
Abstract
Two distinct chitinases have been identified in mammals: a phagocyte-specific enzyme named chitotriosidase and an acidic mammalian chitinase (AMCase) expressed in the lungs and gastrointestinal tract. Increased expression of both chitinases has been observed in different pathological conditions: chitotriosidase in lysosomal lipid storage disorders like Gaucher disease and AMCase in asthmatic lung disease. Recently, it was reported that AMCase activity is involved in the pathogenesis of asthma in an induced mouse model. Inhibition of chitinase activity was found to alleviate the inflammation-driven pathology. We studied the tissue-specific expression of both chitinases in mice and compared it to the situation in man. In both species AMCase is expressed in alveolar macrophages and in the gastrointestinal tract. In mice, chitotriosidase is expressed only in the gastrointestinal tract, the tongue, fore-stomach, and Paneth cells in the small intestine, whereas in man the enzyme is expressed exclusively by professional phagocytes. This species difference seems to be mediated by distinct promoter usage. In conclusion, the pattern of expression of chitinases in the lung differs between mouse and man. The implications for the development of anti-asthma drugs with chitinases as targets are discussed.
Keywords
The physiological function of chitotriosidase is not completely resolved. Its phagocyte-specific expression suggests a role in innate immunity. Highly homologous plant chitinases are prominent “pathogenesis-related proteins” that are induced following attack by pathogens and take part in the defense against chitin-containing fungi (Schlumbaum et al. 1986; Sahai and Manocha 1993). A similar role for chitotriosidase in the human innate immune system is indicated by our observation that recombinant chitotriosidase is fungistatic in mice models of systemic fungal infections (Stevens et al. 2000; van Eijk MC, Boot RG, and Aerts JMFG, unpublished data). Interestingly, ≃1 in 20 individuals is completely deficient in chitotriosidase activity due to a 24-bp duplication in the gene that occurs panethnically (Hollak et al. 1994; Boot et al. 1998). This high prevalence of deficiency suggests that human chitotriosidase either no longer fulfills an important function under normal conditions or that other mechanisms may compensate for the lack of a functional chitinase (Boot et al. 1998). The occurrence of deficiency in chitotriosidase is associated with susceptibility to infection with Wuchereria bancrofti, a filarial parasite whose microfilarial sheath contains chitin (Choi et al. 2001). In search of compensatory chitinases in mammals we identified and characterized a second mammalian chitinase named acidic mammalian chitinase (AMCase) (Boot et al. 2001). This 50-kDa enzyme is structurally highly related to chitotriosidase.
AMCase occurs in the gastrointestinal tract and lungs of rodents and man. Suzuki et al. (2002) also detected AMCase mRNA in exocrine cells of the serous type of mice. Unlike human chitotriosidase, AMCase has an acidic pH optimum and is very acid stable (Boot et al. 2001). The enzyme appears to be adapted to function in the extreme stomach environment, where it may fulfill a role in defense and/or digestion of chitin-containing organisms.
In the lung of mice, but not of man, AMCase is the sole detectable endogenous chitinase. In a number of recent reports the mRNA expression of AMCase in the lung of mice was shown to be highly regulated (Sandler et al. 2003; Xu et al. 2003; Zhu et al. 2004; Zimmermann et al. 2004). Intravenous injection of Schistosoma mansoni eggs was found to cause massive expression of AMCase in the lung of wild-type mice and animals with an exaggerated Th2 response that is dominated by the cytokines IL4 and IL13. This induction did not occur in mice with an exaggerated Th1 response or IL13-knockout mice (Sandler et al. 2003). Zimmermann et al. (2004) reported highly induced AMCase mRNA levels in mouse models of experimental asthma induced either by ovalbumin or by Aspergillus fumigatus antigen. This induction was mediated by the STAT6 signaling pathway, again suggesting a role for IL4 or IL13 (Zimmermann et al. 2004). Very recently it was shown for an aeroallergen asthma mouse model that AMCase is induced in the lung via a Th2-specific, IL13-mediated pathway (Zhu et al. 2004). Interestingly, AMCase activity appeared instrumental for the pathogenesis of asthma. Inhibition of AMCase, either by a specific antibody or the specific chitinase inhibitor allosamidin, alleviated the Th2-mediated inflammatory damage that occurs in asthma (Zhu et al. 2004). It has been suggested that inhibition of chitinase activity may render an attractive new therapy for asthma (Couzin 2004; Zhu et al. 2004).
So far, all attention in studies with mouse models has been focused on AMCase, whereas chitotriosidase, the dominant chitinase in man, has received little attention. We investigated the expression of chitotriosidase and AMCase in mice in more detail. We show here that remarkable differences between man and mouse exist regarding cell type and tissue-specific expression of chitinases. Comparison of promoter sequences of the human and murine chitinase genes helps to explain the species-specific tissue expression of chitinases. The implications for extrapolating observations on chitinase made in mouse models to the human situation are discussed.
Materials and Methods
Enzyme Assays
Chitinase enzyme activity was determined with the fluorogenic substrates 4MU-chitobiose (4-methylumbelliferyl β-D-N,N'-diacetylchitobiose; Sigma, St Louis, MO) and 4MU-chitotriose (4-methylumbelliferyl β-D-N,N',N”-triacetylchitotriose; Sigma). Assay mixtures contained 0.027 mM substrate and 1 mg/ml of bovine serum albumin in McIlvaine buffer (100 mM citric acid, 200 mM sodium phosphate) at the indicated pH. The standard enzyme activity assay for human chitotriosidase with 4MU-chitotriose substrate was performed at pH 5.2, as previously described (Hollak et al. 1994). The standard AMCase enzyme activity assays with 4MU-chitobiose substrate were performed at pH 4.5.
RNA Isolation, Northern Blot, and RNA Master Blot Analysis
Total spleen RNA was isolated using the RNAzol B (Biosolve; Barneveld, The Netherlands) RNA isolation kit according to the manufacturer's instructions. For Northern blot analysis, 15-μg samples of total RNA were run in 10 mM Hepes (pH 7.5), 6% formaldehyde-agarose gels, transferred to Hybond N nylon membranes (Amersham; Buckinghamshire, UK) by the capillary method, and immobilized by UV cross-linking. Full-length mouse chitotriosidase cDNA was used as probe. Human RNA Master Blots (Clontech; Palo Alto, CA) were used to examine the tissue distribution of the human chitotriosidase transcript according to the instructions of the manufacturer, using the full-length human chitotriosidase cDNA as probe. The probes were radiolabeled with 32P using the random priming method. Hybridization conditions were exactly as previously described (Boot et al. 1995).
