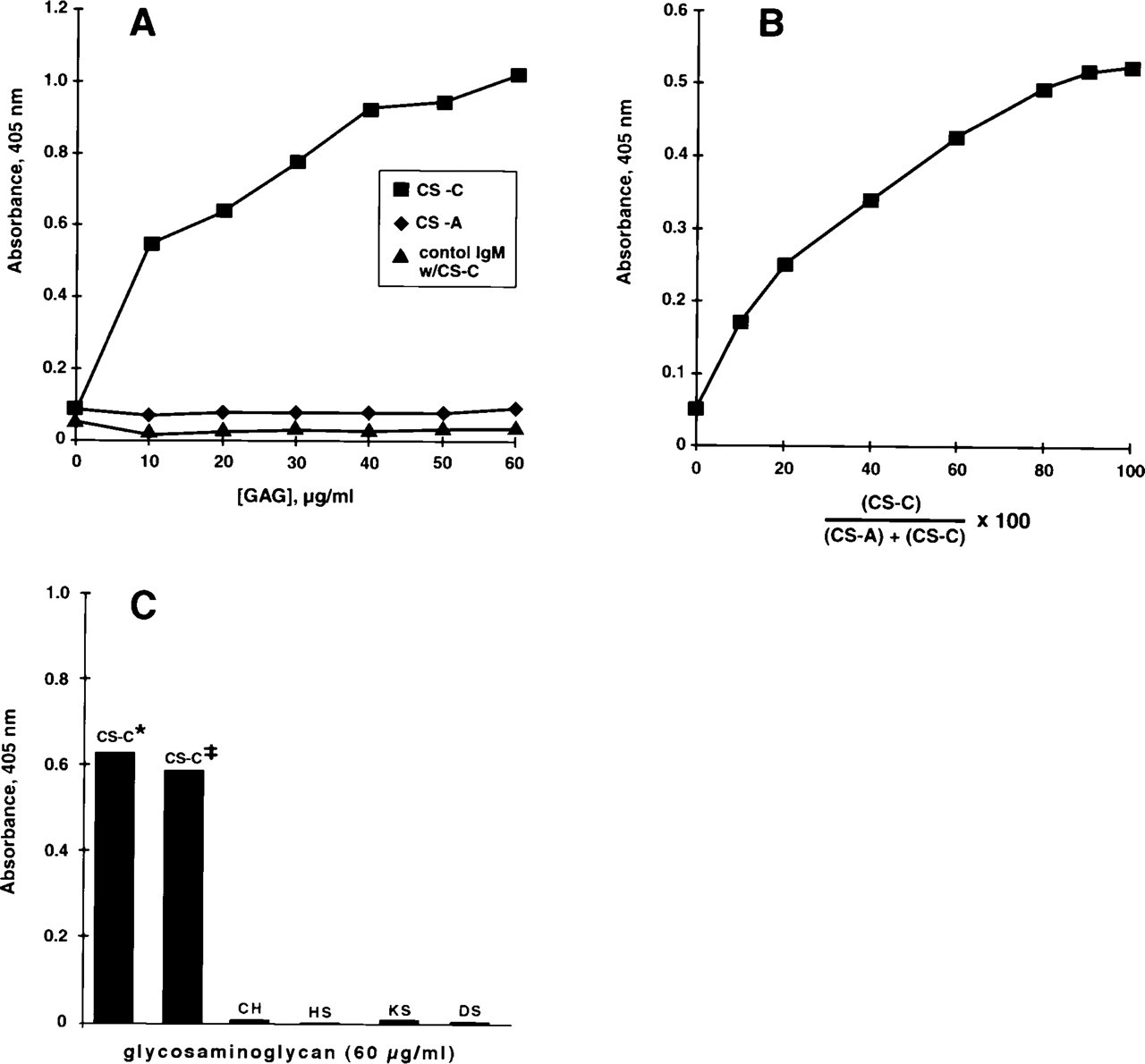
To assess the type of chondroitin sulfate recognized by d1C4, reactivity with chondroitin-4-sulfate (CS-A; sturgeon notochord) and chondroitin-6-sulfate (CS-C; shark cartilage) GAGs was tested by ELISA. The results indicated a preferential reactivity of d1C4 in a concentration-dependent manner with this CS-C GAG preparation and little or no reactivity with the CS-A GAG (Figure 2A). Little or no concentration-dependent reactivity of d1C4 was observed with CS-A GAGs from another tissue source as well (bovine trachea, Sigma and Calbiochem; data not shown). The preferential reactivity of d1C4 with CS-C GAG was corroborated with various ratios of CS-A:CS-C at a constant total concentration of GAG (Figure 2B). The presence of the d1C4 epitope in other GAGs was also tested by ELISA. Little or no reactivity of d1C4 was seen with chondroitin, heparan sulfate, keratan sulfate, or dermatan sulfate (Figure 2C). In addition, immunoreactivity of d1C4 with micromass cultures could be competed by preincubation of the antibody with excess CS-C GAG (Figure 3), whereas preincubation of antibody with the same concentration of CS-A GAG was ineffective. Taken together, these data suggest that the d1C4 epitope is preferentially found in GAGs enriched in CS-C type GAGs.
