
Abstract
Recently, the second mammalian chitinase, designated acidic mammalian chitinase (AMCase), has been identified in human, mouse, and cow. In contrast to the earlier identified macrophage-derived chitinase (chitotriosidase), this chitinase is richly expressed in the gastrointestinal (GI) tract, suggesting its role in digestion of chitin-containing foods as well as defense against chitin-coated microorganisms and parasites. This in situ hybridization study first revealed cellular localization of the gut-type chitinase in the mouse and chicken. In adult mice, the parotid gland, von Ebner's gland, and gastric chief cells, all of which are exocrine cells of the serous type, expressed the gut chitinase mRNA. In the chicken, oxyntico-peptic cells in glandular stomach (proventriculus) and hepatocytes expressed the chitinase mRNA. Because cattle produce the gut chitinase (chitin-binding protein b04) only in the liver, the gut chitinases in mammals and birds have three major sources of production, i.e., the salivary gland, stomach, and liver. During ontogenetic development, the expression level in the parotid gland and stomach of mice increased to the adult level before weaning, whereas in the stomach of chickens intense signals were detectable in embryos from incubation day 7.
Keywords
C
Independently of these studies, our research group (Suzuki et al. 2001) identified a chitinase from bovine serum on the basis of its chitin-binding ability and named it chitin-binding protein b04 (CBPb04). High sequence homology of CBPb04 to human AMCase (83.7% in amino acid sequence) showed that CBPb04 was a bovine counterpart of human AMCase (Suzuki et al. 2001). Surprisingly, the “bovine AMCase” is produced only by the liver, differing in tissue distribution from human and rodent AMCase. The bovine AMCase is synthesized in hepatocytes but not in hepatic macrophages (Kupffer cells), and is secreted into the blood. The serum chitinolytic activity of the bovine AMCase was sufficient for some innate immuno-responses, such as protection against chitin-containing microorganisms, suggesting its defensive role against exogenous pathogens in circulation (Suzuki et al. 2001). In contrast to the hepatic AMCase, salivary and gastric AMCase must be secreted into the lumen of the GI tract. To reveal the functional and evolutional aspects of the AMCase family, information about its cellular distribution is needed. In the present in situ hybridization study, we thoroughly examined the cellular expression of AMCase mRNA in the GI tract and other visceral organs of adult and developing mice and chickens. For nomenclature, we adopted the term “gut chitinase” for AMCase and CBPb04 in the present study, since their counterparts were found in the stomach and liver of the chicken (this study) and in the stomach of the frog (unpublished data).
Materials and Methods
Probes for In Situ Hybridization
Three non-overlapping antisense oligonucleotides were used for in situ hybridization (ISH). They were complementary to nucleotide residues 81–125 (tatttcaccaactgggcccagtatcggccaggtctggggagcttc), 541–585 (ttactgctgctgtagctggtgggatttccaacatccaggctggct), and 1190–1234 (tcctgacgtgccttccgagccagtgactactcctccaggaagtgg) of mouse gut chitinase (AMCase) cDNA (Boot et al. 2001; database accession no. AF290003). Two non-overlapping antisense oligonucleotides each for chicken gut chitinase and mouse pepsinogen C were also designed. Probes for chicken gut chitinase were complementary to nucleotide sequences 328–372 (ttctccacaatggtttccactcctgagaaccgccagaccttcatc) and 556–600 (gctgccgttgctgcaggactttccaacattcaggctggctaccag) of chicken gut chitinase (CBPch04) cDNA (AB071038), which was cloned and sequenced in the present study. The stomach in adult rodents expresses only pepsinogen C among the pepsinogen family (Kageyama et al. 2000). Because the nucleotide sequence of mouse pepsinogen C has not been reported, we searched, via a database, for a mouse EST clone showing high homology with rat pepsinogen C cDNA (Ichihara et al. 1986). Thus, two antisense probes for mouse pepsinogen C were designed against 121–165 (gtgttcggtggcgtggacgagaacctgtacactggcgagctcacc) and 174–218 (tgtcacccaggagctttactggcagatcaccattgacgacttcct) of Mus musculus adult stomach cDNA to mRNA (AV080004). These oligonucleotides were all labeled with [35S]-dATP, using terminal deoxyribonucleotidyl transferase (Promega; Madison, WI) at a specific activity of 0.5 × 109 dpm/μg DNA.
Tissue Samples
Male adult ddY mice, 8 weeks old, were used for the present ISH. The animals were sacrificed by dislocation of cervical vertebrae, and the salivary glands (parotid gland, submandibular gland, and sublingual gland), lips, tongue, palate, stomach, various regions of the intestine (from duodenum to rectum), liver, pancreas, lung, kidney, adrenal gland, and urogenital tract were rapidly removed and frozen in liquid nitrogen. The salivary glands and stomach were also collected from neonatal ddY mice of both sexes at 0, 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, or 24 days of age. Cryostat sections about 14 μm thick, were prepared and mounted on glass slides precoated with 3-aminopropyltriethoxysilane (Shinetsu Chemical Industry; Tokyo, Japan).