Isoelectric Focusing
The native isoelectric point of chitinases was determined by flatbed isoelectric focusing in granulated Ultrodex gels (Pharmacia; Uppsala, Sweden) as described (Renkema et al. 1995).
cDNA Cloning of the Mouse Chitotriosidase
Reverse transcription-polymerase chain reaction (RT-PCR) fragments were generated from mouse pooled tissue total RNA using degenerate oligonucleotides, as described (Boot et al. 1995). Obtained fragments were cloned in pGEM-T (Promega; Madison, WI) and sequenced. A comparison with the GenBank mouse expressed sequence tag (EST) database using the basic local alignment search tool (BLAST) at the National Center for Biotechnology Information showed that several EST clones matched the mouse chitotriosidase cDNA sequence. Full-length mouse chitotriosidase cDNA was generated using specific primers based on these deposited sequences. The nucleotide sequence of two independent clones from the PCR were sequenced from both strands by the procedure of Sanger using fluorescent nucleotides on an Applied Biosystems (ABI; Foster City, CA) 377A automated DNA sequencer following ABI protocols.
Transient Expression in COS-1 Cells
Transient expression of the various cDNAs in COS-1 cells was performed to generate recombinant enzyme exactly as described previously (Boot et al. 1995).
Tissue Processing
More detailed practical protocols for fixation, paraffin embedding, mounting, and sectioning have been described (Moorman et al. 2000). In short, tissues removed from a FVB mouse were fixed for 4 hr to overnight in freshly prepared 4% formaldehyde in PBS by rocking at 4C. The tissues were then dehydrated in a graded ethanol series, paraffin embedded, cut into sections, and carefully mounted on aminoalkylsilane-coated slides to prevent loss of the tissue sections during the extensive treatments of the in situ hybridization (ISH) procedure.
RNA Probes and Probe Specification
Digoxigenin-labeled probes were made according to the manufacturer's specifications (Roche; Mannheim, Germany). RNA probes complementary to the full-length mouse mRNAs encoding chitotriosidase, AMCase, GOB-5 or calcium-activated chloride channel 3, glutamine synthase, and lysozyme P were used. Due to the high identity, the lysozyme probe under the conditions used also detects the lysozyme M mRNA.
Non-radioactive ISH
Non-radioactive ISH was performed as described (Moorman et al. 2001). In short, after removal of paraffin, sections were pretreated by proteolytic digestion for 5–15 min at 37C with 20 μg/ml proteinase K dissolved in PBS, followed by a 5-min rinse in 0.2% glycine/PBS, and two rinses of 5 min in PBS. Sections were then re-fixed for 20 min in 4% formaldehyde/0.2% glutaraldehyde dissolved in PBS to ensure firm attachment of the sections to the microscope slides and washed twice in PBS for 5 min. Sections were prehybridized in hybridization mix without probe for 1 hr at 70C and then hybridized overnight at 70C. The hybridization mixture was composed of 50% formamide, 5 X SSC, 1% block solution (Roche), 5 mM EDTA, 0.1% Tween-20, 0.1% Chaps (Sigma), 0.1 mg/ml heparin (Becton-Dickinson; Mountain View, CA), and 1 mg/ml yeast total RNA (Roche). Probe concentration was ≃1 ng/μl. Approximately 6 μl hybridization mix was applied to the sections, and no coverslips were used. After hybridization, sections were rinsed in 2X SSC, pH 4.5, washed three times for 30 min at 65C in 50% formamide/2X SSC, pH 4.5, followed by three 5-min washes in PBST. Probe bound to the section was immunologically detected using sheep anti-digoxigenin Fab fragment covalently coupled to alkaline phosphatase and NBT/BCIP as chromogenic substrate, essentially according to the manufacturer's protocol (Roche). Sections were washed with double-distilled water, dehydrated in a graded ethanol series and xylene, and embedded in Entellan (Merck; Darmstadt, Germany).
Results
The mouse and human ortholog of the AMCase have been identified recently (Boot et al. 2001). The human AMCase gene is located on chromosome 1p13, whereas the locus of the human chitotriosidase gene is found on chromosome 1q32 (Boot et al. 1998). The mouse AMCase gene is located on chromosome 3F3, a region that is syntenic with human 1p13. Recently, we and others have cloned the mouse ortholog of human chitotriosidase (GenBank Accession numbers: AY536287.1 and AY458654.1, respectively).
Alignment of mouse and human chitotriosidase amino acid sequences revealed that there is an identity of 75% and a similarity of 78%. To group all the known mouse and human chitinase protein family members and to determine whether this cloned sequence is the true ortholog of human chitotriosidase, multiple sequence alignments of their cDNAs, coding for the catalytic 39-kDa domain only, were made using the Clustal X program (Thompson et al. 1997). The sequences used for this alignment are human chitotriosidase (GenBank Accession Number: U29615), human-HC gp39 (GenBank Accession Number: M80927), human-AMCase (GenBank Accession Number: AF290004), human-oviductin (GenBank Accession Number: U09550), human-YKL39 (GenBank Accession Number: U49835), mouse-BRP39 (GenBank Accession Number: X93035), mouse-YM1 (GenBank Accession Number: M94584), mouse-AMCase (GenBank Accession Number: AF290003), and mouse-oviductin (GenBank Accession Number: D32137). Grouping of these members is calculated by the method of Neighbor Joining by Saitou and Nei (Saitou and Nei 1987). This analysis showed that the cloned mouse sequence is grouped together with human chitotriosidase. It suggests that the mouse sequence is more homologous to human chitotriosidase than to any other member of the chitinase protein family, which is indicative of being orthologs of each other.
Using the Mouse Genome Server from the Mouse Genome Sequencing Consortium (at http://www.ensembl.org/Mus_musculus), we identified genomic sequences that contained the chitotriosidase gene, enabling the determination of intron-exon boundaries. The boundaries are all conserved between human and mouse chitotriosidase genes (data not shown). Moreover, it was found that the mouse chitotriosidase gene is located on chromosome 1 band E4 near the boundary with F. This region is syntenic with human chromosome 1q32. On human 1q32, only two genes of members of the chitinase protein family are located, namely, chitotriosidase (CHIT1) and HC gp39 (CHI3L1) (Jin et al. 1998). On mouse chromosome 1 band E4 the mouse BRP39 (Chi3l1), the mouse ortholog of HC gp39, was already identified (Jin et al. 1998). The data present on the Mouse Genome Server show that, in addition to the BRP39 gene, the mouse chitotriosidase gene is also found at this locus (see Figure 1 for overview). In addition to the syntenic chromosomal locations, the homology in intronic sequences of the mouse and human chitotriosidase genes indicate that they are orthologs.