High-density micromass culture of prechondrogenic chick limb mesenchyme duplicates processes that occur during chondrogenesis in vivo (Ahrens et al. 1977) and is a well-accepted culture bioassay for embryonic cartilage formation. As shown in Figure 4A, precartilage aggregates could be detected in limb micromass cultures with RhPNA lectin by 16 hr, as described previously (Aulthouse and Solursh 1987). However, Type II collagen was immunologically undetectable (Figure 4A, inset). Double labeling of cultures with RhPNA (Figure 4A) and d1C4 (Figure 4B) demonstrated a striking colocalization of CS-GAG epitopes with lectin binding molecules in regions of aggregated mesenchyme before overt chondrogenesis. Interestingly, faint d1C4 immunoreactivity could be observed by 8–12 hr of culture in undifferentiated mesenchyme of the micromass cell layers (data not shown). Localization of PNA-reactive molecules (Figure 4D) and the CS-GAG epitope recognized by d1C4 (Figure 4E) became more restricted to the incipient nodular structures by 48 hr. Immunoreactivity with d1C4 persisted in association with cell aggregates as they matured into cartilage nodules possessing an acidic Alcian blue-positive, chondroitin sulfate proteoglycan-rich ECM by 72 hr of culture (Figures 4G and 4H). Micromass cultures treated with β-
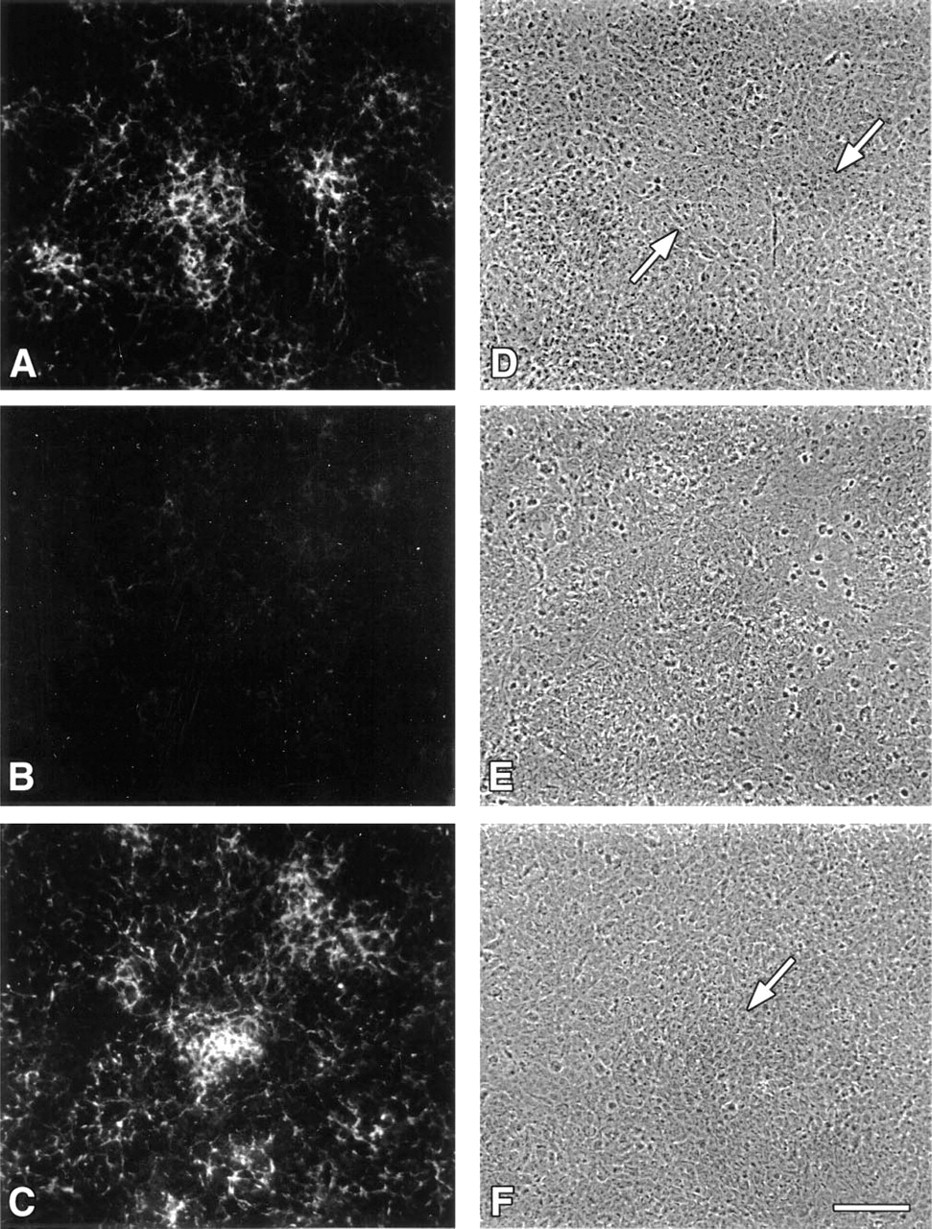
Immunolocalization of d1C4 epitopes in paraffin sections of developing chick limb buds paralleled the micromass data and revealed other interesting patterns of reactivity (Figure 5). At Stage 20, a low level of pericellular immunoreactivity with d1C4 was distributed widely in limb mesenchyme. Limb ectoderm, the ectodermal basement membrane, and the immediately subjacent mesenchyme were strongly immunoreactive (Figure 5A). As shown at higher magnification in Figure 5B, there was an abrupt loss of reactivity associated with the apical ectodermal ridge (AER) and subridge basement membrane of the Stage 20 limb bud. In contrast, PNA binding molecules were localized throughout the ridge, nonridge ectoderm, and associated basement membrane (Figure 5C). The staining pattern found in the body of the Stage 20 embryo showed d1C4 immunoreactivity in and around the neural tube and notochord, and also associated with basement membranes of the dorsal ectoderm, gut, and mesonephros (Figure 5A).
d1C4 immunoreactivity of mesenchymal condensations in micromass cultures of Stage 22–23 limb mesenchyme is sensitive to chondroitinase AC. Indirect immunofluorescence staining with d1C4
d1C4 reacts preferentially with glycosaminoglycans (GAGs) enriched in chondroitin-6-sulfate by ELISA.
