
Abstract
In a comprehensive study, we examined the expression of the membrane and secretory mucins MUC1 and MUC3, respectively, in normal and neoplastic gastrointestinal and breast epithelia before and after specific alterations of their glycan structures by neuraminidase, α-fucosidase, or carbohydrate-specific periodate oxidation. MUC1 mRNA was also identified in normal colorectal tissues by in situ hybridization. The data revealed that normal colorectal epithelia express both MUC1 mRNA and protein, which were detectable after periodate oxidation with all tested MUC1-specific antibodies. During tumorigenesis in the colon, MUC1 became recognizable without periodate treatment concomitantly with highly dysplastic lesions and the malignant state. In the breast, in which MUC1 is detectable with most antibodies in normal epithelium as well as in carcinomas, staining could be enhanced by pretreatment with periodate and casually by enzyme treatments. MUC3 was detectable in normal and neoplastic colorectal tissues and was more intensely stained after periodate oxidation. It was absent in normal breast even after pretreatment but was expressed in seven of 20 breast carcinomas. Therefore, incomplete glycosylation, abnormal distribution, and ectopic expression of mucins are characteristics of malignancy. Periodate oxidation may be widely applicable to immunohistochemistry for examining changes in glycosylation and for detecting antigens masked by glycans.
Keywords
M
In this study, serial sections from various gastrointestinal (normal stomach, ileum, and colon; colorectal adenomas and carcinomas) and mammary tissues (normal breast and mammary carcinomas) were used to examine the presence of MUC1 mRNA in the different cell types of normal gastrointestine by in situ hybridization and the expression of MUC1 and MUC3 proteins, the latter as an example of a secretory mucin, by immunohistology with MAbs to peptide epitopes before and after glycan-intruding measures such as mild periodic acid oxidation, α-fucosidase, or neuraminidase incubation. The results show that MUC1 is indeed expressed in normal colorectal mucosa but is inaccessible to anti-MUC1 peptide-recognizing MAbs. Epitope masking by glycans can be overcome by pre-treatment of the tissue sections with periodate.
Materials and Methods
Normal Adult Human Tissues and Tumor Specimens
Two stomach (including corpus and pyloric antrum), one ileum, and four colon specimens were obtained at autopsy from individuals without colon disease. One ileum and 15 colon tissue samples were taken at surgery or colonoscopy from apparently normal parts of the colon wall distant from the tumor. All mucosae showed normal histomorphological and cytomorphological findings. Twenty-three cases of colorectal adenomas and 22 carcinomas were obtained at surgery or colonoscopy. Adenomatous polyps were classified as to histological type and grade of dysplasia according to the criteria of Konishi and Morson (1982). Classification of primary carcinomas was done with hematoxylin–eosin-stained sections according to World Health Organization (WHO) recommendations (Jass and Sobin 1989).
Twenty mammary carcinomas and 10 normal mammary tissues from patients with cancer were derived from surgical specimens. The tumors were classified according to the criteria of the WHO (1982) and the Bloom–Richardson system (Elston and Ellis 1991). The ABO blood group type and the secretor status of the donors were also documented for all mammary tissues.
Two of the normal stomach, ileum, and colon samples and all mammary samples were fixed in 10% buffered formalin and embedded in paraffin. All other tissues were immediately frozen and stored at −80C.
In Situ Hybridization
RNA Probe. The MUC1 cDNA probe, pum24P (Swallow et al. 1987), was sequenced. A 171-
In Situ Hybridization (ISH). ISH was performed as described (Schaeren–Wiemers and Gerfin–Moset 1993) in two samples each of normal stomach, ileum, and colon. Deparaffinized sections or formalin-fixed cryosections were digested with proteinase K (Boehringer; Mannheim, Germany) [10 μg/ml in PBS, 20 min at room temperature (RT)]. After three rinses in PBS, sections were treated with 0.5% and 1% triethanolamine, pH 8.0, and 0.25% acetic anhydride for 5 min. Prehybridization was done with hybridization buffer [50% formamide, 0.75 M NaCl, 0.075 M sodium citrate, pH 7.0 (5 x SSC), 0.1% bovine serum albumin, 0.1% Ficoll, 0.1% polyvinyl pyrrolidone, 0.25 mg/ml tRNA, 1.25 mg/ml herring sperm DNA, 4 mM EDTA] at 45C for 3 hr. The hybridization mixture was prepared by adding 1 μl DIG-RNA probe per 50 μl hybridization buffer, heated for 5 min at 85C to denature the probe, and then chilled on ice. Fifty μl of this hybridization mixture was added per slide, which was then covered with a coverslip and sealed with rubber cement. Hybridization was done overnight at 72C. Posthybridization washings were successively performed in SSC solutions of decreasing concentrations (5 x SSC, 50% formamide, 10 min at 50C; 2 x SSC, 25% formamide, three times for 10 min at 42C and twice for 30 min at 42C; 0.2 x SSC, twice for 5 min at RT). Thereafter, the sections were rinsed three times with PBS and incubated with Fab fragments from anti-DIG antibody conjugated to alkaline phosphatase (Boehringer; 1:500) for 1 hr at RT. After three washing steps, the color development was performed with the Fast Red Substrate System (DAKO; Hamburg, Germany). Controls for in situ hybridization consisted of (a) DIG-labeled sense RNA probe, (b) competition with a 20-fold excess of unlabeled anti-sense RNA in the presence of DIG-labeled anti-sense RNA, (c) pretreatment of sections with ribonuclease, and (d) hybridization mixture without RNA probe.
Immunohistochemistry
Monoclonal Antibodies. The antibody A76-A/C7 (IgG1k ) was developed in this laboratory (Cao et al. 1997a,b). It recognizes the peptide epitope APDTRP of the MUC1 tandem repeat (O. Nilsson et al., communicated at the ISOBM TD-4 Workshop, San Diego, CA, November 17–22, 1996). The anti-MUC1 antibody HMFG-2 (IgG1) recognizing the epitope DTR (Burchell et al. 1989) was purchased from Dianova (Hamburg, Germany). SM3 (IgG1) specific for the MUC1 epitope PDTRP (Burchell et al. 1989) was kindly donated by Dr. G. Pecher (MDC). M3.2 (anti-MUC3, IgG2) (Apostolopoulos et al. 1995) recognizes a peptide epitope of MUC3. MAbs to group A and B antigens were purchased from DAKO. MAbs A46-B/B10 (IgM), specific for the H Type 2 antigen (Karsten et al. 1988), and A78-G/A7 (IgM), specific for the Thomsen–Friedenreich antigen (Karsten et al. 1995), have been described.