For chicken samples, the oral mucosa, tongue, crop, proventriculus, gizzard, different regions of the intestine (from duodenum to rectum), liver, lung, pancreas, kidney, and spleen were collected from adult White Leghorn hens. The stomach in developing stages was obtained from embryos, from incubation days 6–21, and from neonatal chickens from post-hatching days 0–12.
In Situ Hybridization
The sections were fixed with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.3) for 15 min and acetylated for 10 min with 0.25% acetic anhydride in 0.1 M triethanolamine-HCl (pH 8.0). The sections were prehybridized for 2 hr in a buffer containing 50% formamide, 0.1 M Tris-HCl (pH 8.0), 4 × SSC (1 × SSC = 150 mM NaCl and 15 mM sodium citrate), 0.02% Ficoll, 0.02% polyvinylpyrrolidone, 0.02% bovine serum albumin, 0.6 M NaCl, 0.25% sodium dodecyl sulfate (SDS), 200 μg/ml tRNA, 1 mM EDTA, and 10% dextran sulfate. Hybridization was performed at 42C for 10 hr in the prehybridization buffer supplemented with 10,000 cpm/μl of 35S-labeled oligonucleotide probes. The slides were washed at room temperature (RT) for 30 min in 2 × SSC containing 0.1% sarkosyl and twice at 55C for 40 min each in 0.1 × SSC containing 0.1% sarkosyl. The sections were dipped in Kodak NTB2 nuclear track emulsion (Kodak; Rochester, NY) and exposed for 1 month.
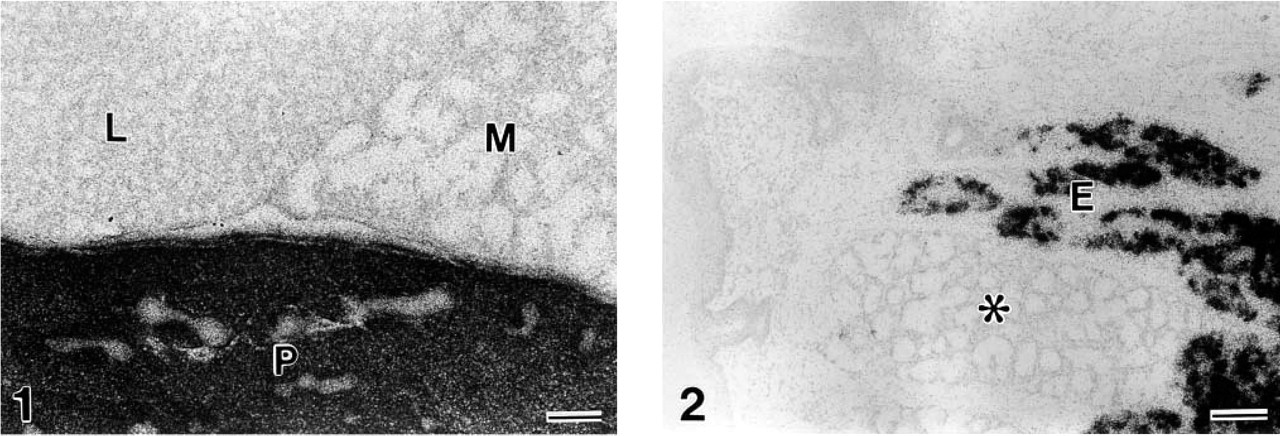
Gut chitinase mRNA expression in the salivary gland of the mouse. Intense signals are localized in the parotid gland (P) but not in the submandibular gland (M) or sublingual gland (L). In the tongue (Figure 2), the mRNA is expressed only in the secretory portion of Ebner's gland (E). It is not detectable in the lingual gland of the mucous type (asterisk) in juxtaposition to Ebner's gland. Bars = 100 μm.