By Stage 22, d1C4 immunoreactivity was prominent in the ECM of the proximal limb, associated primarily with mesenchyme extending from the proximal limb core to the ventral sulcus (Figure 5D). At Stage 23, increased immunopositive material localized to the prechondrogenic core of the developing limb (Figure 5E). It was also noted at these latter stages that the nonridge limb ectodermal basement membrane remained immunoreactive, whereas staining was absent in the basement membrane underlying the AER and the mesenchyme of the distal limb. At Stage 23, immunoreactivity extended into the mesenchyme surrounding the gut epithelium. By Stage 28, d1C4 reactivity was associated predominantly with nascent cartilage tissues (Figure 5F). Immunostaining was also seen surrounding larger blood vessels of the limb.
In addition to the limb, interesting patterns of d1C4 immunoreactivity were found elsewhere in extracellular matrices of the developing chick, including the posterior portion of the somites, surrounding the dorsal aorta, in the eye, and in lung mesenchyme (data not shown). A particularly striking distribution of d1C4 reactivity was found in the developing heart (Figure 6). As shown in Figure 6A, the d1C4 epitope was expressed primarily by mesenchyme within the atrioventricular canal (AV) in the Stage 19 heart. At higher magnification it was apparent that those mesenchyme closest to the AV myocardium were only weakly positive relative to regions nearer the endothelium (Figures 6B and 6C). The AV endothelium was unreactive, as were the premigratory cardiac jelly and the AV myocardium (Figures 6B and 6C). Staining was also observed along the subendothelial myocardium in the atrium and at scattered foci throughout the myocardium of the atrial/AV junction (Figure 6A). By Stage 25, the entire inferior AV cushion ECM and all but the distal aspect of the superior AV cushion stained with d1C4 (Figures 7B and 7C). The acellular matrix of the atrium adjacent to the AV cushions was also positive. In contrast, mesenchyme in the conal ridges of the outflow region differentially expressed the d1C4 epitope. Only those mesenchyme in the outflow tract near the myocardium were stained. Interestingly, the dense layer of myocardium (asterisks in Figures 7B and 7C) in the outflow tract and AV region, as well as the epicardium (arrowheads), were unreactive, whereas the intervening mesenchyme and matrix were positive for d1C4. There was also an abrupt lack of immunoreactivity in the extracardiac mesenchyme of the dorsal mesocardium at the level of the spina vestibuli (Figure 8) that demarcates intrafrom extracardiac mesenchyme.
Discussion
The use of unfixed, air-dried micromass cultures as an in vitro immunogen proved to be a successful method for obtaining MAbs to precartilage aggregate matrix components in their native state, without modification by solubilization or other typical methods of immunogen preparation. The procedure utilized was an adaptation of that described by Gilbert et al. (1990) for the production of MAbs to kidney collecting duct epithelia. Although in vitro immunization frequently results in the production of IgM antibodies, this technique offers several advantages over standard procedures for MAb production, including rapid production of hybridomas, use of small quantities of immunogen, and the ability to override tolerance for conserved immunogens (Reading 1982). The latter two advantages are of particular interest for embryological studies in which quantity or molecular conservation of the immunogen may be problematic for MAb production by traditional methods. However, at present relatively few developmental studies utilizing in vitro immunization have been reported (Gratecos et al. 1987; Milos et al. 1990, 1993; Inai et al. 1993).
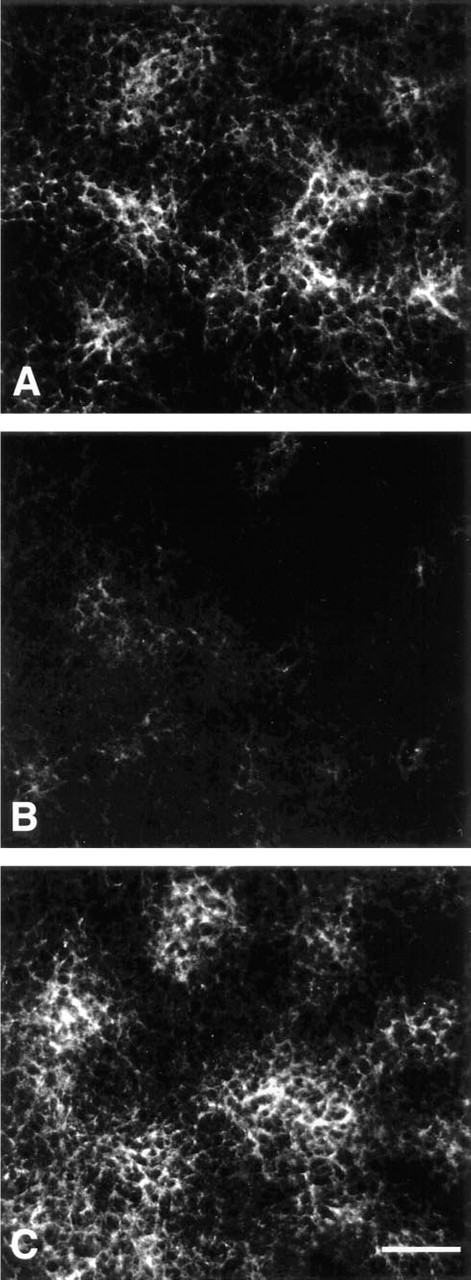
d1C4 immunoreactivity of mesenchymal condensations in micromass cultures of Stage 22–23 limb mesenchyme is blocked by CS-C competition.