Tissue Staining. Staining of tissue sections was performed by the avidin–biotin–peroxidase complex (ABC) method with a commercial kit (Vectastain ABC Elite Kit; Vector Laboratories, Burlingame, CA) as follows. Paraffin sections (4 μm) were deparaffinized in xylene and rehydrated through a graded ethanol series. Frozen sections (4 μm) were air-dried at RT and fixed with 10% formalin in PBS for 15 min at RT. Endogenous peroxidase activity was eliminated by treatment with 3% H2O2 in PBS for 30 min at RT. Nonspecific binding sites were blocked with normal rabbit serum. After washing with PBS, sections were incubated with MAbs in appropriate dilutions for 1 hr at RT. The thoroughly washed sections were treated with biotinylated anti-mouse immunoglobulin antiserum for 30 min at RT and thereafter with the ABC complex. Color development during incubation with the peroxidase substrate (diaminobenzidine) was controlled under the microscope. Counterstaining was done with hematoxylin. Negative controls were incubated with comparable dilutions of IgM or IgG paraproteins from mouse plasmocytomas (Sigma; Deisenhofen, Germany) instead of the MAb.
Partial Deglycosylation. One of the following pretreatments was performed on the sections before incubation with anti-MUC1 or MUC3 MAbs.
Neuraminidase. Sections were incubated with neuraminidase from Vibrio cholerae (Serva; Heidelberg, Germany) at a concentration of 0.02 U/ml in PBS containing 0.01 M Ca++ for 1 hr at RT to remove sialic acid residues. After enzyme treatment, sections were rinsed for 5 min in PBS. Positive control of neuraminidase action was achieved by staining of erythrocytes and endotheliocytes of a tissue section by MAb A78-G/A7 (anti-Thomsen-Friedenreich antigen) after the enzyme treatment. α-Fucosidase. Sections were incubated for 24 hr at 37C with α-fucosidase (bovine kidney, Boehringer; 0.4 U/ml in acetate buffer, 0.2 M, pH 5). Then they were rinsed for 5 min in PBS. Loss of staining of erythrocytes and endotheliocytes in tissue sections from individuals with blood group O with the anti-H2 MAb A46-B/B10 served as control of enzyme action.
Periodate Oxidation. Sections were treated as described (Bara et al. 1992). Briefly, they were incubated for 30 min at RT with 20 mM periodic acid in acetate buffer, 0.05 M, pH 5. After three rinses in PBS, the sections were treated with 1% glycine for 30 min, followed by three rinses in PBS. This treatment destroyed glycotopes containing galactose, N-acetylgalactosamine, or fucose in cell preparations or tissue sections, but did not damage peptide epitopes (Cao et al. 1997a).