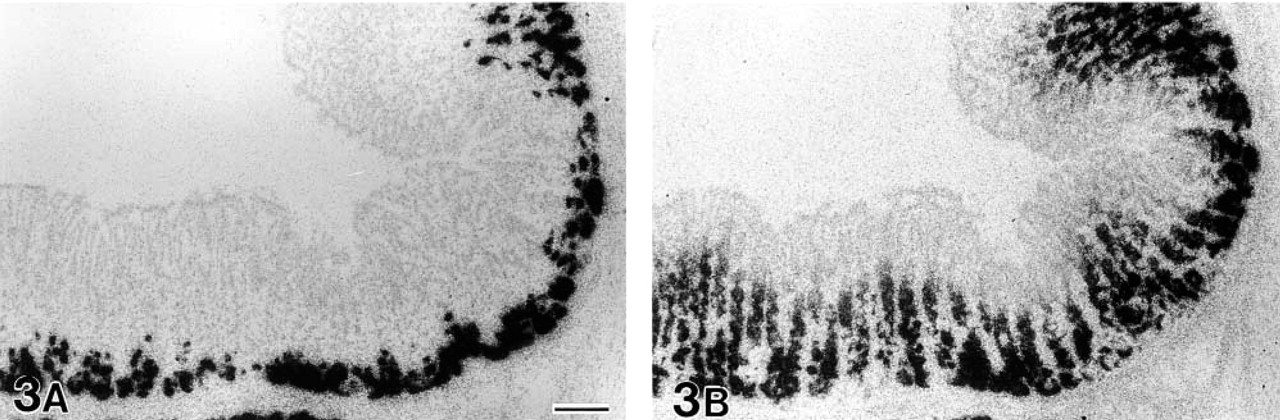
Comparison of gut chitinase and pepsinogen C mRNA expression using serial sections from the glandular stomach of an adult mouse. Chitinase mRNA expression is restricted to the bottom region of gastric glands (
Gut chitinase and pepsinogen C mRNA expression in the proximal part of the pyrolic antrum in an adult mouse. No signals for chitinase mRNA are found in the pyloric antrum (
Cloning of Chicken Gut Chitinase
Total proventriculus RNA was isolated from a Leghorn chicken using TRIzol reagent (Gibco BRL; Grand Island, NY). First-strand cDNA synthesis was performed on 10 μg of total RNA using SuperScript II RNase H− reverse transcriptase (Gibco BRL) and a random primer. The first-strand cDNA was used as the template for polymerase chain reaction (PCR) amplification with Taq DNA polymerase (Promega) and a degenerated primer set. The primers were designed using human (AF290004) and bovine (AB051629) sequences. DNA sequence analysis was performed using the Big Dye terminator method (PE Applied Biosystems; Foster City, CA).
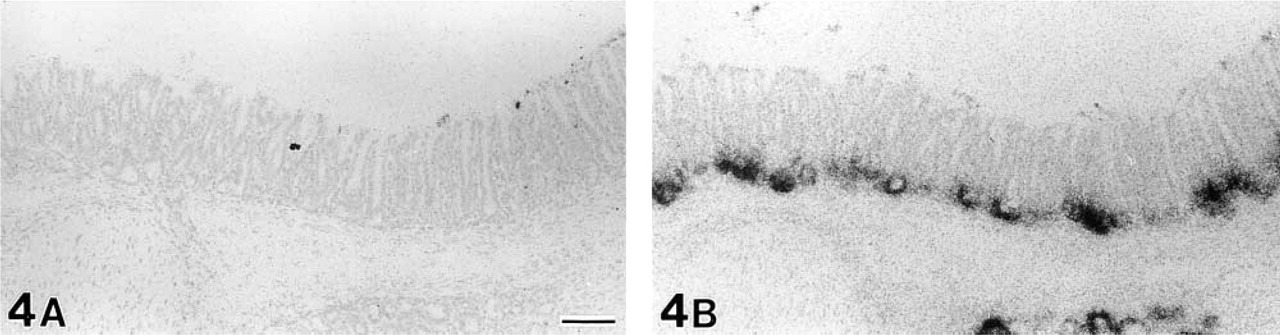
Developmental changes of gut chitinase mRNA expression in the parotid gland at 12, 14, and 16 days after birth. At PD 12 (
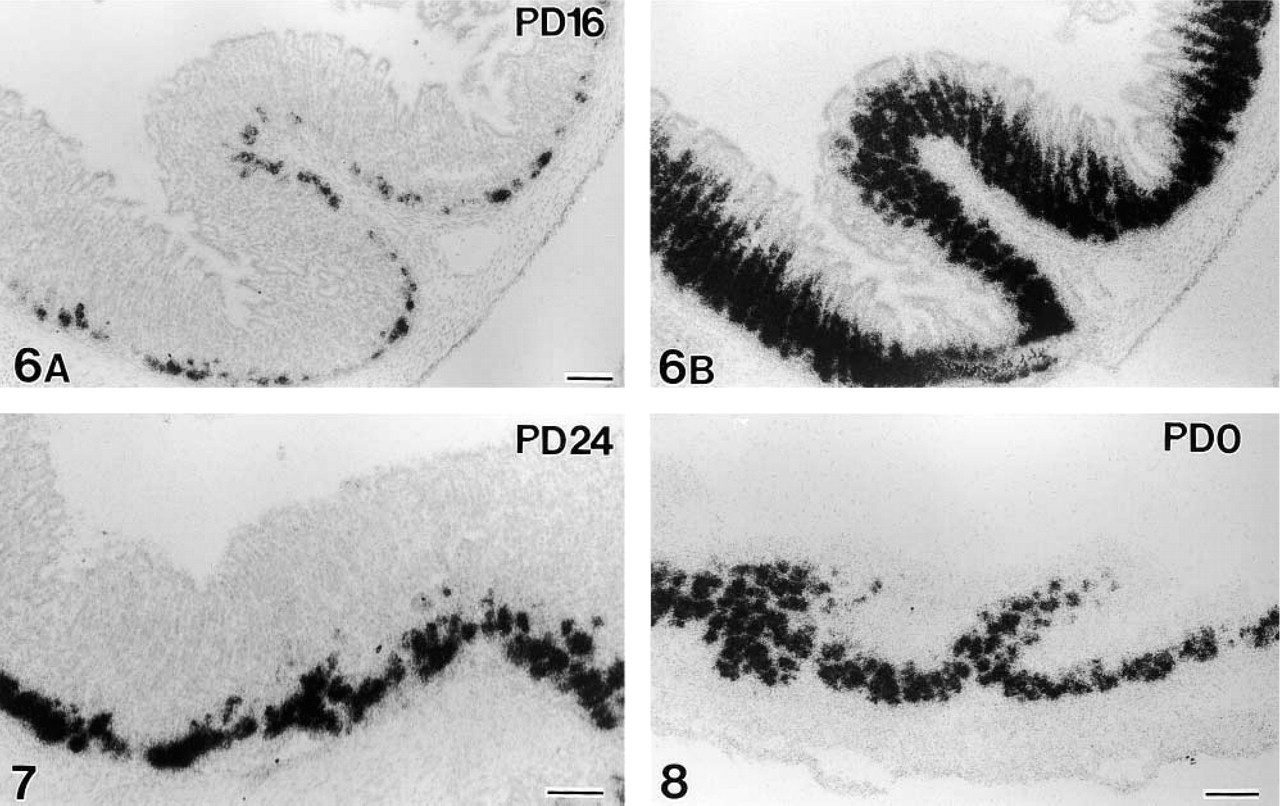
Gut chitinase and pepsinogen C mRNA expression in the glandular stomach during development. The first signals for chitinase mRNA during ontogeny appear at PD 16, dispersed along the bottom of gastric glands (Figure 6A). Pepsinogen mRNA expression at PD 16 (Figure 6B, adjacent section to Figure 6A) shows the same intensity as seen in adults. Intense signals of chitinase mRNA comparable with the signals in adults appear at PD 24 (Figure 7). In contrast, fully intense expression for pepsinogen C mRNA is seen even at birth (PD 0; Figure 8). Bars = 100 μm.
Northern Blotting and RT-PCR
Total RNA was separated in 1% agarose/formaldehyde gels, and transferred to nylon membranes (Amersham Pharmacia Biotech; Piscataway, NJ), which were hybridized with an [α32P]-dCTP-labeled chicken gut chitinase probe (nucleotide residues 332–1310) and exposed to X-ray film (Kodak).
To confirm the distribution of chicken gut chitinase cDNA, an RT-PCR experiment was performed using Taq DNA polymerase and first-strand cDNA as the template. The reaction was done using a specific primer set for chicken gut chitinase.
Results
Expression of Gut Chitinase mRNA in Adult Mouse