Comparison of the properties of human and mouse chitotriosidases revealed that both enzymes show a pH optimum that is relatively broad and peaks around pH 6 (Figure 2A). Like human chitotriosidase, the mouse enzyme is not acid stable in sharp contrast to the mouse AMCase, which is extremely acid stable (Figure 2B).
Northern blot analysis of different mouse tissues with the mouse chitotriosidase cDNA probe revealed highest expression in tongue and slightly less in the stomach. In other tissues, expression was not detectable (Figure 3A). The expression pattern of human chitotriosidase is remarkably different. Human chitotriosidase is expressed at relatively high levels in lymph node, bone marrow, and lung as determined with commercial tissue dot blot (Figure 3B). No expression of chitotriosidase was detectable in the human stomach. Human AMCase was found to be relatively highly expressed in the stomach and to a lesser extent in the human lung using the same commercial RNA Master Blot (Boot et al. 2001). Moreover, previously it was also observed by Northern blot analysis that AMCase is expressed predominantly in the stomach, salivary glands, and lungs of mice (Boot et al. 2001).
Non-radioactive ISH was used to examine more closely the cellular sources of mouse chitotriosidase and mouse AMCase. As references, the expression of other genes was visualized. The genes used as reference are lysozyme P because of the analogy with chitinases, GOB-5, a calcium-activated chloride channel and goblet cell marker, and glutamine synthase as a positive control for expression in the stomach and liver.
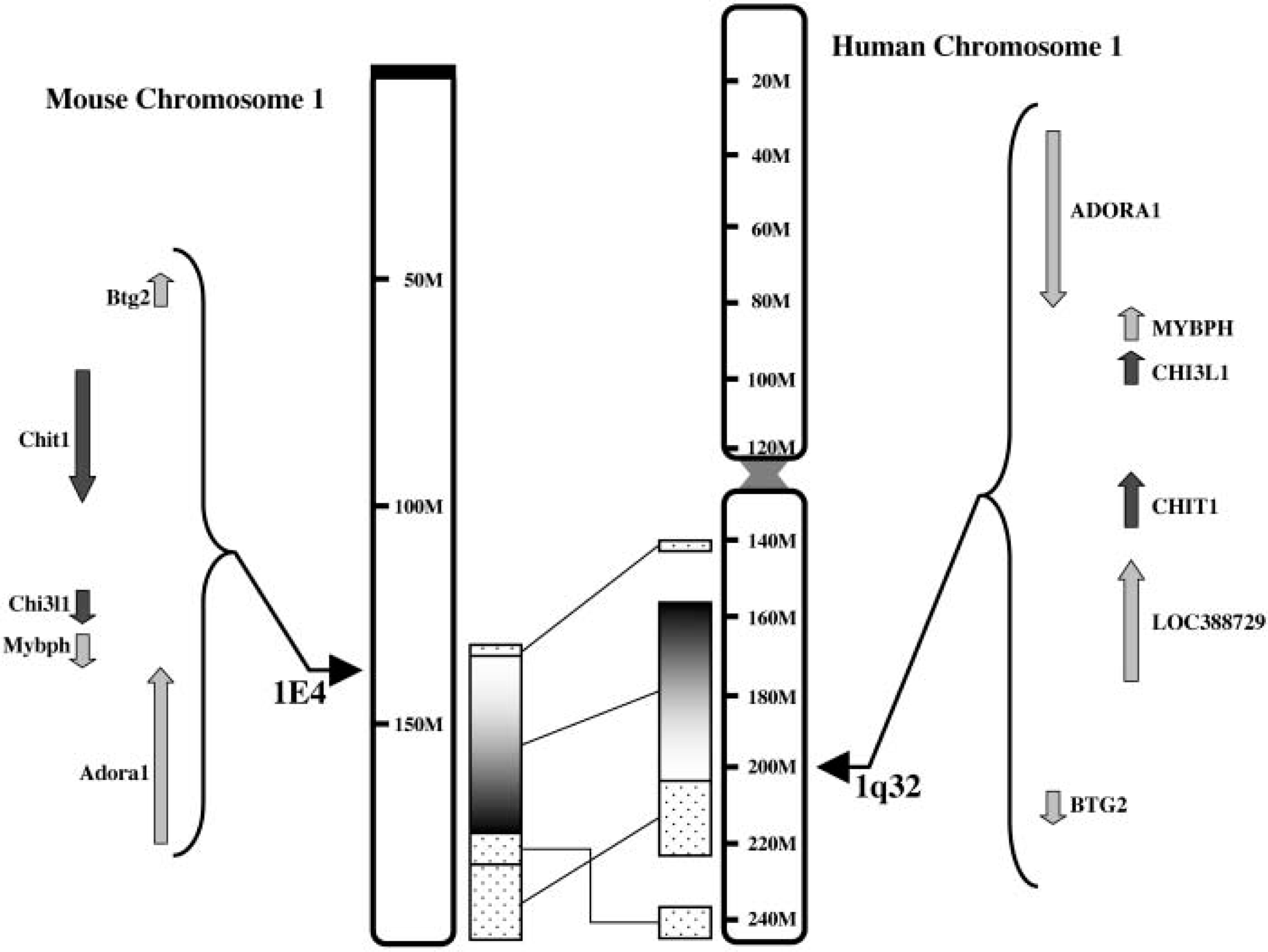
Schematic overview of the synteny of mouse locus 1E4 with human 1q32. On the left-hand side, mouse chromosome 1 with a part of locus 1E4 is indicated. On the right-hand side, the human syntenic region 1q32 is shown. The orientation and position of a few genes in the direct neighborhood of the mouse and human chitotriosidase gene are depicted. The genes of members of the chitinase protein family are depicted in dark gray arrows, whereas the other genes are indicated with light gray arrows. The genes are BTG2, Btg2: B-cell translocation gene 2; CHIT1, Chit1: chitotriosidase; CHI3L1, Chi3l1: cartilage glycoprotein 39; MYBPH, Mybph: myosin-binding protein H; ADORA1, Adora1: adenosine A1 receptor; LOC388729: gene coding for the human hypothetical protein with GenBank Accession Number: XP373882.