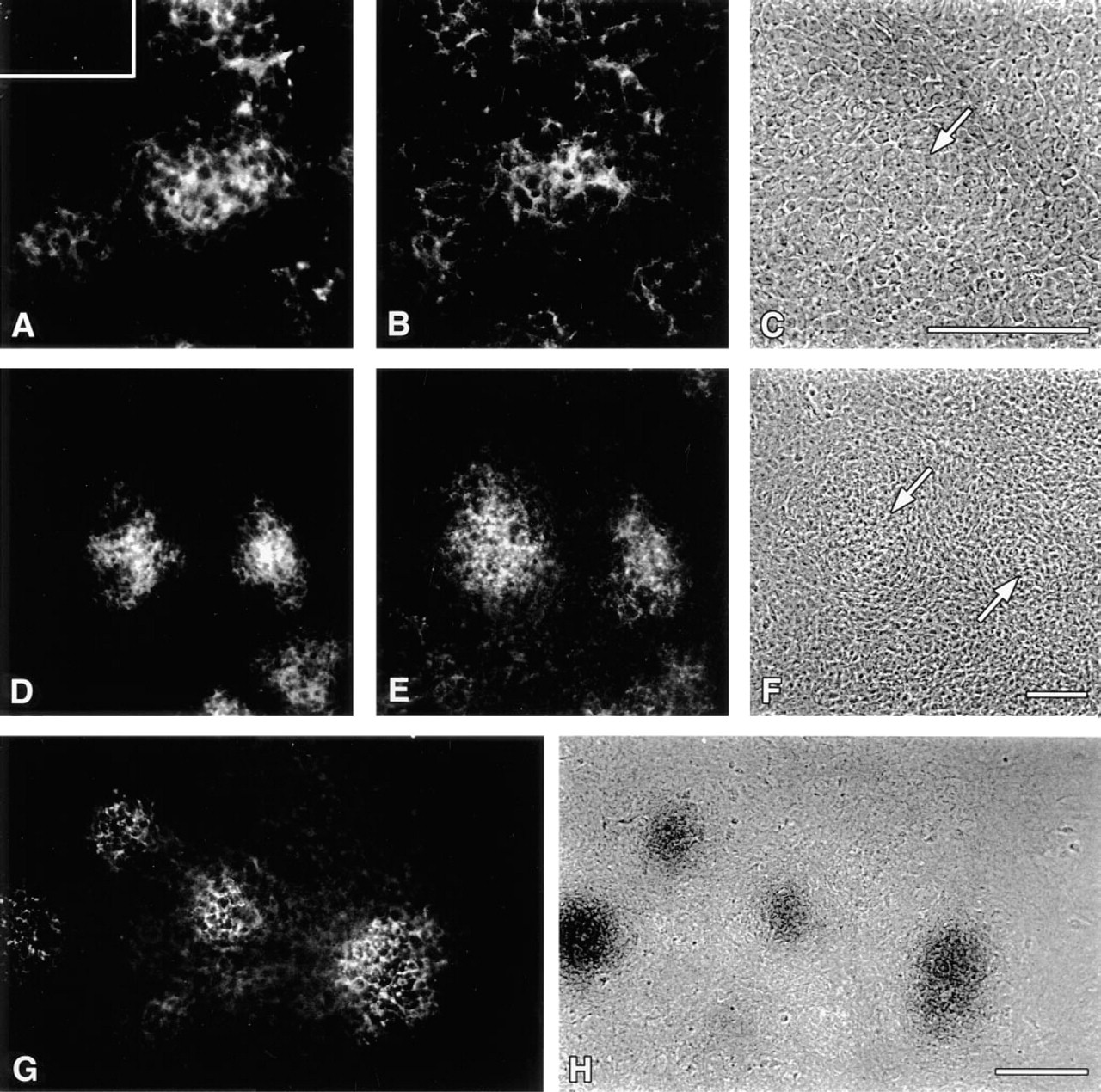
PNA and d1C4 colocalize to prechondrogenic aggregates in micromass cultures of Stage 22–23 limb mesenchyme. RhPNA
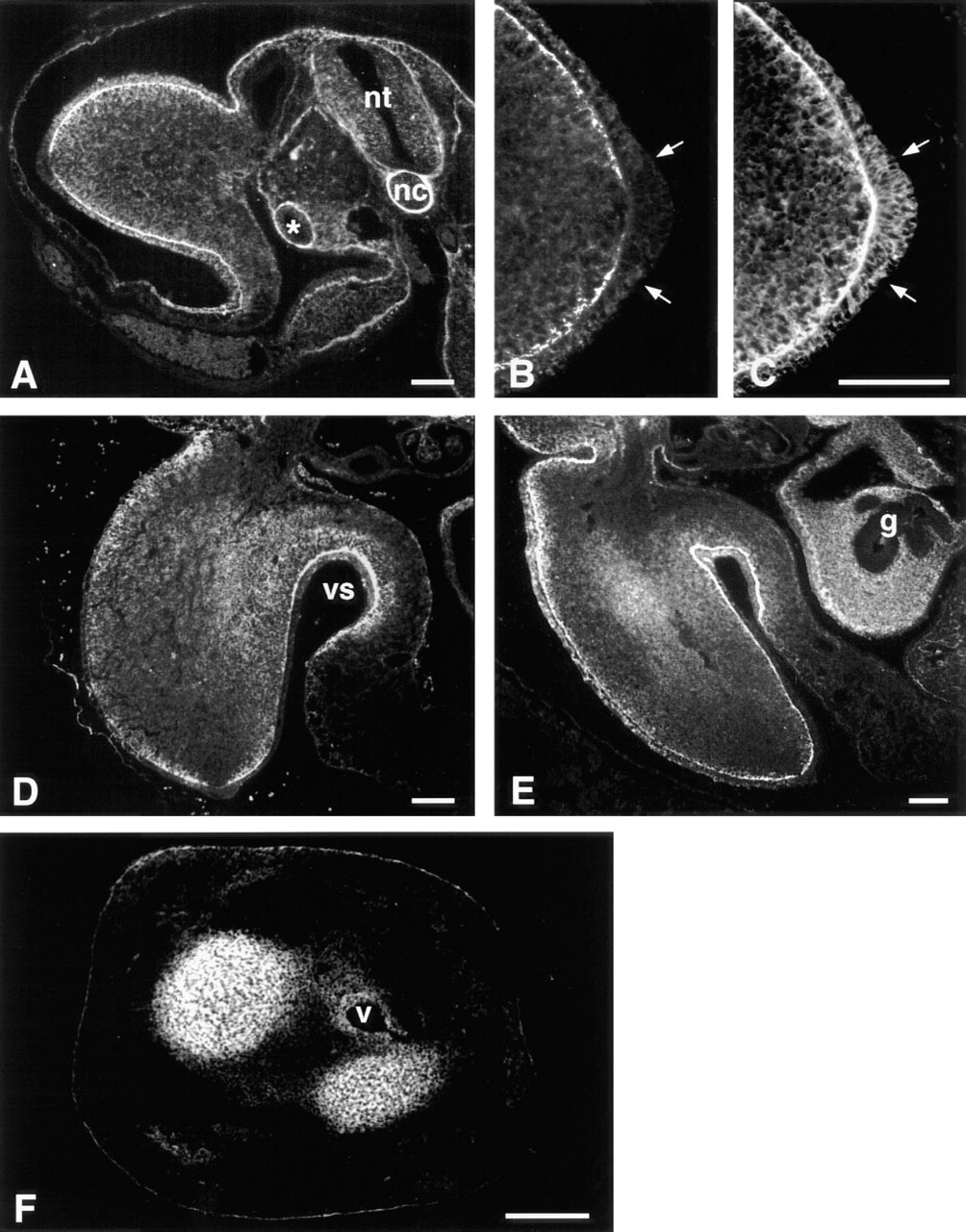
d1C4 immunolocalization in chick limb development.