Intense expression of gut chitinase mRNA was found only in the salivary gland and glandular stomach throughout the GI tract in adult mice (Figures 1–4). Other visceral organs, including the liver, pancreas, lung, kidney, adrenal gland, and urogenital organs, did not exhibit any significant expression. Of the three major salivary glands, only the parotid gland displayed intense expression, with selective localization in the glandular portion (acini), whereas the submandibular and sublingual glands completely lacked the signals (Figure 1). When tissue sections containing small salivary glands, such as the lingual gland, labial gland, and palatal gland, were hybridized, only the lingual gland of the serous type associated with circumvallate papillae, termed von Ebner's gland, expressed similarly intense transcription of gut chitinase mRNA (Figure 2). The specificity of hybridization was confirmed by consistent labeling using three non-overlapping antisense probes specific for mouse gut chitinase mRNA, and also by the disappearance of the signals when an excess dose of cold probes was added to the hybridization fluid (data not shown).

Sequence comparison of chicken gut chitinase with bovine, human and mouse gut chitinases. Amino acids identical to chicken gut chitinase (CBPch04) are blocked in black. ∗ and # indicate the essential acidic amino acids for the catalytic domain and cysteines for the chitin-binding domain, respectively. Chicken, CBPch04 (accession no. AB071038); bovine, CBPb04 (AB051629); human, human AMCase (AF290004); mouse, mouse AMCase (AF290003).
The expression in the glandular stomach was analyzed in detail with special reference to pepsinogen C mRNA expression. Observation of serial sections alternatively hybridized for gut chitinase and pepsinogen C mRNAs revealed limited expression of chitinase mRNA in some pepsinogen C mRNA-expressing cells, thus identifiable as chief cells (Figures 3A and 3B). Pepsinogen C mRNA-expressing cells occupied the basal half of gastric glands, from the bottom to the neck portion of the glands, but distribution of chitinase mRNA-expressing cells was restricted to the basal one-fourth of the gastric glands. Horizontally, pepsinogen cells were distributed throughout the oxyntic mucosa and extended to the proximal part of the pyloric antrum (Figure 4B). However, signals for chitinase mRNA in the oxyntic mucosa gradually decreased in intensity and extent approaching the pyloric mucosa; no significant signals were found in the pyloric antrum (Figure 4A).