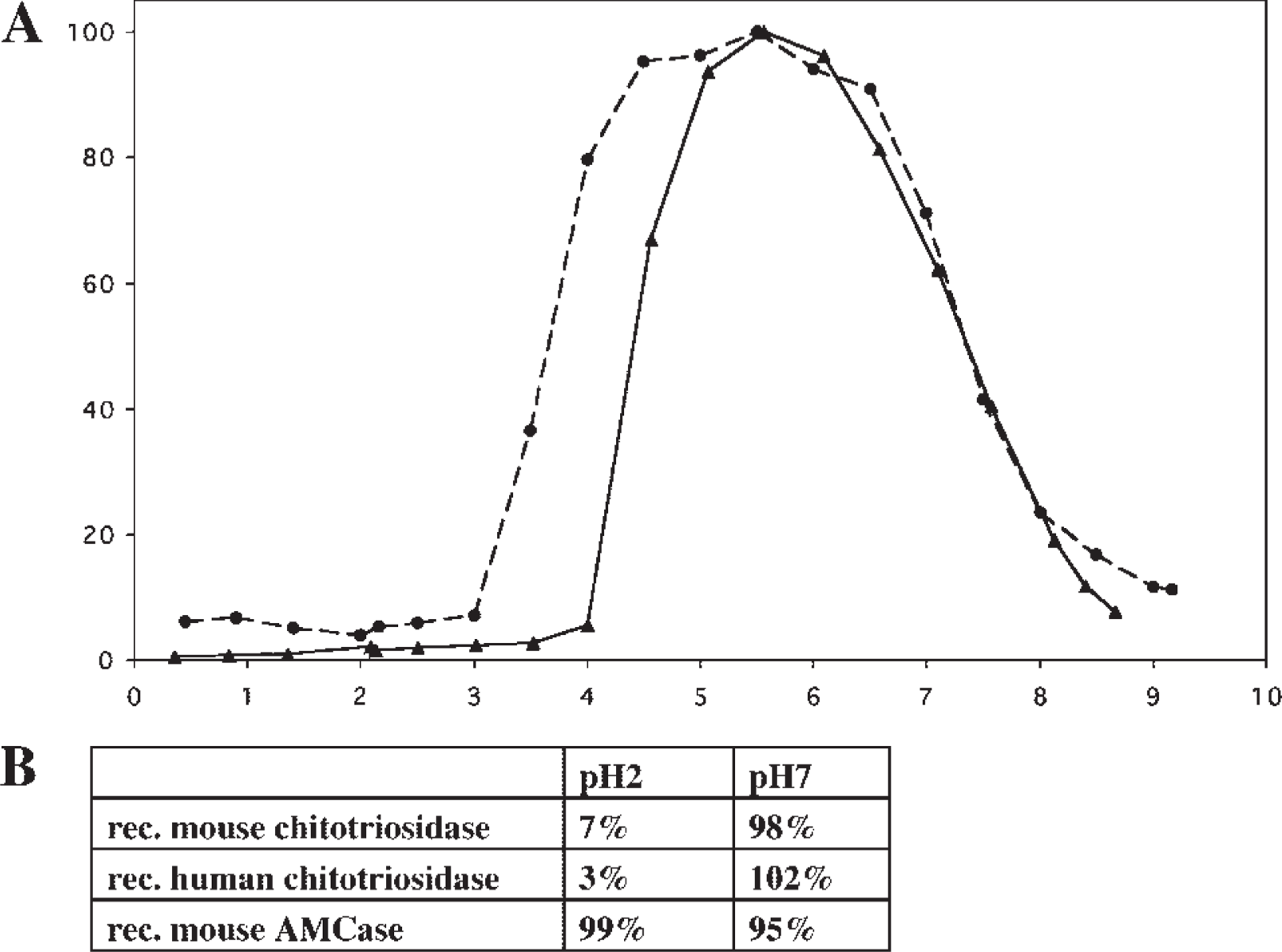
pH activity profile of the different recombinant chitotriosidases. The pH optima were determined by monitoring enzyme activity at the indicated pH in McIlvaine buffer. Purified human recombinant chitotriosidase (closed circle), purified recombinant mouse chitotriosidase (closed triangle).
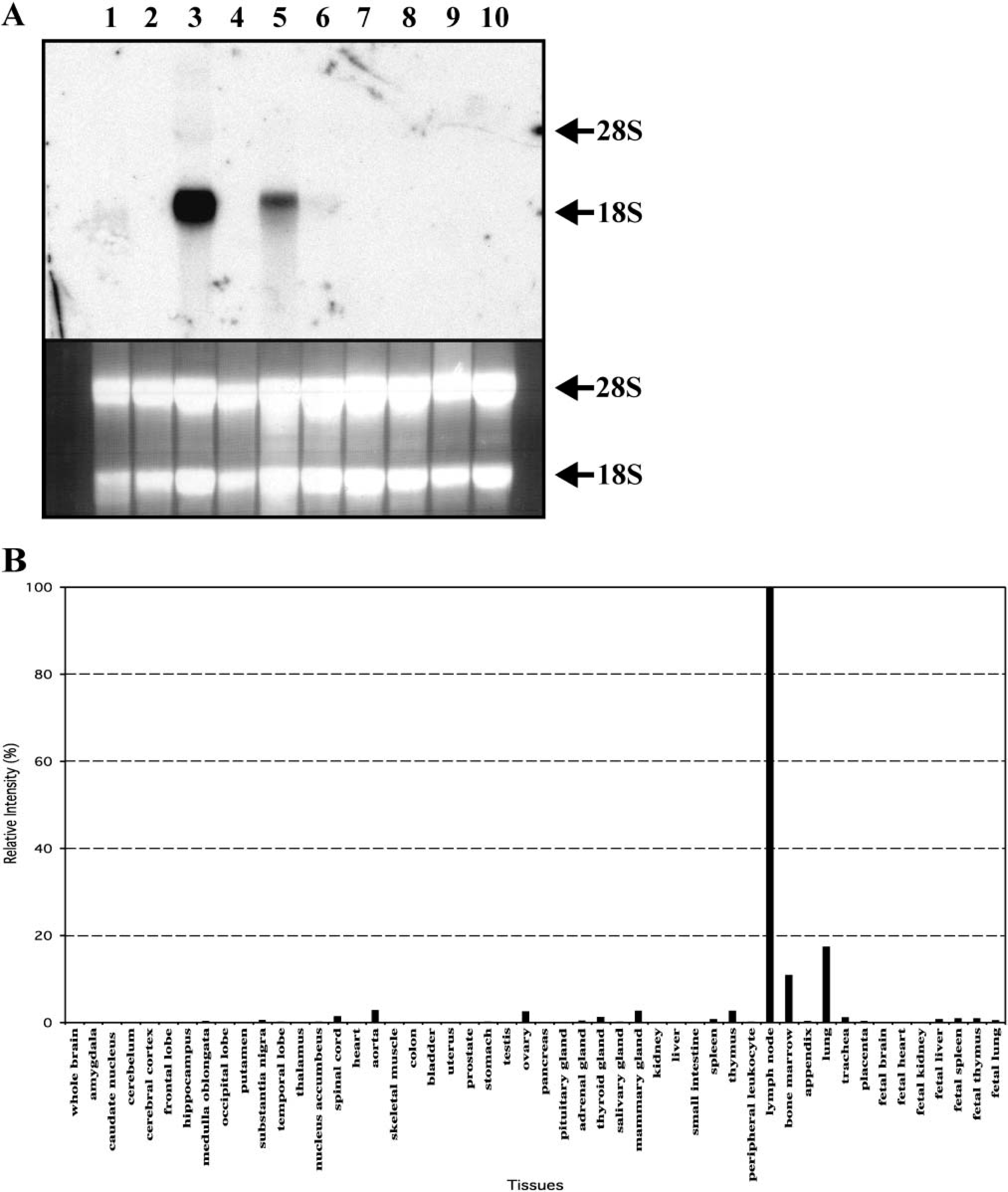
Tissue distribution of mouse chitotriosidase mRNA.