Blood Groups and Secretor Status. Bara et al. (1993) reported that in normal gastric mucosa the immunodetection of MUC1 in secretors was different from that in nonsecretors. We therefore collected blood group and secretor data on patients. In breast cancer patients, ABO data were available from the clinical records. The secretor status was determined from saliva according to standard procedures. Because in the gastrointestinal and normal breast tissue donors neither their blood group nor their secretor status was known to us, we applied anti-H2, anti-A, and anti-B MAbs to some sections, and examined normal epithelial cells, red blood cells, and endothelia for reactivity. According to the results, secretor status and blood group of the donors were determined.
Scoring. For normal mucosa, the percentage of positive crypts was counted, whereas in carcinomas the percentage of positive cells in several optical fields (12.5 x lens) was estimated. For further details, see legends to Figures 2–4 and 6.
Results
Expression of MUC1 mRNA in Normal Gastrointestinal Epithelium
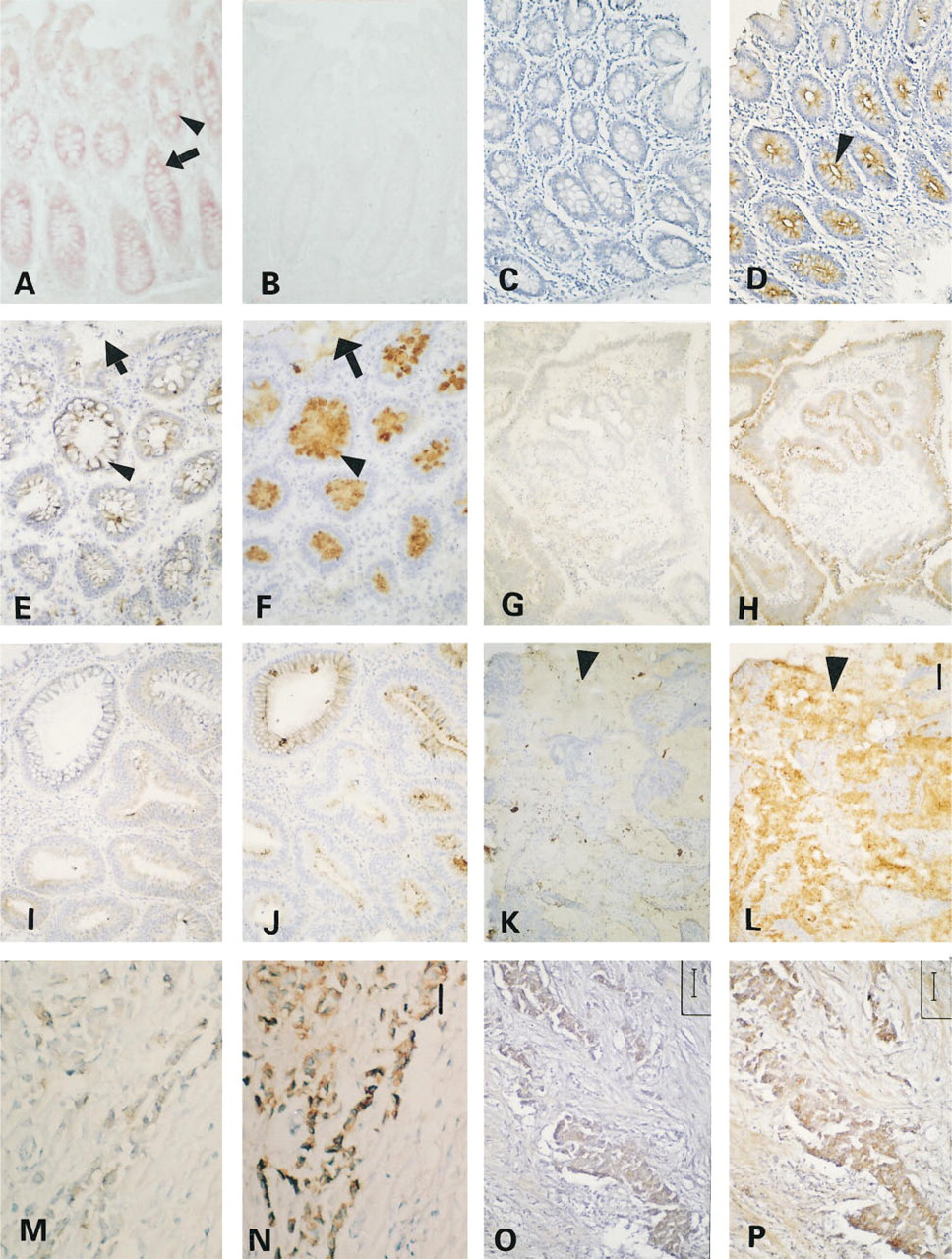
By in situ hybridization, MUC1 mRNA was found to be expressed in surface epithelial cells and in gastric glands of the stomach. In all of two ileum and four colon specimens, the DIG-labeled MUC1 anti-sense RNA probe intensely stained epithelial cells, columnar cells and goblet cells (Figure 1A). The staining corresponded to the cytoplasm of columnar cells and to the perinuclear region of goblet cells. DIG-labeled sense probes did not hybridize to any tissue structure (Figure 1B). The other control experiments also showed no staining.
Expression of Apomucin Along the Normal Gastrointestinal Tract Before and After Partial Deglycosylation
The staining patterns with MUC1 mAbs before and after partial deglycosylation are shown in Figures 1 and 2. Analysis of the stained sections revealed that A76-A/C7 and HMFG-2 did not stain, as expected, any of the following untreated normal tissue sections: two stomach specimens from secretor individuals, two ileum samples from nonsecretor individuals, and 19 colon specimens (including two from ascending colon) from nonsecretor individuals (Figure 