Developmental Changes of Gut Chitinase mRNA Expression in Mouse Parotid Gland and Stomach
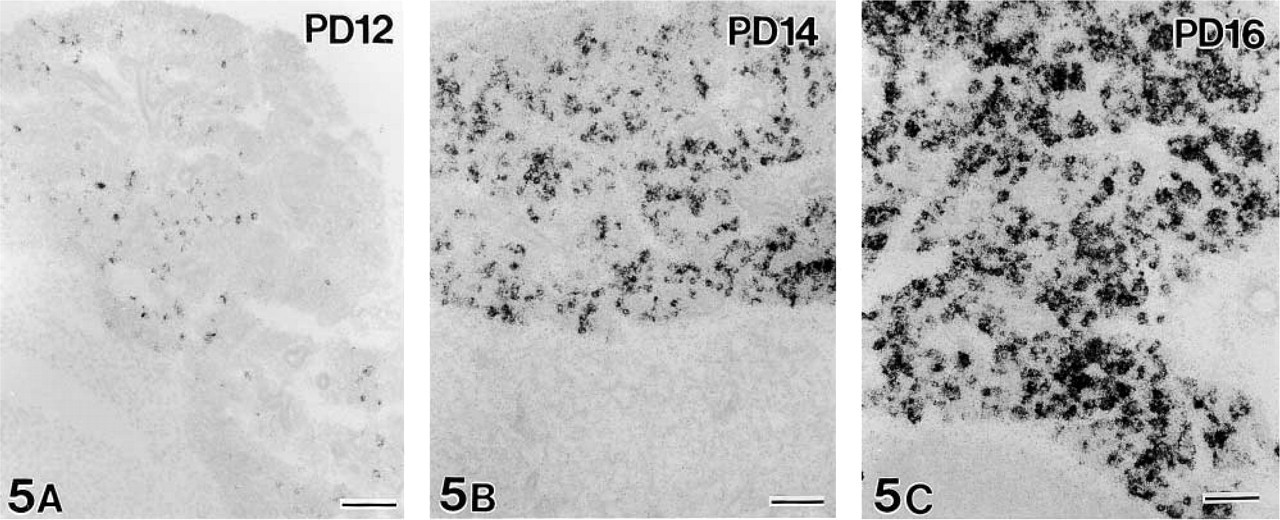
The first signals for gut chitinase mRNA in the developing parotid gland were detected at postnatal day (PD) 12, although they were weak and dispersed (Figure 5A). The intensity of chitinase mRNA expression sharply increased from PD 12 to PD 16 (Figures 5A–5C). The first significant expression of chitinase mRNA in the developing stomach was found at PD 16. Only a small number of cells dispersed at the bottom of gastric glands were labeled (Figure 6A), in comparison to intense and broad expression of pepsinogen C mRNA (Figure 6B). Thereafter, the signals of chitinase mRNA gradually increased in intensity and extent and reached the adult level during PD 20–24 (Figure 7). In contrast, intense signals for pepsinogen C mRNA already appeared at PD 0 (Figure 8). Throughout the developmental stages, no significant expression of gut chitinase mRNA was found in other parts of the GI tract or other visceral organs.
Chicken Gut Chitinase mRNA Expression in the Adult and During Ontogeny
Cloning and sequencing of chicken gut chitinase cDNA was carried out using RNA obtained from the adult chicken proventriculus (glandular stomach). We obtained the cDNA sequence of chicken gut chitinase from a combination of PCR products as described in Materials and Methods. The nucleotide sequence of the cloned cDNA contained an open reading frame 1449 bp long, and the open reading frame encoded a protein 482 amino acids long (CBPch04; accession no. AB071038). Overall sequence comparison of chicken gut chitinase with bovine, human, and mouse chitinases showed 70.2%, 75.8%, and 72.7% identity for the deduced amino acids, respectively (Figure 9). Chicken gut chitinase mRNA was detected in the proventriculus and liver by RT-PCR analysis (Figure 10). On the other hand, by Northern blotting analysis it was detected only in the proventriculus (Figure 11). These results indicated that chicken gut chitinase was expressed more abundantly in the proventriculus than in the liver.
ISH analysis for chicken gut chitinase mRNA expression using two non-overlapping antisense oligonucleotide probes revealed its selective expression in the proventriculus and liver of the adult chicken (Figures 12A and 12B). No signals were detectable in the lung, pancreas, kidney, spleen, and GI tract other than the proventriculus. The proventriculus had extremely intense signals. Oxyntico-peptic cells constituting the deep (proper) gastric glands expressed the mRNA, whereas duct portions of the deep gastric gland and superficial gastric gland lacked the signals (Figure 12A). Less intense but significant signals were found in the liver, where hepatocytes were labeled (Figure 12B).