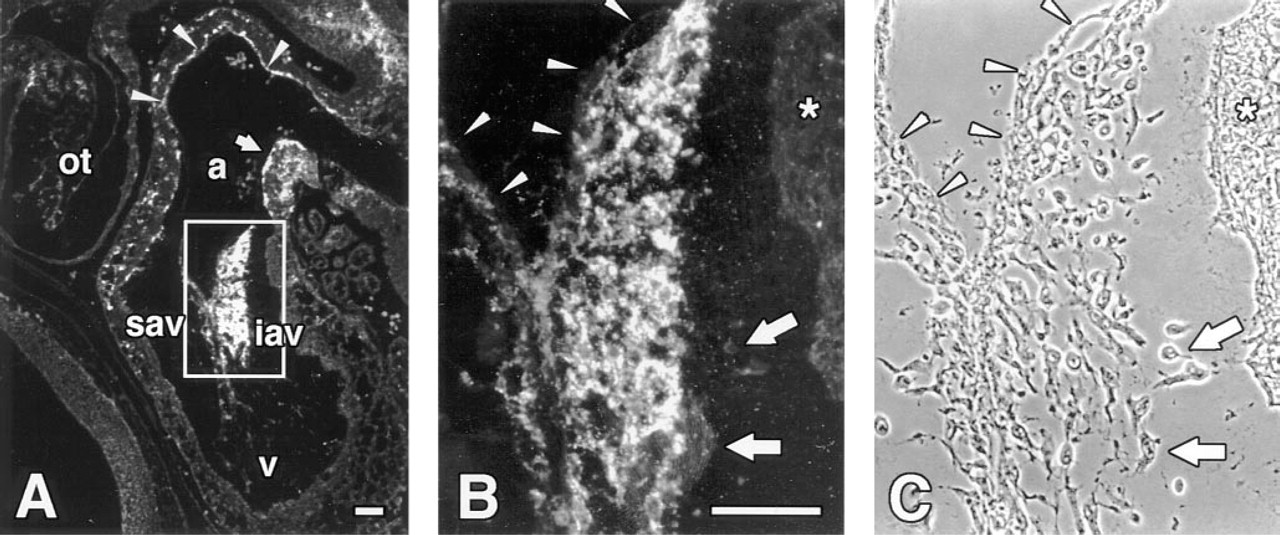
d1C4 recognizes cushion mesenchyme in the Stage 19 chick heart. Sagittal section through the heart of a Stage 19 chick embryo processed with Dent's fixative. Boxed area in
Successful production of hybridomas utilizing the in vitro immunization of mouse lymphocytes in the present study was dependent on several other factors. Micromass cultures fixed lightly with either ethanol or glutaraldehyde (Boss 1986), followed by extensive washing with PBS before the addition of splenocytes, were also utilized but resulted in fewer MAbs reactive with the ECM. The most critical variable was the use of thymocyte conditioned medium as reported previously (Luben and Mohler 1980; van Ness et al. 1984; Reading 1986). The stimulation of lymphocytes was also enhanced by the co-culture of peritoneal macrophages with the splenocyte preparation (Reading 1982) and the inclusion of MDP as adjuvant (Boss 1986). The production of relevant hybridomas was improved by providing splenocytes an opportunity to be challenged for a short period of time in a serum-free environment (van Ness et al. 1984). Omitting one or more of these steps resulted overall in the production of fewer hybridomas.
Several lines of evidence indicate that the epitope recognized by d1C4 resides on GAG chains enriched in chondroitin sulfate type C: (a) d1C4 immunohistochemical reactivity was sensitive to pretreatment with chondroitinase ABC, chondroitinase AC, and testicular hyaluronidase; (b) ELISA results showed a concentration-dependent reactivity with CS-C but not with CS-A GAG; and (c) immunohistochemical reactivity was abolished by preadsorption of d1C4 with CS-C but not with CS-A GAG. In addition, ELISA results showed little or no reactivity of d1C4 with chondroitin, heparan sulfate, keratan sulfate, or dermatan sulfate. The sensitivity of d1C4 reactivity towards preparations digested with chondroitinase AC suggests that it is not crossreactive with dermatan sulfate, nor is the d1C4 epitope a heparan sulfate or keratan sulfate, because these GAGs are not digested by chondroitinases ABC or AC (Yamagata et al. 1968).