1C). SM3 behaved identically except for weak staining of the supranuclear region of columnar cells and goblet cells in one case from colon. In contrast to this, MUC1 MAbs strongly labeled all normal gastrointestinal epithelia after periodate treatment, but in most cases their secretions remained unstained. Enzymatic pretreatment with α-fucosidase led to MUC1 staining exclusively in sections of the two stomach specimens, which were both secretors. Neuraminidase was without effect in all normal gastrointestinal tissues examined. Staining patterns after periodate oxidation varied with the antibody employed. A76-A/C7 staining was predominantly distributed at the apical membranes of surface mucous cells of the stomach and of columnar cells and goblet cells of the intestine (Figure 1D), whereas HMFG-2 and SM3 staining was localized in the supranuclear cytoplasm of surface mucous cells of the stomach and in the perinuclear region of columnar cells and goblet cells of the intestine. All MUC1 MAbs diffusely stained gastric glands proper and pyloric glands.
MUC3 positivity in untreated normal tissue sections from the stomach was restricted to occasional cytoplasmic staining of the surface epithelium of stomach and gastric glands. In normal ileum and colon, M3.2 stained the supranuclear cytoplasm of goblet cells and columnar cells, but not secretions (Figure 1E). After periodate oxidation, mucous cells, gastric glands proper, and pyloric glands, as well as secretions in the stomach became positive. Vacuoles of goblet cells and secretions in the ileum and colon were also positive (Figure 1F).
(
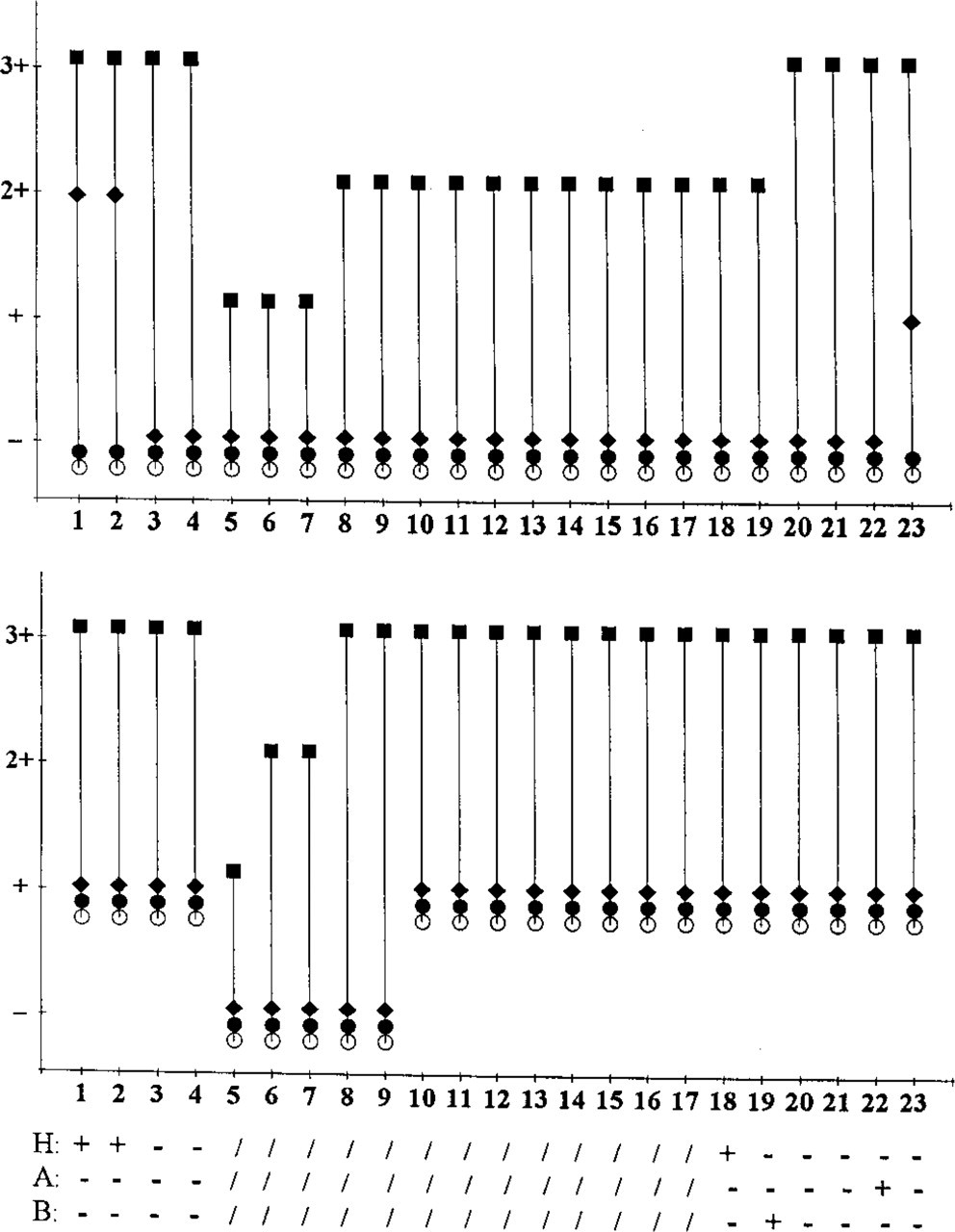
Comparison of the reactivity of anti-MUC1 (
Expression of Apomucin in Colorectal Adenomas and Carcinomas Before and After Partial Deglycosylation
The results of mucin staining in adenomas and carcinomas with and without partial deglycosylation are shown in Figures 1, 3, and 4. For colorectal adenomas and carcinomas, similar results were obtained with all three anti-MUC1 MAbs, A76-A/C7, HMFG-2, and SM3. In Figures 3 and 4, data for A76-A/C7 are shown. The anti-MUC1 MAbs did not stain mildly or moderately dysplastic adenomas, with the exception of a few weak reactions. As expected, staining was greatly enhanced in all cases after periodate treatment. These MAbs, however, reacted with untreated sections of severely dysplastic adenomas; this was enhanced