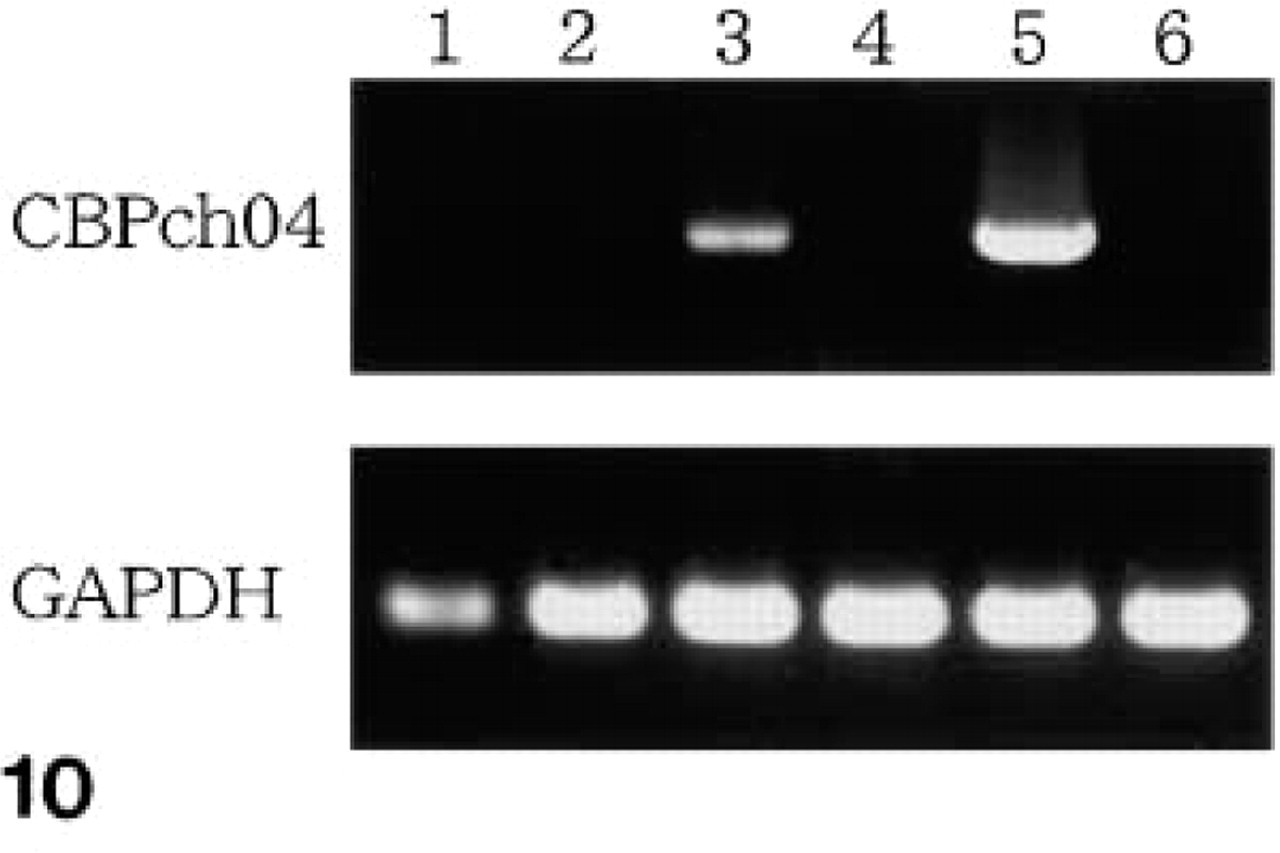
RT-PCR analysis. A specific primer set for CBPch04 was used to detect chicken gut chitinase cDNA (above). As a control, a specific primer set for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used (below). Lane 1, lung; Lane 2, spleen; Lane 3, liver; Lane 4, kidney; Lane 5, proventriculus; Lane 6, gizzard.

Northern blotting analysis of chicken gut chitinase (CBPch04) mRNA. Ten μg of total RNA was separated on an agarose gel. The position 28s and 18s rRNA bands are indicated by arrows. RNA loading was confirmed by ethidium bromide staining (data not shown). Lane 1, heart; Lane 2, lung; Lane 3, liver; Lane 4, proventriculus; Lane 5, gizzard; Lane 6, kidney; Lane 7, spleen; Lane 8, ovary; Lane 9, bursa fabricii.