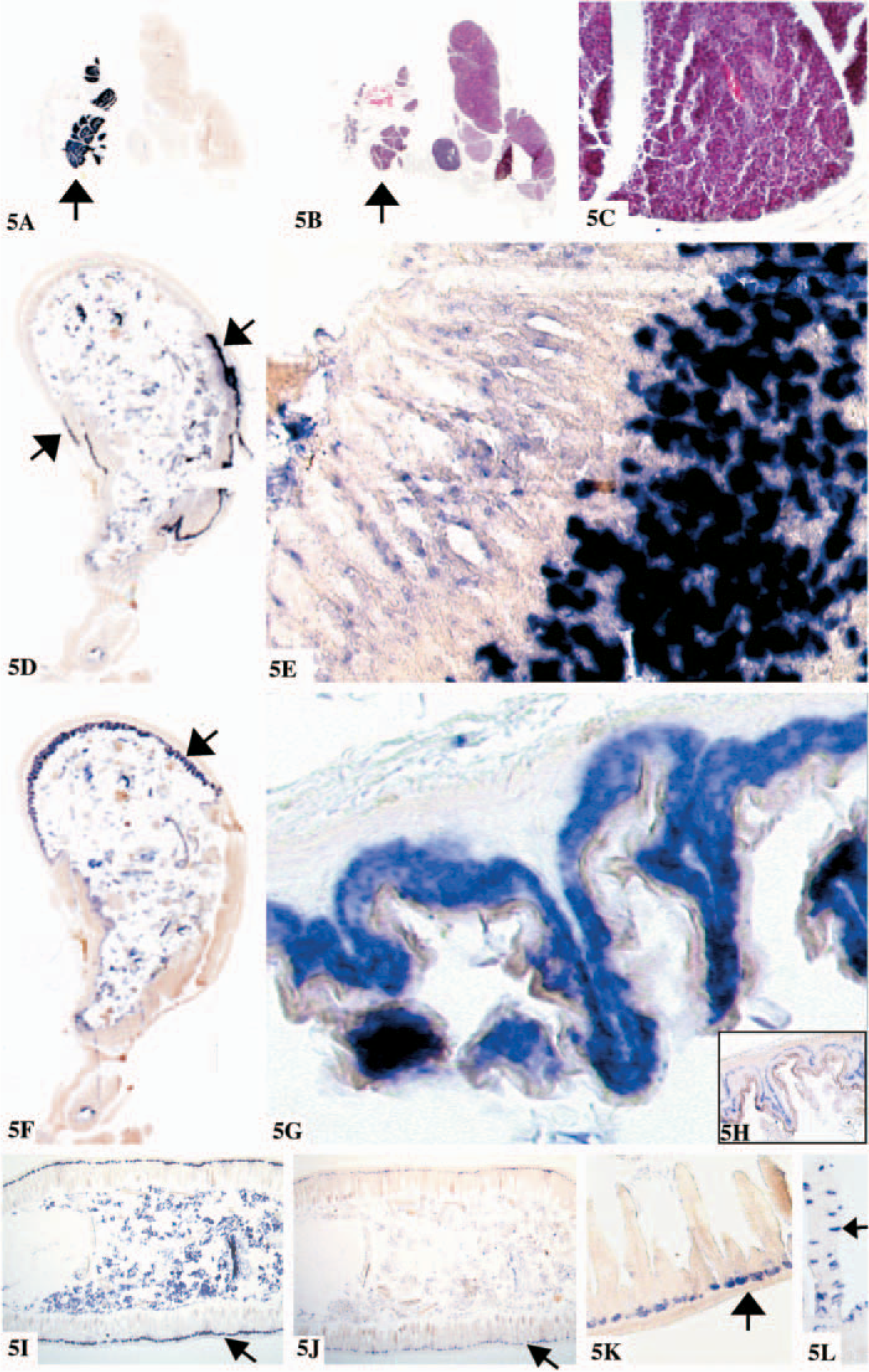
Using ISH, no mRNA for chitotriosidase and AMCase was detectable in brain, colon, pancreas, liver, heart, kidney, skin, and spleen (data not shown) in agreement with the results obtained with the Northern blot analysis. The expression of chitinases in the gastrointestinal tract of mice is complex. Starting in the mouth, profound expression of both chitinases is observed. Chitotriosidase expression was mainly detected in the mucosal surface of the tongue in the stratified squamous epithelium of the papilla (Figures 4A-4E). Whereas AMCase is mainly found in the specialized minor lingual salivary glands of the serous type, the so-called glands of von Ebner (Figures 4F and 4G), in agreement with the results described (Suzuki et al. 2002; Goto et al. 2003), in the major salivary glands intense expression of AMCase was observed in the parotid gland (Figures 5A-5C), and no expression of chitotriosidase was noted (data not shown).
Moreover, unlike the situation in humans, both chitinases are highly expressed in the mouse stomach (Figures 5D-5H). However, chitotriosidase expression is restricted to the non-glandular fore-stomach, whereas mRNA is found in the stratified squamous epithelial layer. Glutamine synthase mRNA is also found in this region of the stomach, in a cell layer just beneath that of the chitotriosidase-expressing cells (Figures 5G and 5H). In agreement with the results of Suzuki et al. (2002) and Goto et al. (2003), we found that AMCase is expressed in the glandular portion of the stomach, at the bottom of the gastric glands in chief cells (Figures 5D and 5E). The pyloric glands in the antrum of the stomach show no expression of the chitinases. Further down the gastrointestinal tract, in the duodenum but also in other parts of the small intestine, chitotriosidase is expressed by Paneth cells in the crypts of Lieberkühn (Figures 5I-5K). A similar expression is observed for lysozyme. We were unable to detect AMCase in the intestine.