to a certain degree after periodate treatment (Figures 1G and 1H). Adenocarcinomas were generally reactive, but periodate treatment still slightly enhanced the reactivity. After α-fucosidase treatment slightly increased staining could be seen. In most cases, secretions in benign lesions did not react with MUC1 MAbs, even after periodate treatment. In contrast, secretions of some carcinomas showed variable (from slight to moderate) reactivity with MUC1 MAbs, and this was increased after periodate treatment. Neuraminidase treatment did not lead to significantly increased staining of MUC1 in gastrointestinal tumor cells and secretions.
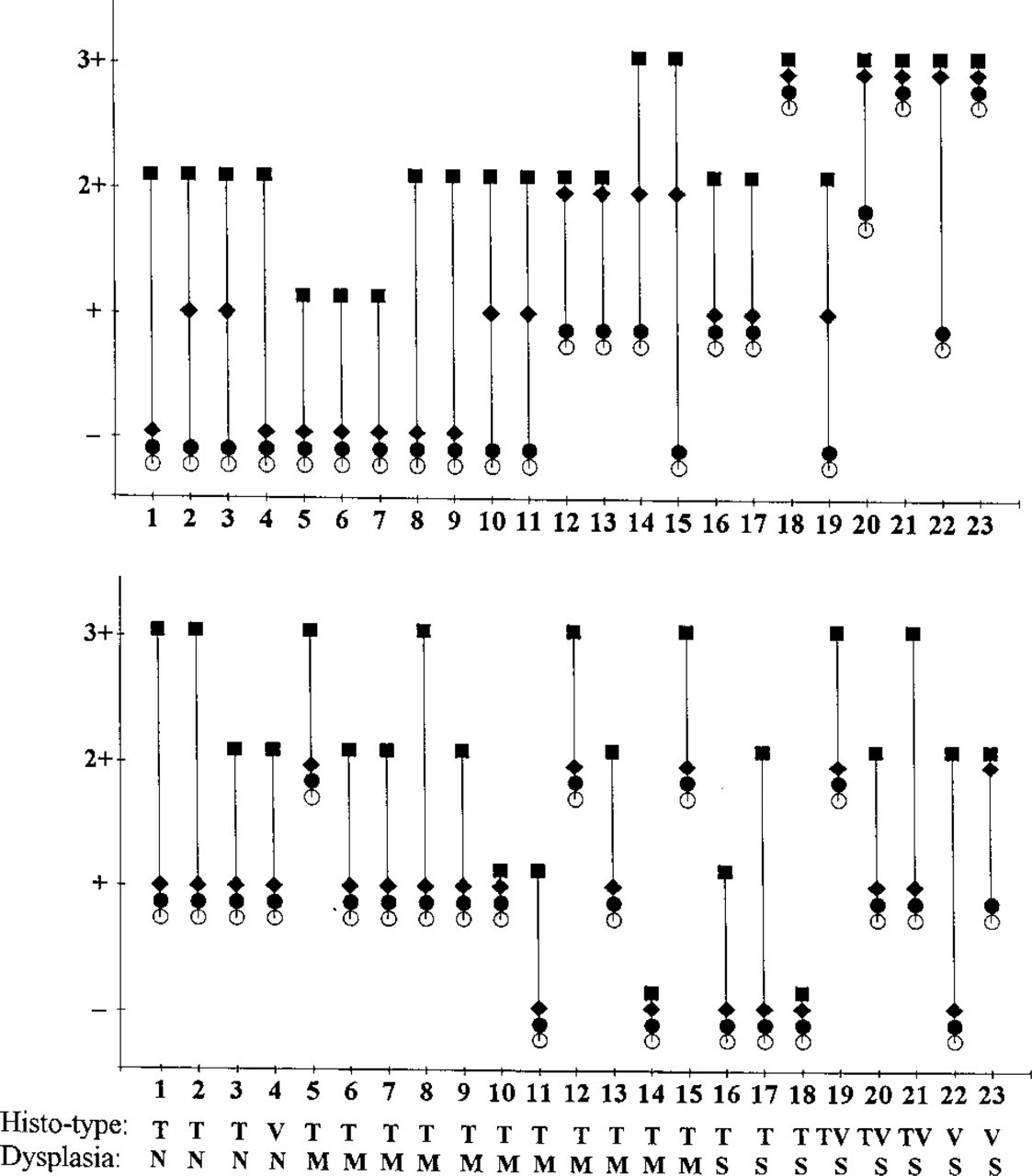
Comparison of the reactivity of anti-MUC1 (
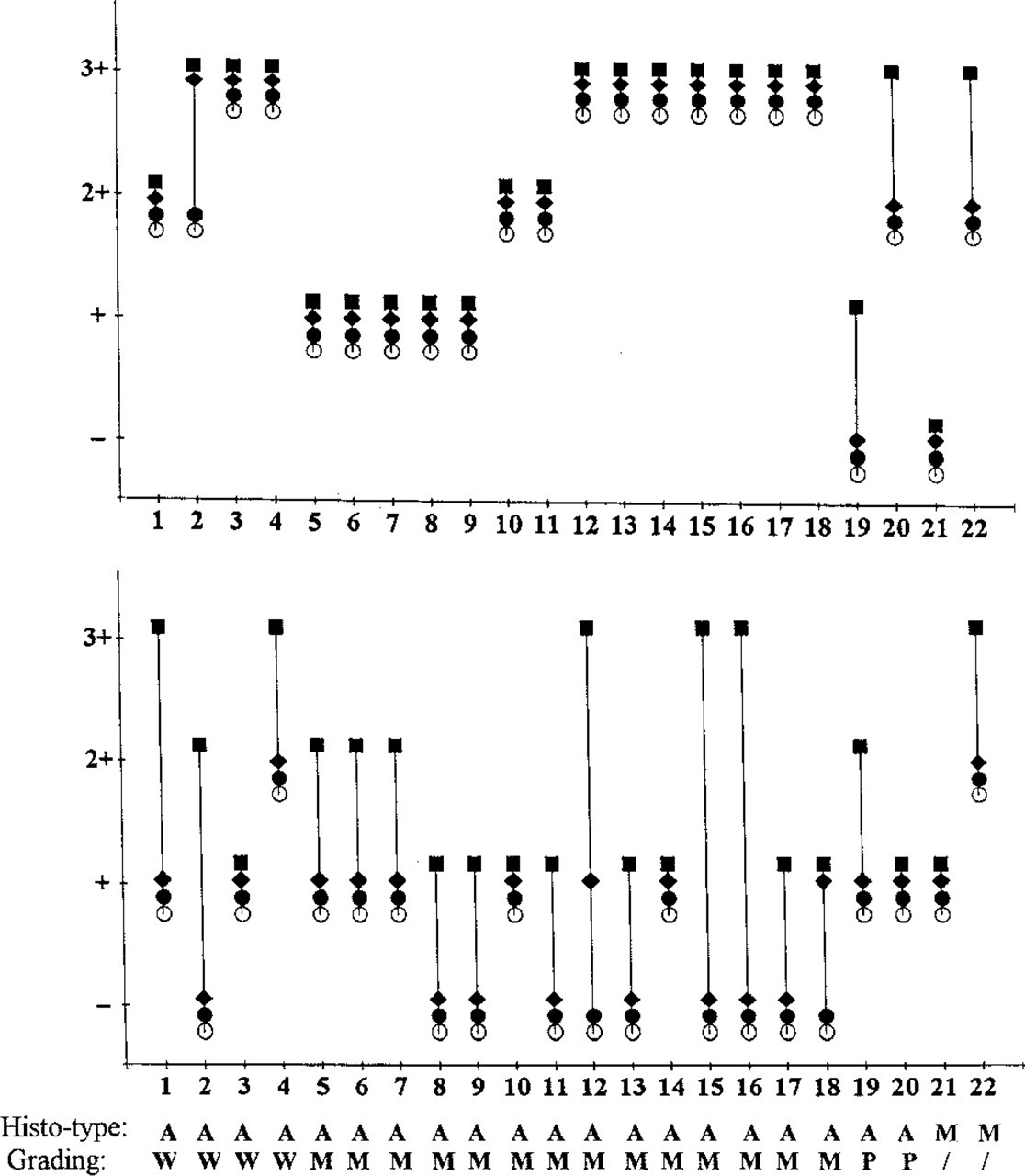
Comparison of the reactivity of anti-MUC1 (
Staining of MUC3 apomucin in colorectal benign and malignant lesions and secretions was significantly enhanced after periodate treatment (Figures 1I and 1J). A slightly increased reaction of MUC3 mAb with several severely dysplastic adenomas and carcinomas was observed after α-fucosidase treatment. Neuraminidase treatment did not enhance staining of MUC3 in gastrointestinal tumor cells and secretions.
In adenomas with mild or moderate dysplasia, MUC1 and MUC3 (native and deglycosylated) were localized in the supranuclear cytoplasm, at the apical membrane, and/or in the apical cytoplasm of tumor cells (Figures 1E, 1F, 1I, and 1J). In carcinomas and adenomas with severe dysplasia, both mucins were observed either diffusely distributed in the whole cell or at the basolateral membrane.
Figure 5 provides a somewhat simplified overview of the staining results during the stages of colorectal carcinogenesis.
Expression of Apomucin in Normal Mammary Tissues Before and After Partial Deglycosylation
Normal mammary gland sections were reacted with A76-A/C7 and HMFG-2 before and after partial deglycosylation (data not shown). Staining was observed in all cases; it was restricted to the apical membranes of epithelial cells. Antibody SM3 did not react with normal mammary gland tissues (independent of whether secretors or nonsecretors), and remained nonreactive after neuraminidase or α-fucosidase treatment. After periodate treatment, however, SM3 reacted with normal mammary gland tissue at the apical membranes of epithelial cells, similar to the localization seen with A76-A/C7 and HMFG-2.