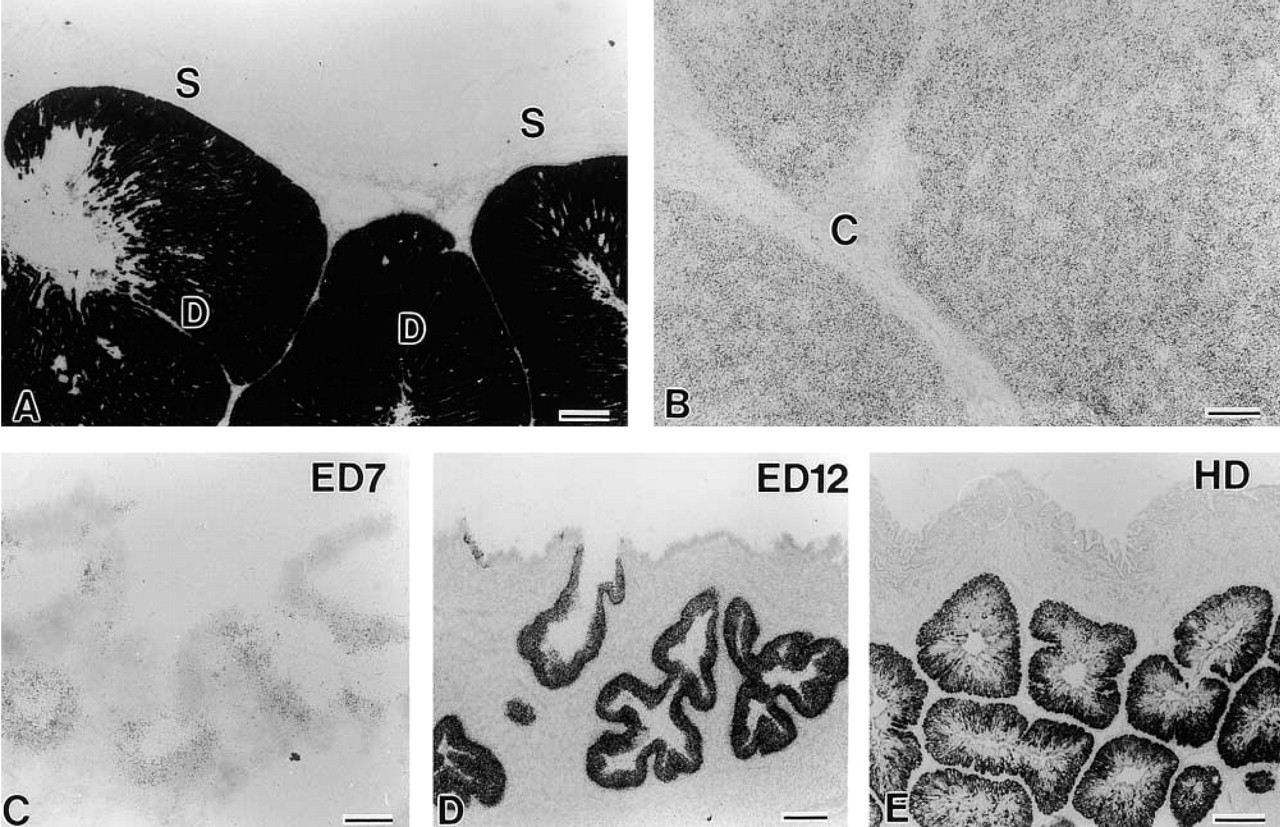
Gut chitinase mRNA expression in the proventriculus of adult chickens and embryos. In the adult the signals are conspicuously intense in the deep gastric gland (D in
In the developing stages, the first significant signals of chicken chitinase mRNA in the proventriculus were found at embryonic day (ED) 7 (Figure 12C), and the signals gradually increased in intensity with development until the hatching day (Figures 12D and 12E). Another sharp increase in intensity appeared around post-hatching day 2, showing the same intensity as seen in the adult chicken.
Discussion
Mammalian gut chitinases have been identified in human, mouse, and cow (Boot et al. 2001; Suzuki et al. 2001). There are high homologies among them, i.e., 67–82% in amino acid sequence. The amino acid residue of chicken gut chitinase is also homologous to those of human (75.8%), mouse (72.7%), and cow (70.2%), as shown by the present study. Similar ho-mology of gut chitinase was found between mammals and amphibians (Rana catesbeiana, Xenopus laevis) (unpublished data). High conservation of the gut chitinase molecules through evolution suggests their important and consistent functions.
Northern blotting analysis by Boot et al. (2001) demonstrated that gut chitinase mRNA was expressed in the submandibular gland and stomach of mouse and in the stomach of human, but not in human salivary gland. The present morphological study first localized gut chitinase expression in the parotid gland and glandular stomach of mouse. There is a discrepancy about the site of expression in the salivary gland between our results and those of Boot et al. (2001). The discrepancy concerning the expression site in the mouse salivary gland may be caused by an uncertain sampling procedure or by difficulty in the exact separation of each gland in their study. Moreover, the selective expression in the parotid gland of mice may explain the failure by Boot et al. (2001) to detect chitinase mRNA in the human salivary gland with Northern blotting analysis. It is possible to consider that their “salivary gland” samples did not contain the parotid gland. The intense expression of chitinase mRNA in the parotid gland and in von Ebner's gland leads us to the conclusion that glands of the purely serous type secrete chitinase. This idea is true in the rodent glandular stomach. The gastric chief cells, which are serous-type secretory cells and gather at the bottom of gastric glands, were selectively labeled in the present ISH.