Although the data to date are consistent with the d1C4 epitope being found on GAGs enriched in CS-C, the precise structure of the reactive carbohydrate moiety recognized by d1C4 has yet to be identified. However, mild periodate oxidation (Woodward et al. 1985) of high-density micromass cultures did not diminish reactivity with d1C4, suggesting that the epitope either is a terminal carbohydrate sequence resistant to periodate treatment or is a nonterminal component of the CS-GAG chain (Caterson et al. 1990). Individual GAG chains of a CS subtype may be composed predominantly of a single repetitive disaccharide unit, but may also exhibit heterogeneity with regard to length, GAG substitution, or sulfation pattern, resulting in structural differences within a particular CS species, all of which are potential targets for developmental or tissue-specific regulation (Seno et al. 1974; Caterson et al. 1990; Sorrell et al. 1990; Fernandez–Teran et al. 1993). Although the data in the present study indicate preferential reactivity of d1C4 with GAGs enriched in chondroitin-6-sulfate, it should be noted that the commercial CS-C preparations tested contain other CS derivatives, which may be from either CS-A or CS-C GAG chains. This raises the possibility that the CS-GAG epitope recognized by d1C4 could be a derivative of CS-A or CS-C that is not as highly represented in the CS-A preparations utilized. Further investigation will be required to determine whether the epitope recognized by d1C4 resides on the common CS-C unit (glucuronic acid-N-acetyl-
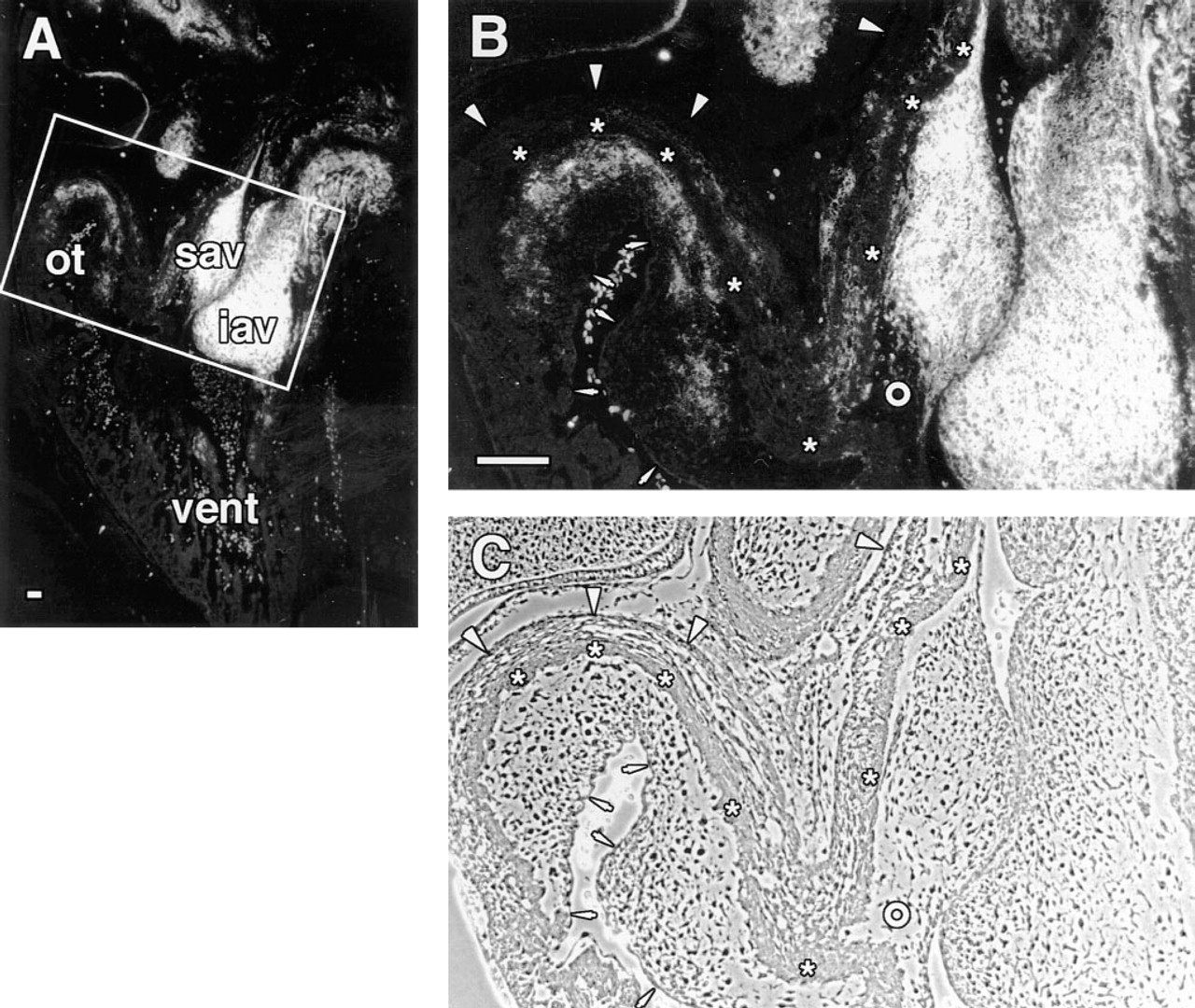
d1C4 staining co-distributes with a subset of mesenchyme cells in the outflow tract of the Stage 25 chick. Sagittal section through the heart of a cryopreserved Stage 25 chick embryo. Boxed area in
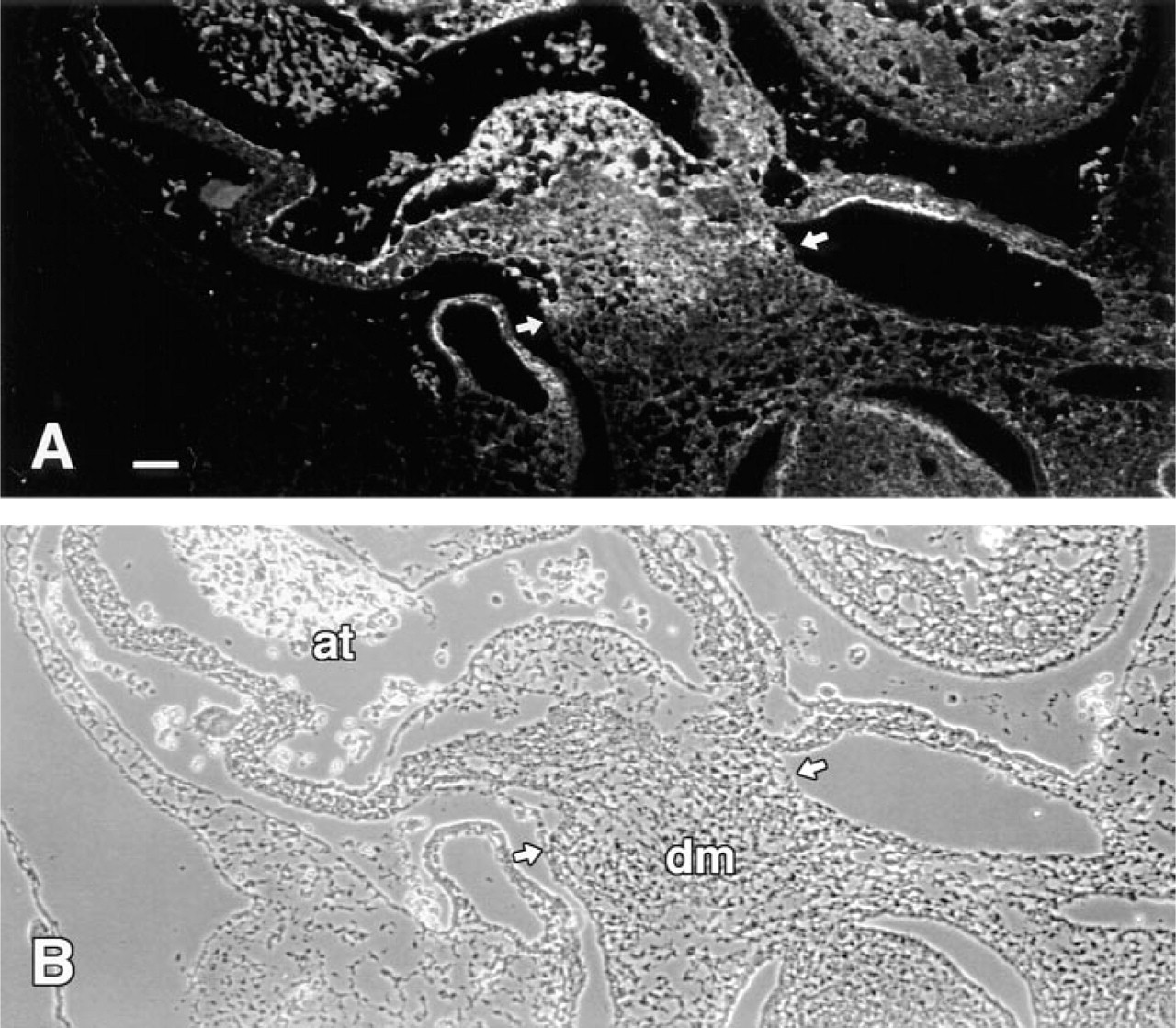
Intracardiac mesenchyme derived from the dorsal mesocardium acquires immunoreactivity with d1C4 in the Stage 22 chick. Cross-section through the atrium (at) of a Stage 22 chick embryo processed with Dent's fixative.