The expression of chitinases in the lung is of particular interest. In normal human lung, chitotriosidase is prominently expressed by alveolar macrophages. In sharp contrast, we were unable to detect significant amounts of chitotriosidase mRNA in murine lung. On the other hand, AMCase mRNA is expressed by alveolar macrophages in the mouse (Figure 6).
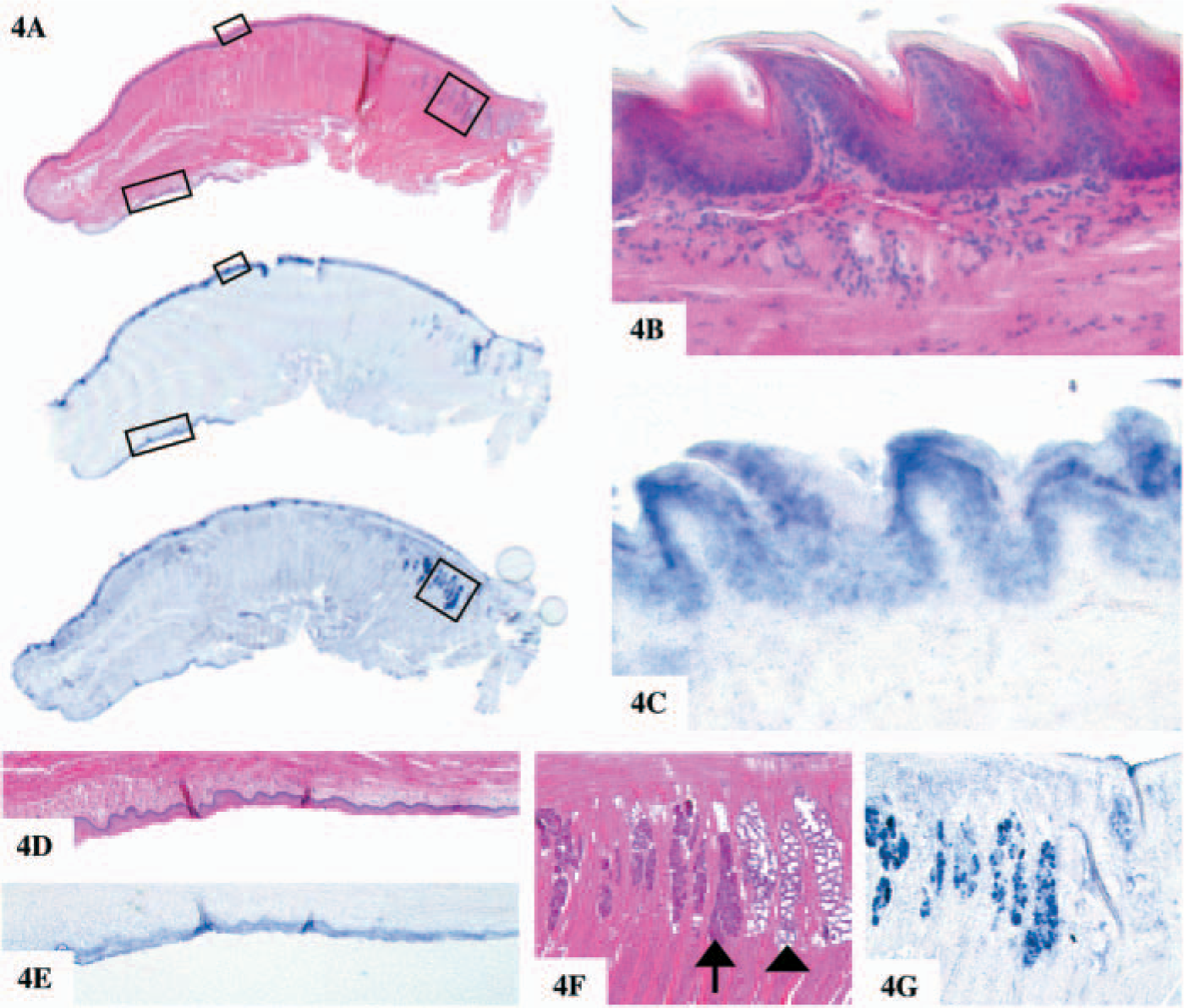
Non-radioactive in situ hybridization (ISH) on the tongue of normal mice. Non-radioactive ISH was performed on sections of the mouse tongue. Probe bound to the sections were immunologically detected using sheep anti-digoxigenin Fab fragment covalently coupled to alkaline phosphatase and NBT/BCIP as chromogenic substrate. The sections are not counterstained, and blue/black precipitates indicate specific mRNA expression.
Non-radioactive ISH on the gastrointestinal tract of normal mice. Non-radioactive ISH was performed on sections containing different tissues of the gastrointestinal tract. Probe bound to the sections were immunologically detected using sheep antidigoxigenin Fab fragment covalently coupled to alkaline phosphatase and NBT/BCIP as chromogenic substrate. The sections are not counterstained, and blue/black precipitates indicate specific mRNA expression.
To rule out mice strain-related effects as the basis for the observed differences in chitotriosidase expression, the activity of chitinases was measured in various relevant tissues (lung, liver, spleen, stomach, several parts of the intestine, kidney, salivary glands, tongue, and blood) from male FVB and C57BL/6 mice at different pHs. The relative distribution of activities was identical for both mice strains at all pHs measured, clearly indicating that the pattern of chitinase expression in these different mouse strains is preserved. However, it was noticed that enzyme levels were ≃3-fold higher in FVB tissues compared with the corresponding tissues from C57BL/6 mice.
The remarkable difference in expression of mouse and human chitotriosidase was further investigated by analysis of the promoter regions. Mouse EST cDNA sequences containing the 5’ untranslated region indicate that the mouse chitotriosidase gene has an extra exon compared with the human gene. This first exon is located at least 7000 bp upstream. The human chitotriosidase promoter is homologous to a region found in mouse intron 1 just upstream of exon 2. In conclusion, the promoter of the mouse chitotriosidase gene differs fundamentally from the human one (Figure 7). This likely explains the noted differences in cell type expression of human and mouse chitotriosidase.