Comparison of staining results obtained with anti-MUC1 and anti-MUC3 antibodies on various colorectal tissues. Untreat, untreated; PO, periodate oxidation; N'ase, neuraminidase incubation; F'ase, α-fucosidase incubation. Scoring: -, no staining; +, weak staining; + +, moderate staining; + + +, strong staining. Tendency of treatment effect in parentheses: =, staining not increased; ↑, staining slightly increased; ↑↑, staining moderately increased; ↑↑↑, staining strongly increased. MUC1 immunohistochemistry results as obtained with MAb A76-A/C7 or HMFG-2.
All normal mammary tissues were negative for MUC3 with or without partial deglycosylation.
Expression of Apomucin in Breast Carcinomas Before and After Partial Deglycosylation
The staining results with anti-mucin MAbs in mammary carcinoma sections with and without partial deglycosylation are shown in Figures 1 and 6. The stain-ability of MUC1 in mammary carcinoma cells was strongly increased after periodate oxidation (Figures 1M and 1N). Neuraminidase treatment of the sections also distinctly increased the reactivity of MUC1 with the respective MAbs, whereas α-fucosidase slightly increased staining. Three patterns of MUC1 localization could be distinguished: (a) the antigen was present at the apical membranes of tumor cells in glandular adenocarcinomas; (b) the antigen was diffusely distributed in the cytoplasm of tumor cells in non-gland-forming areas of tumors; and (c) in some poorly differentiated carcinomas, a proportion of positive cells exhibited distinct granular cytoplasmic staining (focal cytoplasmic staining).
Unexpectedly, five of 20 mammary carcinomas expressed MUC3. Two further cases became positive after periodate oxidation, and in one case the staining increased after this treatment (Figures 1O and 1P). Native and deglycosylated MUC3 was diffusely distributed in the cytoplasm of tumor cells. Neuraminidase and α-fucosidase treatments had no influence on the reactivity (except in one case).
Expression of MUC1 and MUC3 apomucins before and after partial deglycosylation was not related to the secretor status of the patients.
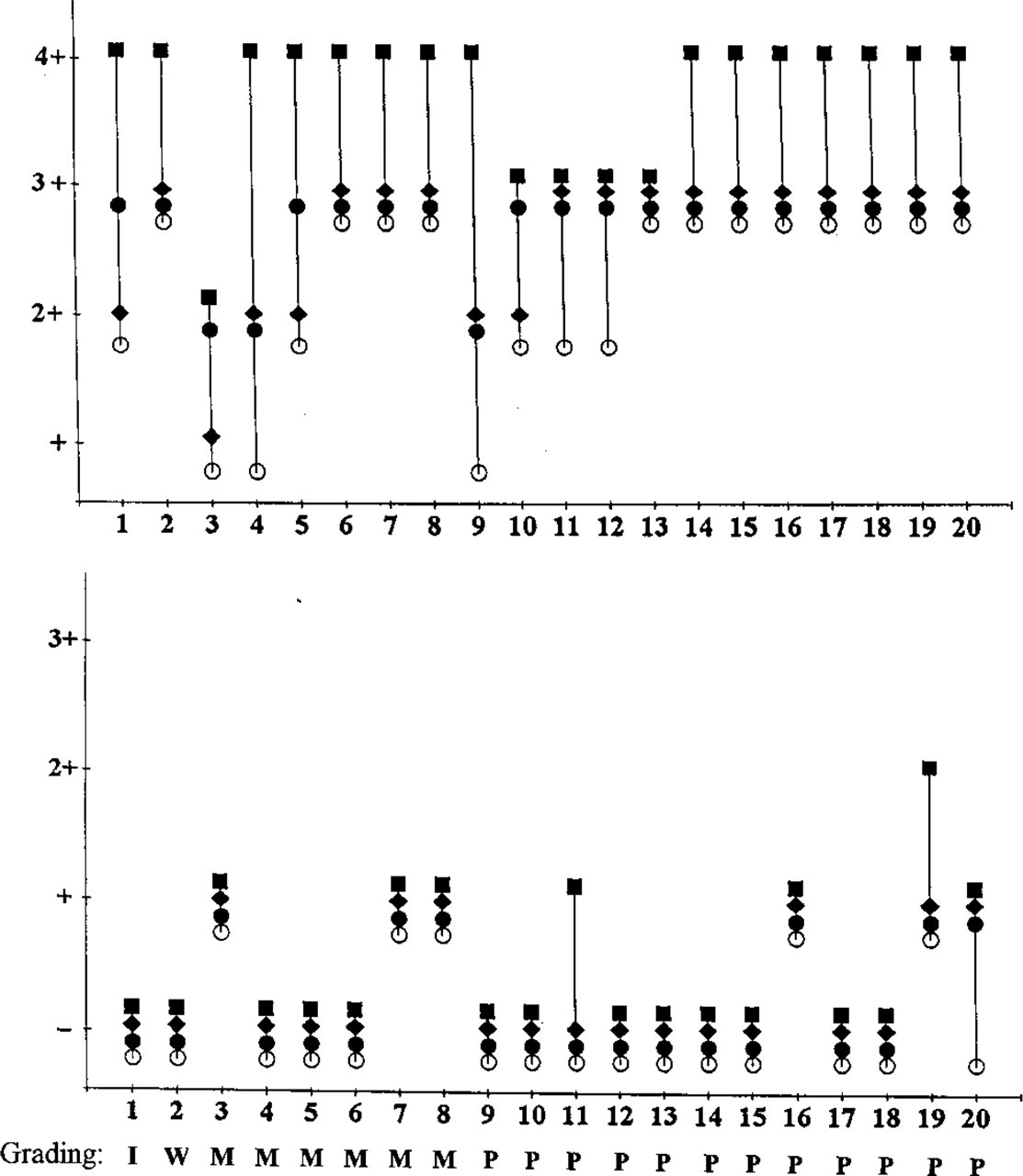
Comparison of the reactivity of anti-MUC1 (
Discussion
A common feature of glandular epithelial tissues is the apical (luminal) expression and secretion of mucins. MUC1 is generally assumed to be the major mucin expressed in normal breast, pancreas, stomach, and other glandular epithelia, but not or only sparsely in the intestine (Zotter et al. 1988; Girling et al. 1989; Ho et al. 1993; Jass and Roberton 1994). However, the puzzle is that normal adult intestinal epithelia have been recently shown to express MUC1 mRNA (Ogata et al. 1992; Ho et al. 1993; Nakamori et al. 1994), and a few reports claim that the normal adult intestine could be stained by certain MUC1 MAbs (Carrato et al. 1994; Hilkens et al. 1995). From our earlier studies, it was already known that during colorectal tumor development MUC1-associated glycotopes [e.g., Thomsen-Friedenreich antigen (Galβ1–3GalNAcα1-O-Ser/Thr), Tn (GalNAcα1-O-Ser/Thr), sialosyl-Tn (NeuA-cα2-6Gα1NAca1-O-Ser/Thr)] were detectable by immunohistochemistry at an earlier stage than MUC1 itself (Cao et al. 1997b). In this study we examined MUC1 at the mRNA level by in situ hybridization and at the protein level by immunohistochemistry after chemical or enzymatic glycan alterations. From these data, the conclusion can be drawn that normal adult gastrointestinal epithelial cells express not only MUC1 mRNA but also the MUC1 protein. This has been established by MUC1 peptide-specific MAbs, among them the well-known MUC1 antibodies HMFG-2 and SM3. It is shown for the first time that MUC1 mRNA and the MUC1 peptide are localized at the surface of mucous cells and gastric gland cells of the stomach and in both types of epithelial cells of the intestine, columnar and goblet cells. Access of most MAbs to peptide regions of MUC1 in normal adult intestinal epithelial cells is hindered by tissue-specific glycosylation. This was established by indirect