Immunostaining of d1C4 in the developing limb is similar but not identical to that reported for other antibodies to CS-GAG epitopes. Using several MAbs, Fernandez-Teran et al. (1993) described differences in the distribution of a variety of CS-GAG epitopes during limb development. As shown with d1C4 in the present study, the MAb 3B3 (Couchman et al. 1984) recognizes a CS-C epitope in the prechondrogenic core and developing cartilage in the limb, although chondroitinase pretreatment of tissue sections was required for 3B3 reactivity. Interestingly, unlike d1C4, 3B3 does not stain the limb ectodermal basement membrane. MAb MO-225, which recognizes a native disulfated disaccharide sequence on the nonreducing end of CS-GAG chains (Yamagata et al. 1987), stains similarly to d1C4 in the prechondrogenic limb core and ectodermal basement membrane but, unlike d1C4, also stains the AER and subridge basement membrane. MAb CS-56 (Avnur and Geiger 1984), which recognizes a native epitope on both CS-A and CS-C residues, stains the ectodermal basement membrane in the developing limb but not the region subjacent to the AER (Critchlow and Hinchcliffe 1991), similar to that observed with d1C4. Together, these results suggest a heterogeneity in composition and/or accessibility of CS-GAGs in the basement membrane of the limb bud and, in particular, emphasize differences in the subridge basement membrane.
The CS-GAG epitope recognized by d1C4 is likely to be represented on several PGs. PGs are composed of GAG chains linked covalently to a core protein, exhibit a wide tissue distribution, and represent a major component of the ECM (Rouslahti 1988). The localization of several PG core proteins has been described in the prechondrogenic limb (Kimata et al. 1986; Shinomura et al. 1990; Solursh et al. 1990; Nishiyama et al. 1991; Gould et al. 1992), and a heterogeneous distribution of chondroitin sulfate/dermatan sulfate GAGs in the embryonic chick limb bud has been reported as well (Fernandez–Teran et al. 1993). Fernandez–Teran et al. (1993) have demonstrated dynamic spatial and temporal changes in the localization of chondroitin sulfate/dermatan sulfate epitopes during limb development that may reflect different functional roles for individual PGs. We have not attempted in the present study to identify the core proteins of chondroitin sulfate proteoglycans recognized by d1C4, but PG-M, which has been localized to the prechondrogenic core of the developing limb (Kimata et al. 1986; Shinomura et al. 1990), is a likely candidate in that location.