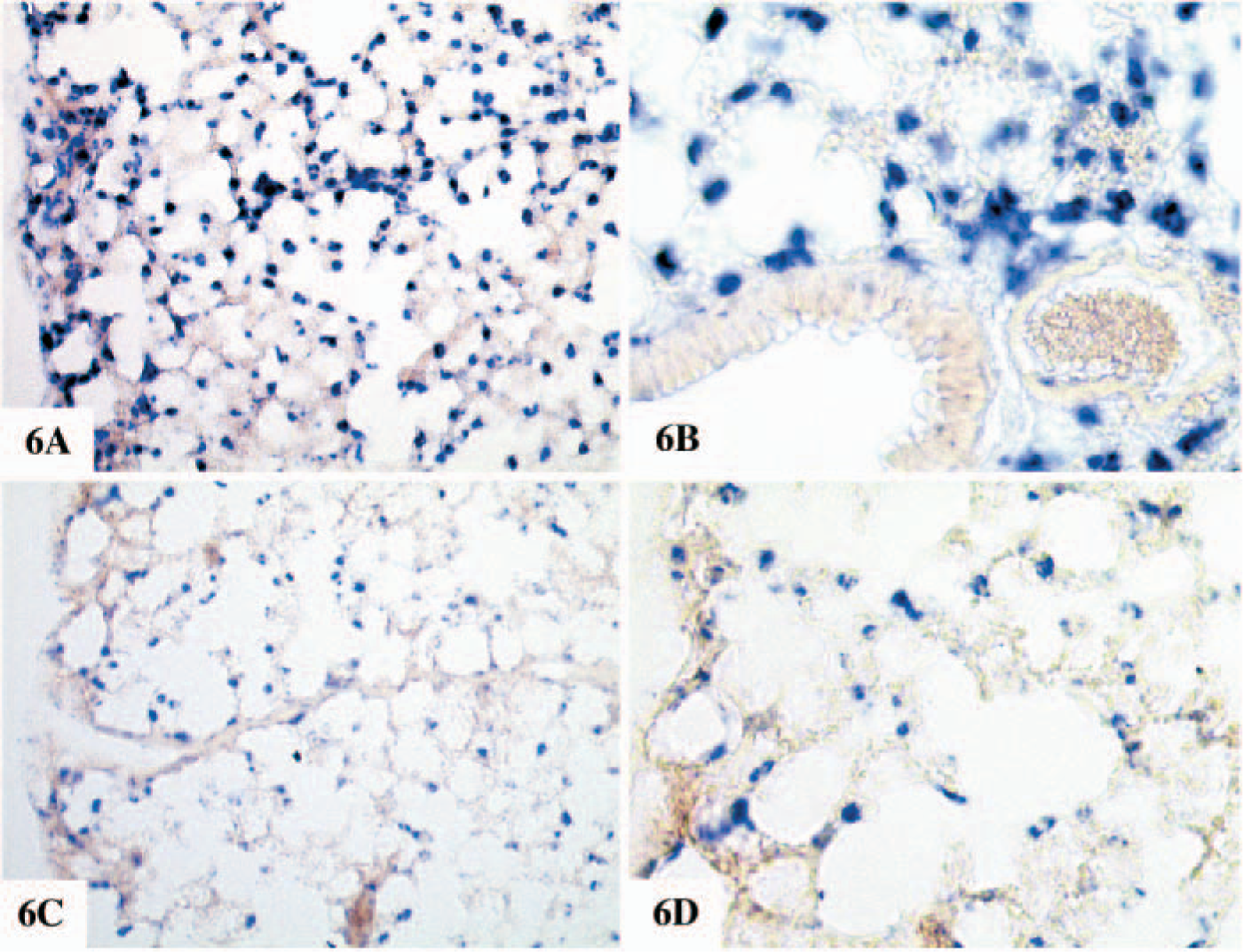
Non-radioactive ISH on mouse lung. Non-radioactive ISH was performed as described in Materials and Methods. Probe bound to the sections were immunologically detected using sheep anti-digoxigenin Fab fragment covalently coupled to alkaline phosphatase and NBT/BCIP as chromogenic substrate. The sections are not counterstained, and blue precipitates indicate specific mRNA expression. In the mouse lung, strong mRNA expression could be detected for lysozyme M
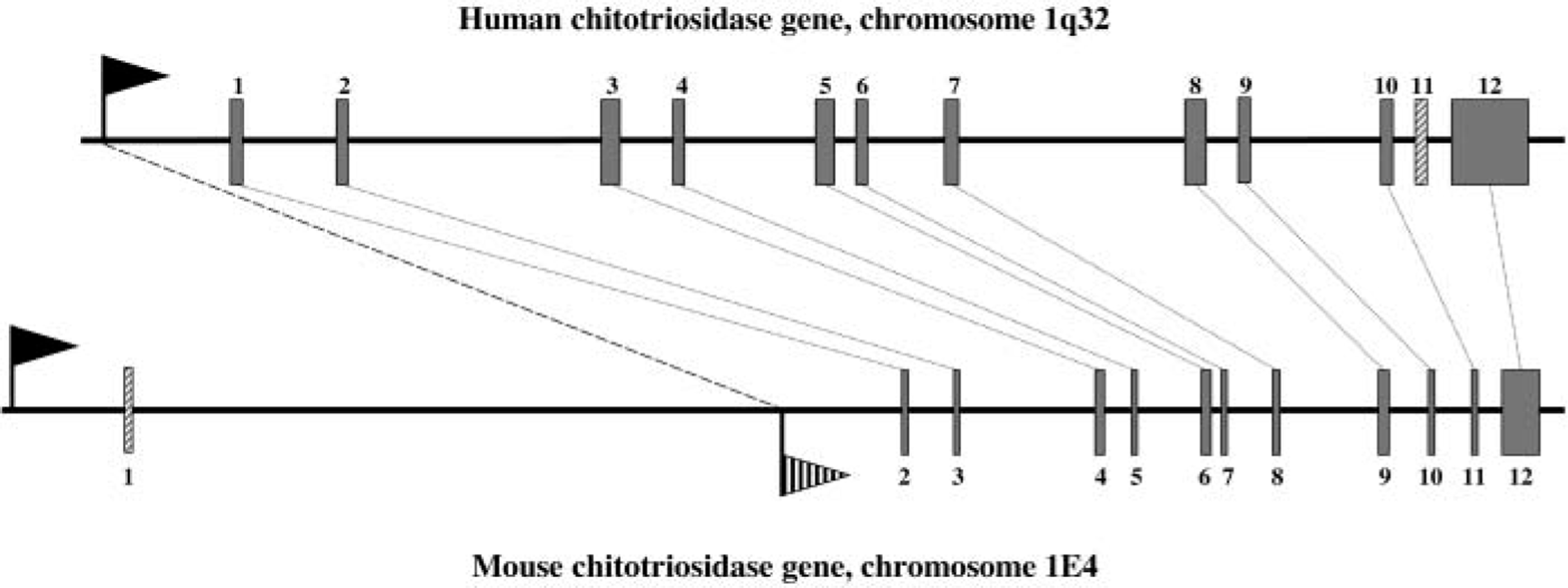
Schematic overview of mouse and human chitotriosidase gene. Homology between mouse and human chitotriosidase exons is indicated with dotted lines. Promoters are indicated with the black flags. The shaded flag indicates the region in mouse intron 1 that is homologous to the human chitotriosidase promoter.