and direct evidence. The effect of periodate oxidation (Spiro 1966; Woodward et al. 1985) is obvious and specific, although not fully explained. What is certain is that the rigid ring structures of saccharides are destroyed by the procedure. This may result in a greater flexibility of the mucin molecule and hence in a better access of MAbs to the peptide backbone. Our indirect evidence consisted of the highly significant and reproducible appearance of MUC1-specific peptide epitopes after carbohydrate-restricted periodate oxidation performed on tissue sections before immunostaining. Direct evidence was obtained by pretreatment of the sections with glycosidases. In the latter case, it appeared that sialic acid and fucose alone did not play a major role in epitope masking, despite the fact that the normal gastrointestinal mucosa carries a large quantity of sialic acid (Cao et al. 1996). This differs from the results obtained by Bara et al. (1993), who found that anti-MUC1 MAbs immunoreact strongly with gastric epithelium of non-secretors, whereas tissues of secretors reacted only after treatment with α-fucosidase. Therefore, gastric and colorectal MUC1 may differ in their glycosylation and, accordingly, in their staining behavior.
Localization of partially deglycosylated MUC1 (observed after periodate oxidation) was found, as expected for a membrane-bound mucin, at the the apical membrane and in the cytoplasm of surface mucous cells of the stomach and gastric gland cells, and at the apical membrane and in the supranuclear or perinuclear cytoplasm of goblet cells and columnar cells of the intestine; it was absent in secretions. Staining patterns were different between A76-A/C7 and HMFG-2 or SM3. A76-A/C7 reacted primarily with the apical membrane of epithelial cells, whereas HMFG-2 and SM3 stained the apical membrane and the supranuclear cytoplasm. This is interpreted as indicating that A76-A/C7 recognizes a later stage in MUC1 processing, whereas HMFG-2 and SM3 may bind to an earlier stage (in terms of glycosylation or conformation). Such differences have also been frequently observed in immunofluorescence experiments on tumor cells in vitro with other MUC1-specific MAbs (Cao et al. 1997a).
Compared to most other epithelial tissues, such as mammary gland, salivary gland, esophageal epithelium, pancreas, bile ducts, lung epithelium, distal tubules and collecting ducts of kidney, urothelium, uterus, and rete testis, MUC1 of gastrointestinal epithelial cells appears to be more heavily glycosylated and its peptide backbone therefore not accessible to MAbs such as HMFG-2 or A76-A/C7. One consequence of heavy O-glycosylation is the relative resistance of these regions in glycoproteins towards proteases (Kozarsky et al. 1988; Jentoft 1990). Therefore, the more heavily glycosylated MUC1 molecules of intestinal epithelia may be instrumental in protecting the epithelial cells from the overwhelming presence of proteases in the gastrointestinal tract.
The immunohistological distribution of MUC3 in epithelial cells of the gastrointestinal tract largely confirmed earlier reports (Audie et al. 1993; Ho et al. 1993; Chang et al. 1994; Apostolopoulos et al. 1995). We found that native MUC3 (whose staining is observed without periodate oxidation) was located in the supranuclear cytoplasm of epithelial cells of the gastrointestine, whereas the more heavily deglycosylated MUC3 was seen mainly at the surface of mucous cells of the stomach, in gastric glands, at vacuoles of goblet cells of the intestine, and in secretions of the gastrointestinal tract. Masking of MUC3 by sialic acid or fucose was of minor importance at these sites.
Alterations of expression of MUC1 and MUC3 in colorectal carcinomas and adenomas of various stages have been studied by several groups (Ho et al. 1993; Chang et al. 1994). In our study, the stainability of native and partially deglycosylated mucins was compared in various colorectal tissues from normal epithelium through adenomas to carcinomas. The following results were obtained (Figure 5).