The present comparative staining for gut chitinase and pepsinogen C mRNAs showed that some chief cells expressed the chitinase mRNA. The distribution of pepsinogen C mRNA-expressing cells in the gastric glands of mice was in accordance with that reported in the rat (Ge et al. 1998). Chief cells, mucous neck cells, and intermediate mucopeptic cells expressed pepsinogen C mRNA. The chief cells originate from mucous neck cells gathering at the neck portion of gastric glands and differentiate into pepsinogen-secreting cells moving towards the bottom of the glands (Suzuki et al. 1983; Ge et al. 1998). Therefore, it is likely that fully differentiated chief cells express only chitinase. The gut chitinase secreted from the parotid gland and gastric chief cells may digest the ingested chitin into fragments in the stomach. The rodent and human gut chitinases are stable in acidic conditions and have a pronounced pH optimum at pH 2.3. The high level of chitinase activity detected in extracts of the intestine has been considered to be due to chitinase secreted from the stomach (Boot et al. 2001). However, because the enzyme activity prominently decreased at over pH 6, the functional significance of the chitinase in the intestinal lumen is doubtful. Interestingly, a pancreatic chitinase with an optimal pH of 6.0 has been identified from the pancreas of the toad Bufo japonicus, suggesting its functional role in the intestinal lumen. This chitinase showed considerable, but not high, amino acid sequence homology (about 50%) with known mammalian gut chitinases (Oshima et al. in press). Further examination is needed to clarify whether intestinal chitinase activity in mammals is derived from gastric and salivary chitinases or from the pancreatic chitinase.
Among the chitinase protein family, Ym1 and Ym2 are known to be expressed in the mammalian stomach (Jin et al. 1998; Ward et al. 2001). The cDNAs of both molecules in the mouse show a homology of about 75% at the open reading frame with that of the mouse gut chitinase. However, it is unlikely that the three probes used in the present study cross-hybridized to Ym1 and Ym2, owing to the low homology of sequences selected for design of the probes with any regions of Ym1 and Ym2 cDNAs. Furthermore, Ym1 and Ym2 are more widely distributed in organs and cell types than the gut chitinase, such as the kidney and thymus, which contained no detectable levels of gut chitinase mRNA in the present ISH study. A comparison of expression patterns between Ym1/Ym2 and gut chitinase under normal and pathological conditions is an interesting topic for future studies.
The liver is another important source of gut chitinase in mammalian species. However, there is a clear species difference in the hepatic expression. Humans and rodents express chitinase mRNA in the salivary gland and stomach but not in the liver, whereas in the cattle only the liver expresses chitinase mRNA (Suzuki et al. 2001). The chicken may be a key animal model in which to explain the species difference in chitinase expression. Avian species have dual sources for chitinase production in the stomach and liver, and mammals may have selected the major source, either the alimentary canal or liver, through evolution. The chitinase produced in hepatocytes circulates in the bloodstream and specifically defends against chitin-containing microorganisms via its chitin-binding and chitin-fragmenting abilities (Suzuki et al. 2001). In human, which lacks hepatic chitinase, serum contains the macrophage-derived chitinase, chitotriosidase. On the other hand, because cattle have abundant chitinolytic activities derived from lysosomes or chitinolytic microorganisms in the stomach (abomasum and rumen) (Pahud et al. 1983; Kopecny et al. 1996), the bovine GI tract may not require chitinase of host origin. Northern blotting analysis for the gut chitinase demonstrated mRNA expression, although far less, in the lungs, and the enzyme activity was found in the lung extracts (Boot et al. 2001). Moreover, Boot et al. (2001) and Saito et al. (1999) used these sources to clone mouse and human gut chitinase cDNAs, respectively. These data are contrary to the present study, which failed to detect a significant expression of gut chitinase mRNA in the lung. This discrepancy may be explained by the fact that mRNA expression was below detectable levels by the ISH.
During the developing stages, it is reasonable that the first significant expression of gut chitinase appeared at PD 14 and 16 in the parotid gland and stomach, respectively, and rapidly increased in intensity in the following several days. This (possibly digestive) enzyme may be prepared just before the weaning period (about 21 days), whereas pepsinogen C is fully expressed at birth in the mouse (this study) and rat (Ge et al. 1998) to digest milk proteins. In the chicken, however, significant expression of gut chitinase was recognized in embryos even at day 7 of incubation, although the mRNA expression further increased after hatching. Although the functional significance of the expression in the early stages of development remains to be elucidated, this finding may be comparable to previous observations showing that chitinase and chitinase-related proteins affect embryogenesis in zebrafish (Semino and Allende 2000) and were identified as growth factors in Drosophila (Kawamura et al. 1999). Further studies are necessary to reveal the involvement of gut chitinase in organogenesis or differentiation of the chick embryo stomach.
Footnotes
Acknowledgements
Supported by a grant from the “Research for the Future” Program of the Japan Society for the Promotion of Science (JSPS-RFTF 97L00906).