Discussion
Striking analogies exist between mammalian chitinases and lysozymes. Both classes of enzymes are endoglucosaminidases with a compact globular structure lacking N-linked glycans. Expression of both classes of enzymes is highly regulated, being restricted to certain cell types. A dual function in defense and polysaccharide processing is envisioned for lysozymes and chitinases. For example, in many animals lysozyme functions as a bacteriolytic enzyme expressed in tissue macrophages, but during evolution of various species it has also been recruited for a nutritional function in the gastrointestinal tract (Dobson et al. 1984; Jolles et al. 1984). The expression in man of chitotriosidase in phagocytes and that of acid-stable AMCase in the gastrointestinal tract is reminiscent of this. Like lysozyme, gastrointestinal chitinases might have a dual function in defense and food processing. Research on lysozyme has provided insight into the mechanisms that allow the evolution of specialized lysozymes, driven by the need of a specific organism. For example, ruminants have duplicated the lysozyme gene to yield about ten copies, of which several are expressed in the gastrointestinal tract. Some of these enzymes have evolved into extremely acid-stable proteins with an acidic pH optimum dependent on the site of expression (Irwin 1996; Prager 1996). A very similar process seems to have taken place in the mammalian chitinase family. Duplications have occurred, ultimately leading to specialized chitinases. In the gastrointestinal tract of the mouse, chitotriosidase is produced at sites of near neutral pH such as the mouth, the non-glandular portion of the stomach, and the small intestine. The expression from the oral cavity to the fore-stomach seems to be restricted to the stratified squamous epithelial cell layer. Chitotriosidase and AMCase activity can be found in the lumen of the gastrointestinal tract, suggesting that these cells secrete the enzyme to the luminal site. However, besides AMCase, considerable chitotriosidase activity can also be found in the circulation. At this time we do not know the exact cellular source of chitotriosidase in the blood. We therefore cannot exclude that a portion of the chitotriosidase produced by the stratified epithelial layer is secreted to the blood. Alternatively, it is possible that another, so far unidentified, cell type is responsible for the chitotriosidase in the circulation.
The acid-stable AMCase is produced predominantly in the glandular portion of the stomach, at the bottom of the gastric glands in chief cells, close to the acid-producing parietal cells, consistent with its features. Stomach lysozymes from ruminants have adapted to function optimally in the hostile acid environment by subtle changes in amino acid composition. The same is also the case for the mouse AMCase that has also adapted to function in the stomach. Analysis of the three-dimensional structure of chitotriosidase and AMCase should reveal the key amino acid substitutions required for the adaptation. The available information on the structure of chitotriosidase should assist further investigations on this matter (Fusetti et al. 2002).
The human AMCase is less well equipped to function in a highly acidic environment compared with the mouse enzyme. Differences in physiology or diet may have contributed to this. Live insects are an important component in the diet of wild mice (Landry 1970).
Intriguingly, in the cow AMCase is produced and secreted by the liver and is present in large amounts in serum (Suzuki et al. 2001). The function of the enzyme is unclear. It seems likely that expression is driven by yet another type of promoter, adding to the rapidly growing complexity of the mammalian chitinase protein family.
Overexpression of chitinases occurs in a number of pathologies in mouse and man. Chitotriosidase is the dominant chitinase in man that is highly expressed in specific cell types including tissue macrophages. In various disorders in which activated macrophages are implicated, elevated plasma chitotriosidase levels occur, e.g., lysosomal lipid storage disorders, sarcoidosis, visceral Leishmaniasis, extended atherosclerosis such as Tangier disease, and thalassemia (Hollak et al. 1994; Barone et al. 1999; Boot et al. 1999; Grosso et al. 2004). In sharp contrast, in mice, expression of chitotriosidase is confined to the gastrointestinal tract and AMCase seems to be the sole endogenous chitinase in tissues such as lung. These species differences seem to be due to distinct promoters of the chitotriosidase genes in mouse and man. The mouse chitotriosidase transcription start site is located far upstream compared with the human situation because the mouse gene contains an extra exon at the 5’ end. Homology is noted between the human promoter and a region in intron 1 of the mouse gene, just upstream of the second exon. Whether this region can indeed act as an alternative promoter for the mouse chitotriosidase gene is at present not known and a topic of further investigation.
The promoter regions of the mouse and human AMCase genes are relatively comparable.
In contrast to Suzuki and coworkers (2002), we were able to detect AMCase mRNA in murine lung with ISH, confirming our Northern blot analysis (Boot et al. 2001). This discrepancy might be due to differences in mice strains or to the sensitivity of the ISH methods. The expression of AMCase mRNA is found to be induced in the lung of mice under various pathological conditions, most strikingly in Th2-driven asthma models (Sandler et al. 2003; Xu et al. 2003; Zhu et al. 2004; Zimmermann et al. 2004). Prominently increased expression of AMCase has also been observed in lung biopsies from asthmatic patients (Zhu et al. 2004).
A surprising role has recently been ascribed to chitinases in the pathogenesis of asthma. It was found that inhibition of chitinase activity in lungs of asthmatic mice alleviates the inflammatory pathology (Zhu et al. 2004). This opens a potential new avenue for therapeutic intervention, i.e., the use of specific chitinase inhibitors such as allosamidin (Zhu et al. 2004). However, extrapolation of the findings with mice to man is complicated by the fact that it is presently unclear whether in addition to AMCase, chitotriosidase is also overexpressed in lung of asthmatic patients. It has been shown that in sarcoidosis, a systemic granulomatous disease, chitotriosidase is elevated in plasma and bronchoalveolar lavage (Hollak et al. 1994; Grosso et al. 2004). It is therefore unclear whether inhibition of both AMCase and chitotriosidase would be required for intervention in the inflammatory process in lungs of asthmatic patients. It may also be possible that selective inhibition of AMCase is more desirable. In this regard, it will be of interest to study whether the relatively common chitotriosidase deficiency influences the clinical course of asthma.
In conclusion, the remarkable differences in cell type and tissue-specific expression of chitotriosidase in man and mouse seem to be mediated by usage of different promoter regions, gene duplications, and different environmental pressures during the evolution of these different species. Further research is required regarding the physiological functions of the chitinase and their potentially harmful role in excessive inflammatory responses.
Footnotes
Acknowledgements
We would like to acknowledge Wilma Donker-Koopman, Roelof Ottenhoff, and Anneke Strijland for their skillful technical assistance. Ans Groener and Marco van Eijk are acknowledged for their helpful comments and suggestions during the preparation of the manuscript.