Of particular interest in the present study was the spatial and temporal localization of d1C4 reactivity in prechondrogenic aggregates of chick limb mesenchyme that enabled us to monitor the process of mesenchymal condensation in culture. In the limb, chondrogenesis is preceded by the aggregation of mesenchymal cells into cellular condensations (Fell 1925), which are of paramount importance in the establishment of limb primordia (Hall and Miyake 1992). Condensation of precartilage mesenchyme represents a requisite step during chondrogenesis and aggregates must achieve a threshold size before cell differentiation can proceed (Solursh et al. 1978; Hall and Miyake 1992). Prechondrogenic aggregate formation is a multifactorial process and several ECM molecules have been implicated, including Type I collagen (Dessau et al. 1980), fibronectin (Dessau et al. 1980; Downie and Newman 1995), PG-M (Kimata et al. 1986), and hyaluronate (Toole et al. 1972; Knudson and Toole 1987). Previously, RhPNA, which recognizes terminal galactose-N-acetylgalactosamine residues generally found on glyco-proteins, has been demonstrated to demarcate reliably precartilage aggregates in cultured chick limb mesenchyme by 16 hr (Aulthouse and Solursh 1987). As reported in the present study, d1C4 reactivity overlapped with that of RhPNA in prechondrogenic condensations after 16 hr of culture and was detected throughout the differentiation of aggregates into cartilage nodules. However, a difference often observed in the staining pattern between RhPNA and d1C4 during the initial phases of culture was that PNA binding molecules appeared more restricted to the microenvironment of the aggregate, whereas d1C4 reactivity was observed consistently at earlier time points and extended into the peripheral mesenchyme, perhaps indicative of the recruitment of surrounding mesoderm into the aggregate (Ede 1983; Cottrill et al. 1987). Although PNA and d1C4 stain different carbohydrate structures, our data do not necessarily preclude their recognition of the same macromolecule.
Many of the ECM molecules expressed in precartilage aggregates of the developing limb are also expressed in the developing endocardial cushions, including Type I collagen (Johnson et al. 1974; Thompson et al. 1979), fibronectin (Icardo and Manasek 1984; Kitten et al. 1987), hyaluronate (Orkin and Toole 1978), and CS-GAGs (Markwald et al. 1978, 1979; Colvee and Hurle 1981). The atrioventricular (AV) and outflow tract (OT) endocardial cushions exist initially as an expansive acellular ECM derived in part from the adjacent myocardium (Markwald et al. 1978; Krug et al. 1985). This ECM is invaded by mesenchyme originating from a subpopulation of endocardial cells that have undergone an epithelial–mesenchymal transformation (Markwald et al. 1975) that is regionally specific for the AV and OT. Funderburg and Markwald (1986) showed biochemically that the ratio of CS-C to CS-A decreases in both the AV and OT during cushion tissue formation due to conditioning of the ECM with a mesenchyme-derived chondroitin sulfate proteoglycan (presumably CS-A-rich) that restricts the migration of trailing invasive cells. In the present study, expression of the CS-GAG epitope recognized by d1C4 was essentially absent in the premigratory matrix and increased markedly as AV mesenchyme was formed. Until recently, little has been done to examine the in situ expression pattern of individual CS-GAG chains. Using two MAbs that recognize subpopulations of CS-GAGs, Funderburg et al. (1991) showed regional differences in certain CS-GAGs within the endocardial cushions. CS-56 (Avnur and Geiger 1984), which recognizes an epitope on both CS-A and CS-C chains, stained the premigratory acellular ECM. However, an antibody (MO-225; Yamagata et al. 1987) that recognizes a disulfated CS-D epitope stained only those regions of the AV cushions that contained invasive mesenchyme. In the present study, the CS-GAG epitope recognized by d1C4 was expressed throughout the AV endocardial cushions of the Stage 25 chick embryo. However, mesenchyme within the OT stained differentially with d1C4, indicative of two phenotypically distinct mesenchymal populations. It is uncertain if these different populations of mesenchyme represent separate contributions of endothelially derived mesenchyme and neural crest cells to this region of the OT (Kirby et al. 1983; Miyagawa–Tomita et al. 1991; Noden 1991) or the localized differentiation of a subset of a previously homogeneous mesenchyme. Perhaps this polarization of staining could represent a “territorial boundary” for invasion of neural crest into this region, similar to that observed by Oakley et al. (1994). Another interesting feature demonstrated by d1C4 immunostaining was the abrupt change in the expression of this CS-GAG epitope at the interface between the intracardiac and contiguous extracardiac mesenchyme of the dorsal mesocardium. This region of the heart is important because it serves as the entrance point for the spina vestibuli, an extracardiac mesenchymal structure at the venous pole of the developing heart that invades the heart to contribute to atrial septation (Asami and Koizumi 1995; Tasaka et al. 1996).
Functions attributed to PGs are numerous and range from organization of the ECM (Oldberg and Rouslahti 1982; Kleinman et al. 1986) to a role in the control of cell division (Grammatikakis et al. 1995) and the inhibition (Funderburg and Markwald 1986; Perris and Johansson 1987) or facilitation (Lane and Solursh 1991) of cell migration. The striking localization pattern of d1C4 at sites of critical morphogenetic events underscores the importance of future studies to identify the precise structure of its CS-GAG epitope. Such information will aid in identification of the enzymes responsible for its synthesis, which are likely to be tightly regulated during development. The fact that d1C4 immunoreactivity does not require enzymatic pretreatment may make this an ideal reagent for functional bioassays as well. Studies are under way to examine the functional role of the d1C4 antigen in limb and heart development.
Footnotes
Acknowledgements
Supported by grants from the NIH to EK (HL52813), MS (HD05505), and GK (HL42266) and from the AHA to MW (94004580).
We thank Sue Tjepkema–Burrows and John Busey for photographic assistance, Dr Tom Trusk and Josh Spruill for help in preparing the figures, and Jan King, Sandra Kolker, Tanya Rittmann, and Alison Chilton for preparation of tissue sections. Thanks also to Dr Roger Markwald for insightful discussion and critical comments and to Dr Corey Mjaatvedt for suggesting poly-