In normal colonic epithelia, MUC1 is heavily glycosylated, preventing its immunostaining with anti-peptide MUC1-specific MAbs. During tumorigenesis, the apparent expression of native MUC1 showed a gradual increase from mildly through moderately to severely dysplastic adenomas. Periodate treatment greatly enhanced the reactivity of anti-MUC1 MAbs with lesions of a lower grade of dysplasia, but less so with lesions of a higher grade of dysplasia. This reflects the fact that lesions of a higher grade of dysplasia express MUC1 with a lower degree of glycosylation. In malignant lesions, native MUC1 was strongly expressed, and periodate treatment only slightly enhanced the staining. This indicates that, similar to MUC1 in highly dysplastic adenomas, MUC1 in carcinomas is less glycosylated, resulting in exposed peptide epitopes that can be recognized by their corresponding antibodies. It is obvious that the grade of dysplasia of adenomas is closely related to their size. Adenomas of larger size, of a higher grade of dysplasia, or of the villous type have a higher malignant potential (Konishi and Morson 1982). The low glycosylation of MUC1 and reduced degree of glycosylation may be associated with the malignant potential and status of lesions in the colon. Several defects in glycosyltransferase activities in colorectal cancer have been described (King et al. 1994; Vavasseur et al. 1994; Yang et al. 1994), which may explain the observed changes in MUC1 glycosylation. In some carcinomas, especially mucinous carcinomas and poorly differentiated carcinomas, even periodate oxidation did not result in positive immunohistochemical staining with MUC1-specific MAbs. These carcinomas apparently have lost the ability to express MUC1. Periodate oxidation therefore provides a simple method to distinguish between lack of expression and masking of epitopes by glycans. The secretions of some tumors are positive for MUC1, which is unusual because normal gastrointestinal secretions do not contain this mucin. MUC1 in tumor secretions may originate from shedding or destruction of tumor cells. MUC3 revealed a similar change in staining behavior (increased staining without pretreatment) during malignant transformation. In most adenocarcinomas, MUC3 expression was decreased compared with normal colon and benign lesions owing to downregulated synthesis (Weiss et al. 1996). Whereas normal epithelial cells and benign lesions show the typical apical distribution of MUC1 and MUC3 in addition to some supranuclear staining, malignant lesions and “benign” lesions with very high malignancy risk reveal apolar distribution of both mucins. This indicates abnormal intracellular transport in addition to incomplete glycosylation in malignant lesions. Finally, the expression of mucins within a given tumor is often heterogeneous, from zero to very high amounts per cell.
Normal breast and mammary carcinomas have been examined for the expression of MUC1 and MUC3 in a number of reports (Zotter et al. 1988; Xing et al. 1994). In this study, changes in immunostaining in normal breast and mammary carcinomas after partial deglycosylation have been investigated. Normal mammary epithelial cells were found positive for A76-A/C7 and HMFG-2 but negative for SM3, in accord with previous reports (Burchell et al. 1987). This difference has been explained by the more extended SM3 epitope compared to the HMFG-2 epitope, which makes it more dependent on the influence of glycosylation (Burchell et al. 1989). We demonstrate here that, after periodate treatment, identical staining results were obtained with A76-A/C7, HMFG-2, and SM3. Neuraminidase or α-fucosidase treatment did not expose the SM3 epitope, indicating that sialic acid and fucose do not play major roles in epitope masking in breast epithelium.
For carcinomas of the breast, some general aspects of MUC1 immunoreactivity were found similar to that of colorectal carcinomas, e.g., decreased epitope masking compared to normal epithelial cells, disturbed polar distribution in cancer cells, and heterogeneous reactivity within a given tumor. In addition, some features of apomucin expression specific for mammary carcinomas were observed. The reactivity of MUC1 MAbs with mammary carcinomas could be enhanced by prior neuraminidase treatment to a greater extent than in the case of colorectal carcinomas. This is in accord with a report by Ho et al. (1995). In another study by us, all mammary carcinomas examined were found to become positive for the MUC1-associated, normally masked Thomsen–Friedenreich glycotope after neuraminidase treatment (unpublished data), whereas in colorectal carcinomas only 37 of 52 cases (71%) responded in this manner (Cao et al. 1995). We interpret this as indicating that in the gastrointestinal tract sialic acids contribute less to epitope masking than in the breast. In general, mammary carcinoma cells showed stronger staining of MUC1 than normal cells before or after partial deglycosylation. This may be the result of an upregulated mucin synthesis (Hilkens et al. 1995) and/or of mucin accumulation owing to abnormal transport within the tumor cells. A minority (7/20) of mammary carcinomas express ectopic MUC3. Its absence in normal epithelium was demonstrated in a previous report (Ho et al. 1993) and is confirmed in this study. Therefore, it appears that MUC3 is a neoantigen in breast cancer. We also noted that partial deglycosylation slightly enhanced its staining intensity.
The observed changes of mucin expression and its glycosylation in colorectal and mammary carcinomas may be of biological and clinical significance. For example, the exposed glycan core structures on the surface of cancer cells may lead to or enhance binding to the basal membrane and extracellular matrix (Schwartz et al. 1992), to hepatocytes (Cao et al. 1995), and to endothelial cells (Regimbald et al. 1996). Enhanced mucin expression with apolar distribution strongly reduces the contacts between epithelial cells (Hilkens et al. 1992) and may protect tumor cells from the immune system (Gimmi et al. 1996). Such changes may participate in tumor progression. On the other hand, the less glycosylated, “tumor-specific” mucins in tissues, blood, and secretions may serve as neoplastic markers for clinical diagnosis and as targets for immunotherapy.
In conclusion, we have presented evidence that normal gastrointestinal epithelia express both MUC1 mRNA and the protein itself. The immunohistochemical detection of the MUC1 protein is hampered by a qualitatively different type of glycosylation compared to other secretory epithelia (e.g., breast). This can be overcome by carbohydrate-specific pretreatment with periodate, which not only leads to the detection of masked MUC1 but also allows evaluation of the changes in MUC1 glycosylation that occur in neoplastic lesions. During malignant transformation, epithelial mucins experience dramatic (yet tissue-specific) alterations in their glycosylation and cellular localization, and in some cases show ectopic expression. These changes may be significant in tumor progression, and possibly also in the clinical setting. Periodate oxidation as a complementary measure in immunohistochemistry may help us to examine and understand these changes.
Footnotes
Acknowledgements
Supported by Deutsche Forschungsgemeinschaft grant no. Ka 921/1.
We thank Dr D. Swallow (London) for providing the cDNA probe, and especially acknowledge the help of Ms Andrea Schaffrath (Bremen) with the in situ hybridization. Drs G. Pilgrim and W. Fregin are thanked for their help in providing blood group and secretor data for breast cancer patients.